
Comparison of Phenolic Metabolites in Purified Extracts of Three Wild-Growing Herniaria L. Species and Their Antioxidant and Anti-Inflammatory Activities In Vitro

 ,
,  ,
, 

Abstract
:1. Introduction
2. Results and Discussion
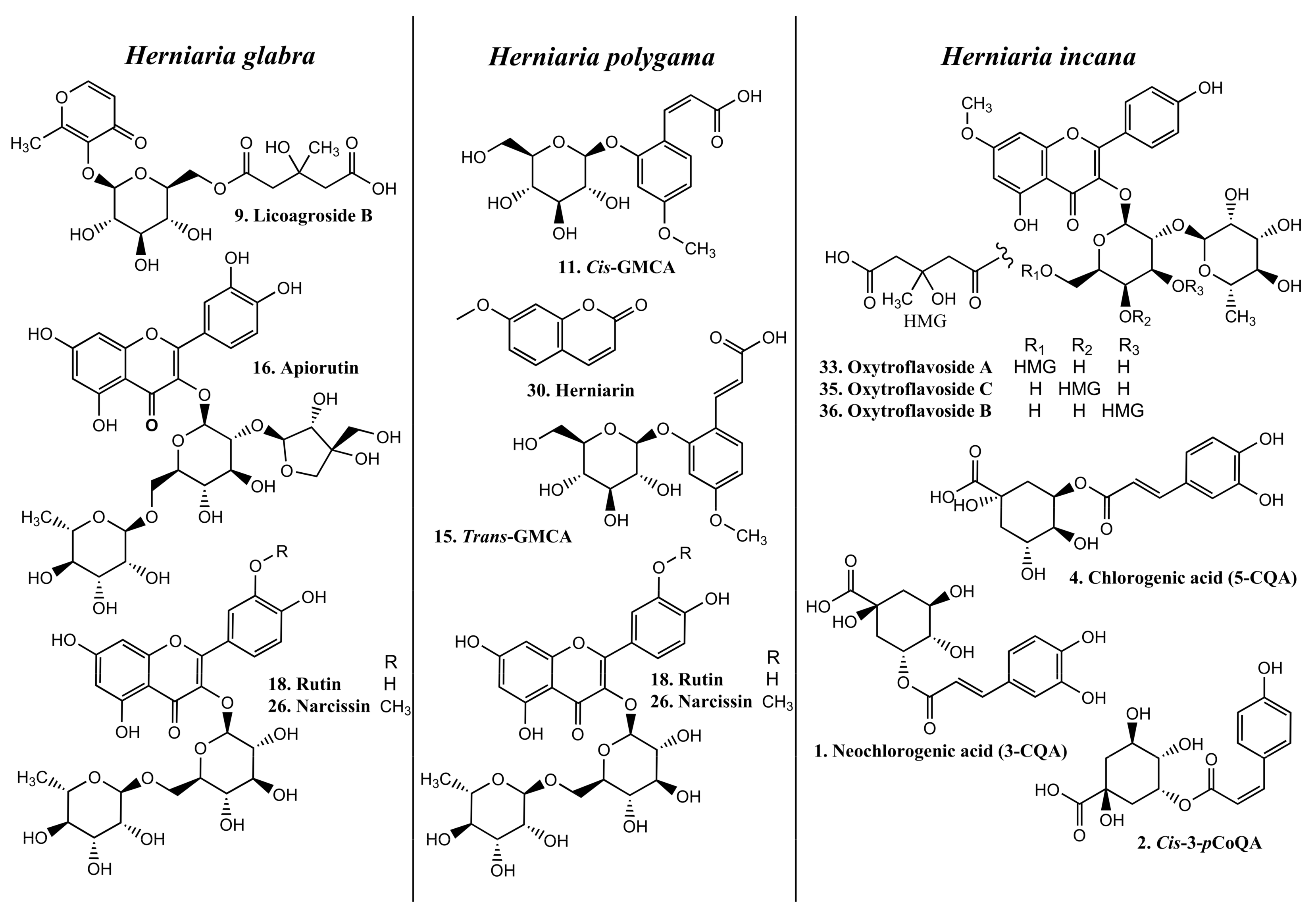
