
Antiglycation Effects of Adlay Seed and Its Active Polyphenol Compounds: An In Vitro Study

 ,
,  ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Effects of the Individual Parts of Adlay on Protein Glycation According to BSA-Glucose Assay
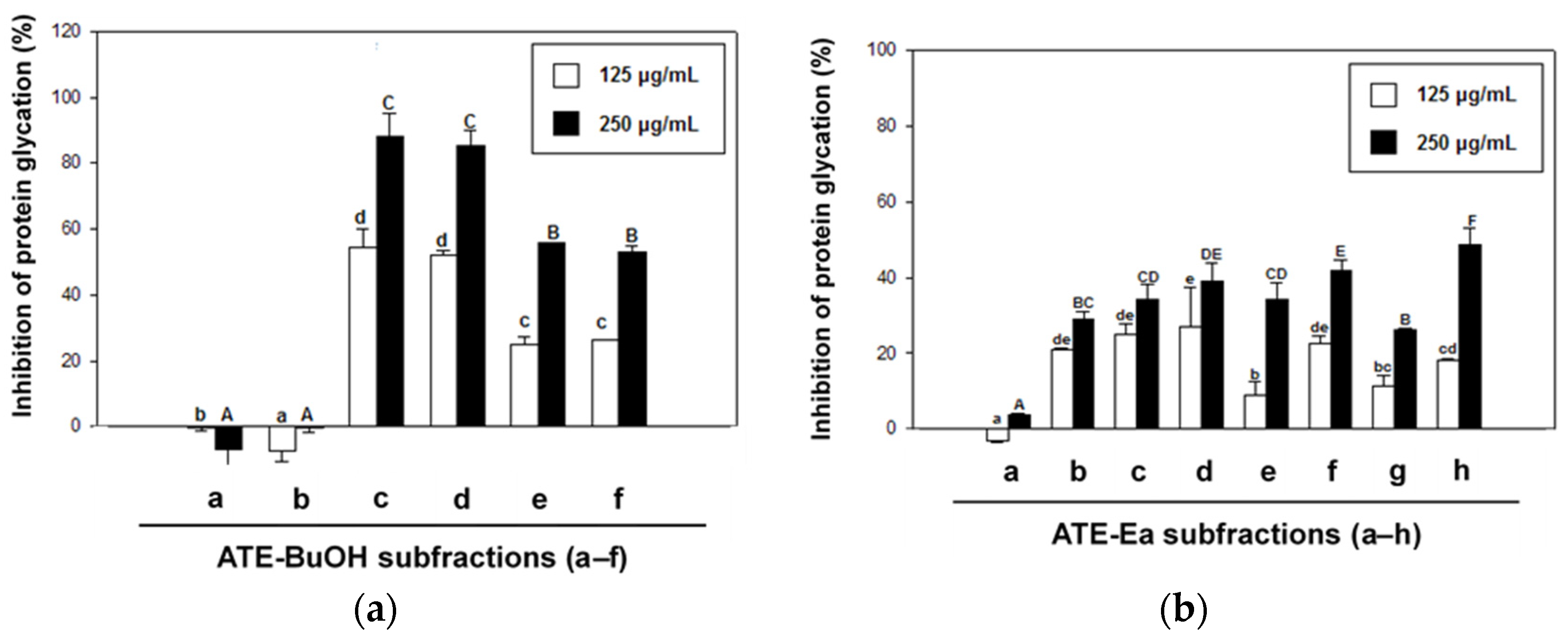
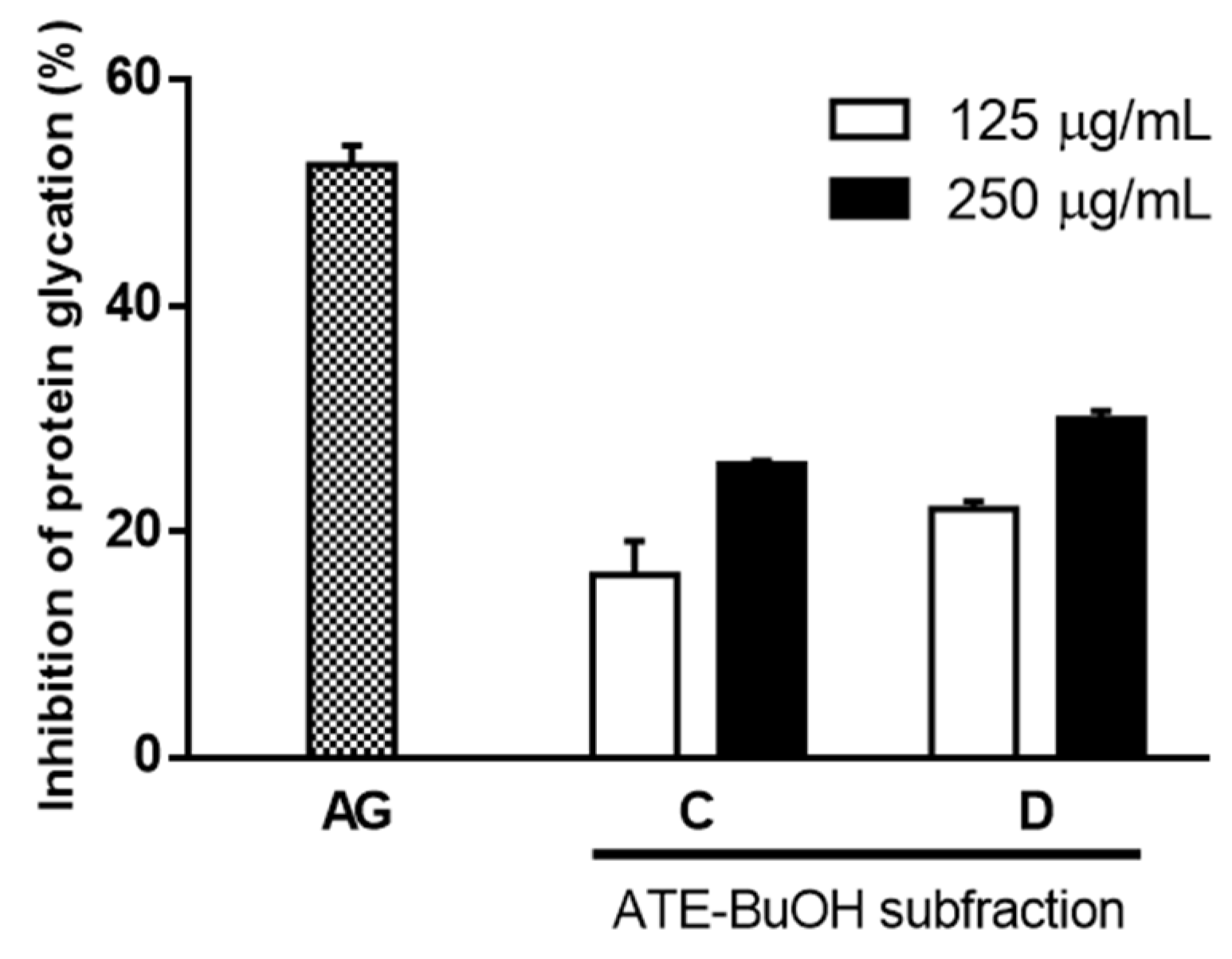
2.2. Effects of ATE and ABE Subfractions on Protein Glycation
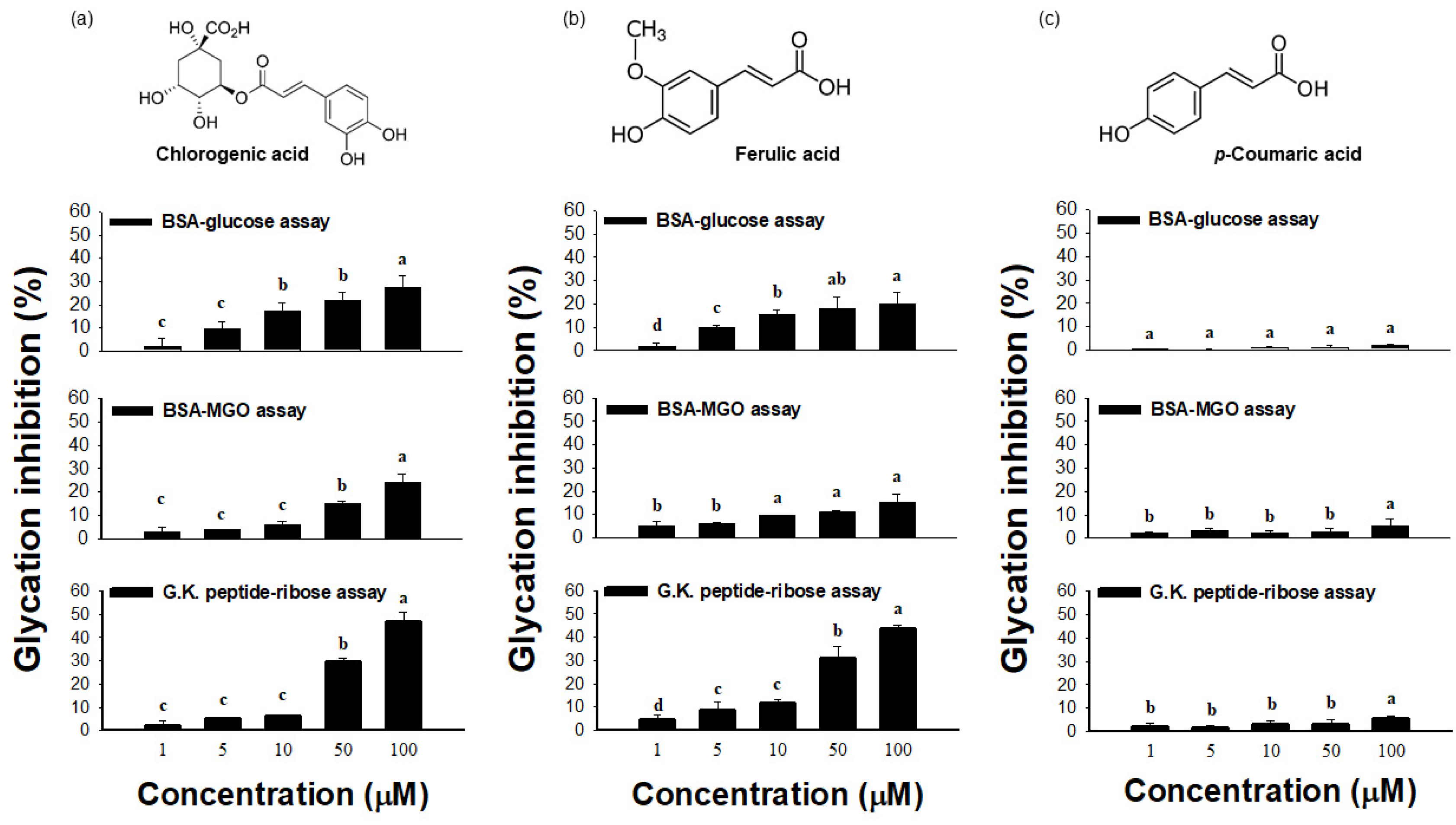
2.3. Effect of ATE-BuOH-Containing Phenolics on the Individual Stage of Protein Glycation
3. Discussion
4. Materials and Methods
4.1. Preparation of Adlay Extracts
4.2. BSA-Glucose Assay
4.3. BSA-MGO Assay
4.4. G.K. Peptide−Ribose Assay
4.5. HPLC Analysis
4.6. Statistics
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Yeh, W.J.; Hsia, S.M.; Lee, W.H.; Wu, C.H. Polyphenols with antiglycation activity and mechanisms of action: A review of recent findings. J. Food Drug Anal. 2017, 25, 84–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vistoli, G.; De Maddis, D.; Cipak, A.; Zarkovic, N.; Carini, M.; Aldini, G. Advanced glycoxidation and lipoxidation end products (AGEs and ALEs): An overview of their mechanisms of formation. Free Radic. Res. 2013, 47, 3–27. [Google Scholar] [CrossRef] [Green Version]
- Takeuchi, M. Toxic AGEs (TAGE) theory: A new concept for preventing the development of diseases related to lifestyle. Diabetol. Metab. Syndr. 2020, 12, 105. [Google Scholar] [CrossRef] [PubMed]
- Gill, V.; Kumar, V.; Singh, K.; Kumar, A.; Kim, J.J. Advanced glycation end products (AGEs) may be a striking link between modern diet and health. Biomolecules 2019, 9, 888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sellegounder, D.; Zafari, P.; Rajabinejad, M.; Taghadosi, M.; Kapahi, P. Advanced glycation end products (AGEs) and its receptor, RAGE, modulate age-dependent COVID-19 morbidity and mortality. A review and hypothesis. Int. Immunopharmacol. 2021, 98, 107806. [Google Scholar] [CrossRef] [PubMed]