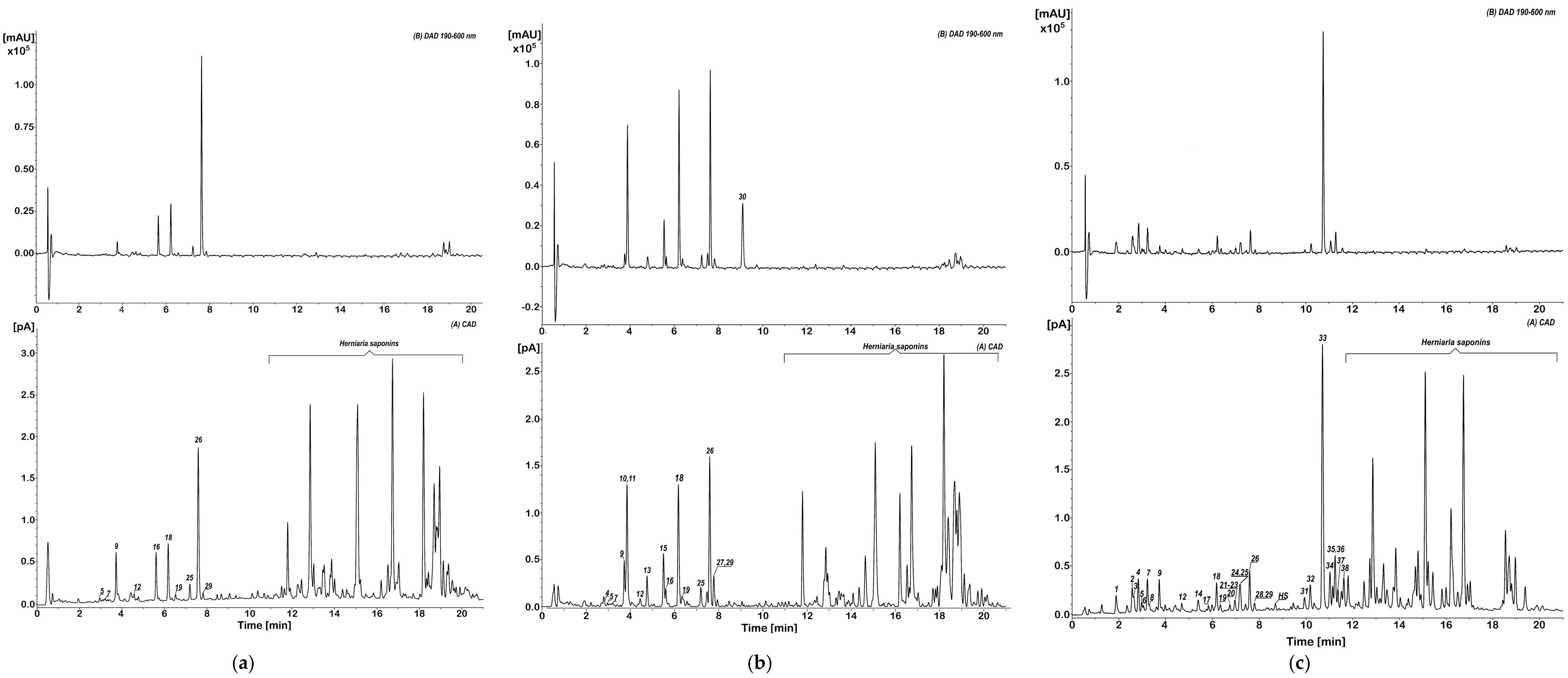
2.1. Phytochemical Profiles and the Content of Individual Phenolics in HG, HP, and HIh Purified Extracts