- González, I.; Morales, M.A.; Rojas, A. Polyphenols and AGEs/RAGE axis. Trends and challenges. Food Res. Int. 2020, 129, 108843. [Google Scholar] [CrossRef]
- Thornalley, P.J. Use of aminoguanidine (Pimagedine) to prevent the formation of advanced glycation endproducts. Arch. Biochem. Biophys. 2003, 419, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.H.; Yen, G.C. Inhibitory effect of naturally occurring flavonoids on the formation of advanced glycation endproducts. J. Agric. Food Chem. 2005, 53, 3167–3173. [Google Scholar] [CrossRef] [PubMed]
- Jahan, H.; Choudhary, M.I. Glycation, carbonyl stress and AGEs inhibitors: A patent review. Expert Opin. Ther. Pat. 2015, 25, 1267–1284. [Google Scholar] [PubMed]
- Zeng, Y.; Yang, J.; Chen, J.; Pu, X.; Li, X.; Yang, X.; Yang, L.; Ding, Y.; Nong, M.; Zhang, S.; et al. Actional mechanisms of active ingredients in functional food adlay for human health. Molecules 2022, 27, 4808. [Google Scholar] [CrossRef] [PubMed]
- Kuo, C.C.; Chen, H.H.; Chiang, W.J. Adlay (yi yi; “soft-shelled job’s tears”; the seeds of Coix lachryma-jobi L. var. ma-yuen Stapf) is a potential cancer chemopreventive agent toward multistage carcinogenesis processes. Tradit. Complement. Med. 2012, 2, 267–275. [Google Scholar] [CrossRef] [Green Version]
- Yang, R.S.; Lu, Y.H.; Chiang, W.; Liu, S.H. Osteoporosis prevention by adlay (yì yǐ: The seeds of Coix lachryma-jobi L. var. ma-yuen Stapf) in a mouse model. J. Tradit. Complement. Med. 2013, 3, 134–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, D.W.; Wu, C.H.; Shih, C.K.; Liu, C.Y.; Shih, P.H.; Shieh, T.M.; Lin, C.I.; Chiang, W.C.; Hsia, S.M. Application of the solvent extraction technique to investigation of the anti-inflammatory activity of adlay bran. Food Chem. 2014, 145, 445–453. [Google Scholar] [CrossRef] [PubMed]
- Tseng, Y.H.; Chang, C.W.; Chiang, W.; Hsieh, S.C. Adlay Bran oil suppresses hepatic gluconeogenesis and attenuates hyperlipidemia in type 2 diabetes rats. J. Med. Food 2019, 22, 22–28. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Zhang, W.; Shi, R.; Tang, B.; Xie, S. Coix lachryma-jobi extract ameliorates inflammation and oxidative stress in a complete Freund’s adjuvant-induced rheumatoid arthritis model. Pharm. Biol. 2019, 57, 792–798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiang, H.; Lu, H.F.; Chen, J.C.; Chen, Y.H.; Sun, H.T.; Huang, H.C.; Tien, H.H.; Huang, C. Adlay seed (Coix lacryma-jobi L.) extracts exhibit a prophylactic effect on diet-induced metabolic dysfunction and nonalcoholic fatty liver disease in mice. Evid. Based Complement. Altern. Med. 2020, 2020, 9519625. [Google Scholar] [CrossRef] [PubMed]
- Chung, C.P.; Lee, M.Y.; Hsia, S.M.; Chiang, W.; Kuo, Y.H.; Hsu, H.Y.; Lin, Y.L. Suppression on allergic airway inflammation of dehulled adlay (Coix lachryma-jobi L. var. ma-yuen Stapf) in mice and anti-degranulation phytosterols from adlay bran. Food Funct. 2021, 12, 12788–12799. [Google Scholar] [CrossRef]
- Yeh, W.J.; Ko, J.; Cheng, W.Y.; Yang, H.Y. Dehulled adlay consumption modulates blood pressure in spontaneously hypertensive rats and overweight and obese young adults. Nutrients 2021, 13, 2305. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.J.; Shih, C.K.; Hsu, H.Y.; Chiang, W. Mast cell-dependent allergic responses are inhibited by ethanolic extract of adlay (Coix lachryma-jobi L. var. ma-yuen Stapf) testa. J. Agric. Food Chem. 2010, 58, 2596–2601. [Google Scholar] [CrossRef] [PubMed]
- Hsia, S.M.; Kuo, Y.H.; Chiang, W.; Wang, P.S. Effects of adlay hull extracts on uterine contraction and Ca2+ mobilization in the rat. Am. J. Physiol. Endocrinol. Metab. 2008, 295, E719–E726. [Google Scholar] [CrossRef]
- Nagai, E.; Iwai, M.; Koketsu, R.; Okuno, Y.; Suzuki, Y.; Morimoto, R.; Sumitani, H.; Ohshima, A.; Enomoto, T.; Isegawa, Y. Anti-influenza virus activity of adlay tea components. Plant Foods Hum. Nutr. 2019, 74, 538–543. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.W.; Chung, C.P.; Kuo, Y.H.; Lin, Y.L.; Chiang, W. Identification of compounds in adlay (Coix lachryma-jobi L. var. ma-yuen Stapf) seed hull extracts that inhibit lipopolysaccharide-induced inflammation in RAW 264.7 macrophages. J. Agric. Food Chem. 2009, 57, 10651–10657. [Google Scholar] [CrossRef]
- Rahbar, S.; Yerneni, K.K.; Scott, S.; Gonzales, N.; Lalezari, I. Novel inhibitors of advanced glycation endproducts (part II). Mol. Cell Biol. Res. Commun. 2000, 3, 360–366. [Google Scholar] [CrossRef] [PubMed]
- Rowan, S.; Bejarano, E.; Taylor, A. Mechanistic targeting of advanced glycation end-products in age-related diseases. Biochim. Biophys. Acta. Mol. Basis Dis. 2018, 1864, 3631–3643. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.Q.; Zhang, D.D.; Wang, Y.N.; Tan, Y.Q.; Yu, X.Y.; Zhao, Y.Y. AGE/RAGE in diabetic kidney disease and ageing kidney. Free Radic. Biol. Med. 2021, 171, 260–271. [Google Scholar] [CrossRef] [PubMed]
- Huebschmann, A.G.; Regensteiner, J.G.; Vlassara, H.; Reusch, J.E.B. Diabetes and advanced glycation glycoxidation end products. Diabetes Care 2006, 29, 1420–1432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sultana, B.; Anwar, F.; Ashraf, M. Effect of extraction solvent/technique on the antioxidant activity of selected medicinal plant extracts. Molecules 2009, 14, 2167–2180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Y.J.; Chen, Y.C.; Chen, H.Y.; Chiang, Y.F.; Ali, M.; Chiang, W.; Chung, C.P.; Hsia, S.M. Ethanolic extracts of adlay testa and hull and their active biomolecules exert relaxing effect on uterine muscle contraction through blocking extracellular calcium influx in ex vivo and in vivo studies. Biomolecules 2021, 11, 887. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.J.; Chang, C.C.; Wang, Y.Y.; Chiang, W.C.; Shih, Y.H.; Shieh, T.M.; Wang, K.L.; Ali, M.; Hsia, S.M. Adlay testa (Coix lachryma-jobi L. var. Ma-yuen Stapf.) Ethanolic extract and its active components exert anti-proliferative effects on endometrial cancer cells via cell cycle arrest. Molecules 2021, 26, 1966. [Google Scholar] [CrossRef]
- Kim, D.O.; Lee, C.Y. Comprehensive study on vitamin C equivalent antioxidant capacity (VCEAC) of various polyphenolics in scavenging a free radical and its structural relationship.Crit. Rev. Food Sci. Nutr. 2004, 44, 253–273. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.H.; Huang, H.W.; Lin, J.A.; Huang, S.M.; Yen, G.C. The proglycation effect of caffeic acid leads to the elevation of oxidative stress and inflammation in monocytes, macrophages and vascular endothelial cells. J. Nutr. Biochem. 2011, 22, 585–594. [Google Scholar] [CrossRef] [PubMed]
- Woodside, J.V.; Yarnell, J.W.; McMaster, D.; Young, I.S.; Harmon, D.L.; McCrum, E.E.; Patterson, C.C.; Gey, K.F.; Whitehead, A.S.; Evans, A. Effect of B-group vitamins and antioxidant vitamins on hyperhomocysteinemia: a double-blind, randomized, factorial-design, controlled trial. Am. J. Clin. Nutr. 1998, 67, 858–866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Bari, L.; Scirè, A.; Minnelli, C.; Cianfruglia, L.; Kalapos, M.P.; Armeni, T. Interplay among oxidative stress, methylglyoxal pathway and S-glutathionylation. Antioxidants 2020, 10, 19. [Google Scholar] [CrossRef] [PubMed]
- Nagaraj, R.H.; Shipanova, I.N.; Faust, F.M. Protein cross-linking by the Maillard reaction. Isolation, characterization, and in vivo detection of a lysine-lysine cross-link derived from methylglyoxal. J. Biol. Chem. 1996, 271, 19338–19345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuura, N.; Aradate, A.; Sasaki, C.; Kojima, H.; Ohara, M.; Hasegawa, J.; Ubukata, M. Screening system for the Maillard reaction inhibitor from natural product extracts. J. Health Sci. 2002, 48, 520–526. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chung, C.-P.; Hsia, S.-M.; Chang, W.-S.; Huang, D.-W.; Chiang, W.-C.; Ali, M.; Lee, M.-Y.; Wu, C.-H. Antiglycation Effects of Adlay Seed and Its Active Polyphenol Compounds: An In Vitro Study. Molecules 2022, 27, 6729. https://doi.org/10.3390/molecules27196729
Chung C-P, Hsia S-M, Chang W-S, Huang D-W, Chiang W-C, Ali M, Lee M-Y, Wu C-H. Antiglycation Effects of Adlay Seed and Its Active Polyphenol Compounds: An In Vitro Study. Molecules. 2022; 27(19):6729. https://doi.org/10.3390/molecules27196729
Chicago/Turabian StyleChung, Cheng-Pei, Shih-Min Hsia, Wen-Szu Chang, Din-Wen Huang, Wen-Chang Chiang, Mohamed Ali, Ming-Yi Lee, and Chi-Hao Wu. 2022. "Antiglycation Effects of Adlay Seed and Its Active Polyphenol Compounds: An In Vitro Study" Molecules 27, no. 19: 6729. https://doi.org/10.3390/molecules27196729
APA StyleChung, C. -P., Hsia, S. -M., Chang, W. -S., Huang, D. -W., Chiang, W. -C., Ali, M., Lee, M. -Y., & Wu, C. -H. (2022). Antiglycation Effects of Adlay Seed and Its Active Polyphenol Compounds: An In Vitro Study. Molecules, 27(19), 6729. https://doi.org/10.3390/molecules27196729