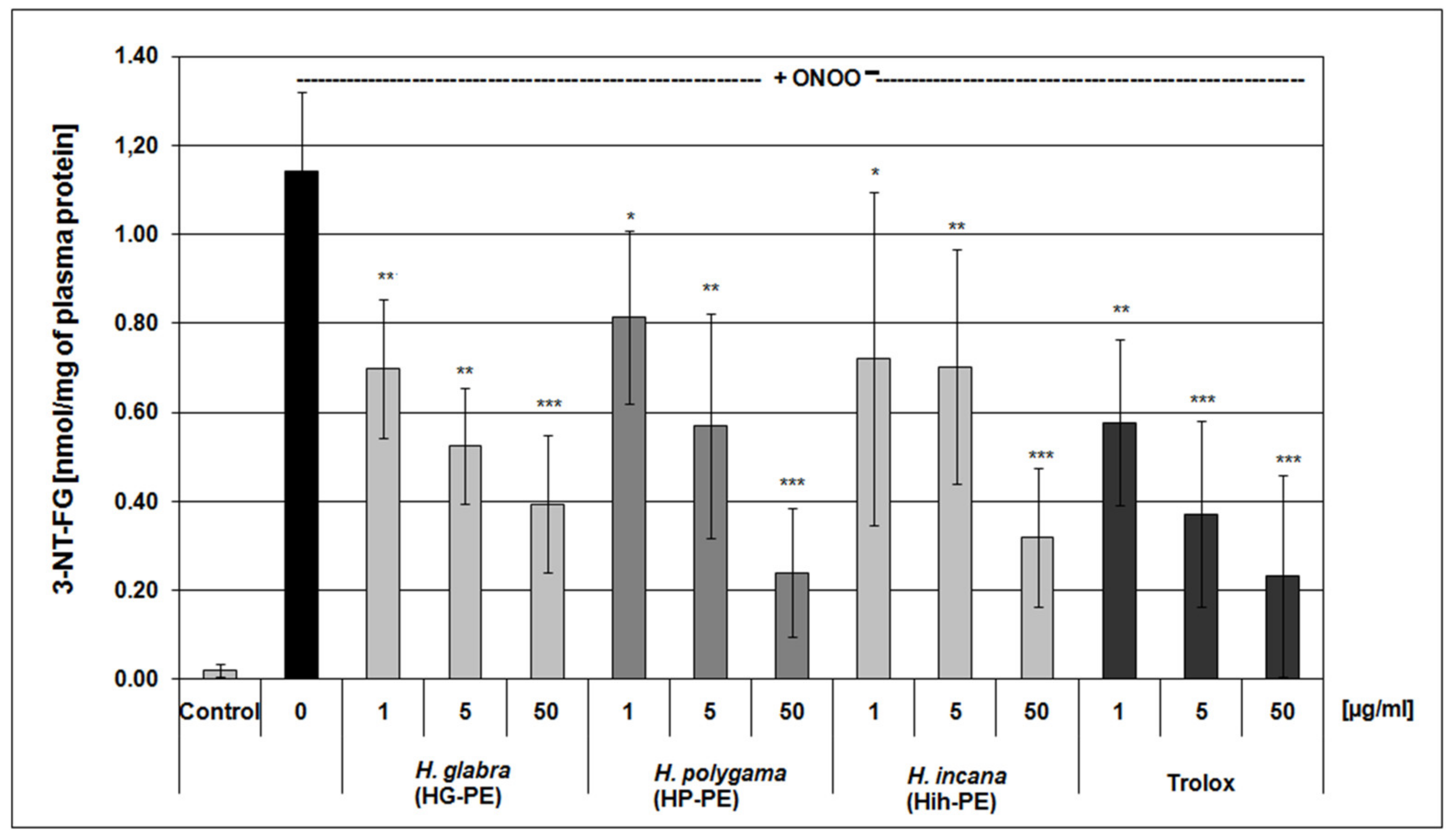
2.2. Evaluation of Antioxidant Effectiveness of the Examined Purified Extracts
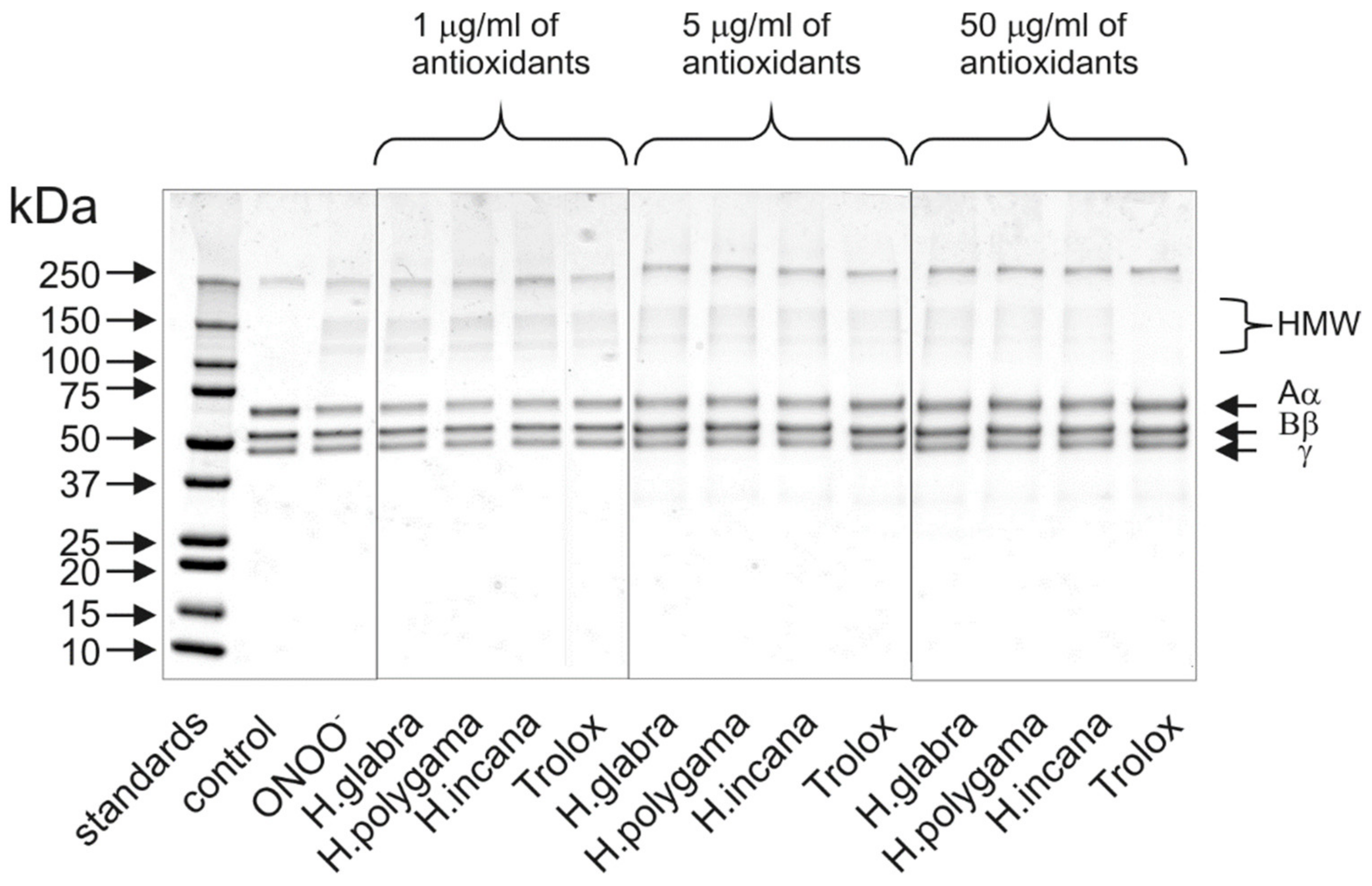
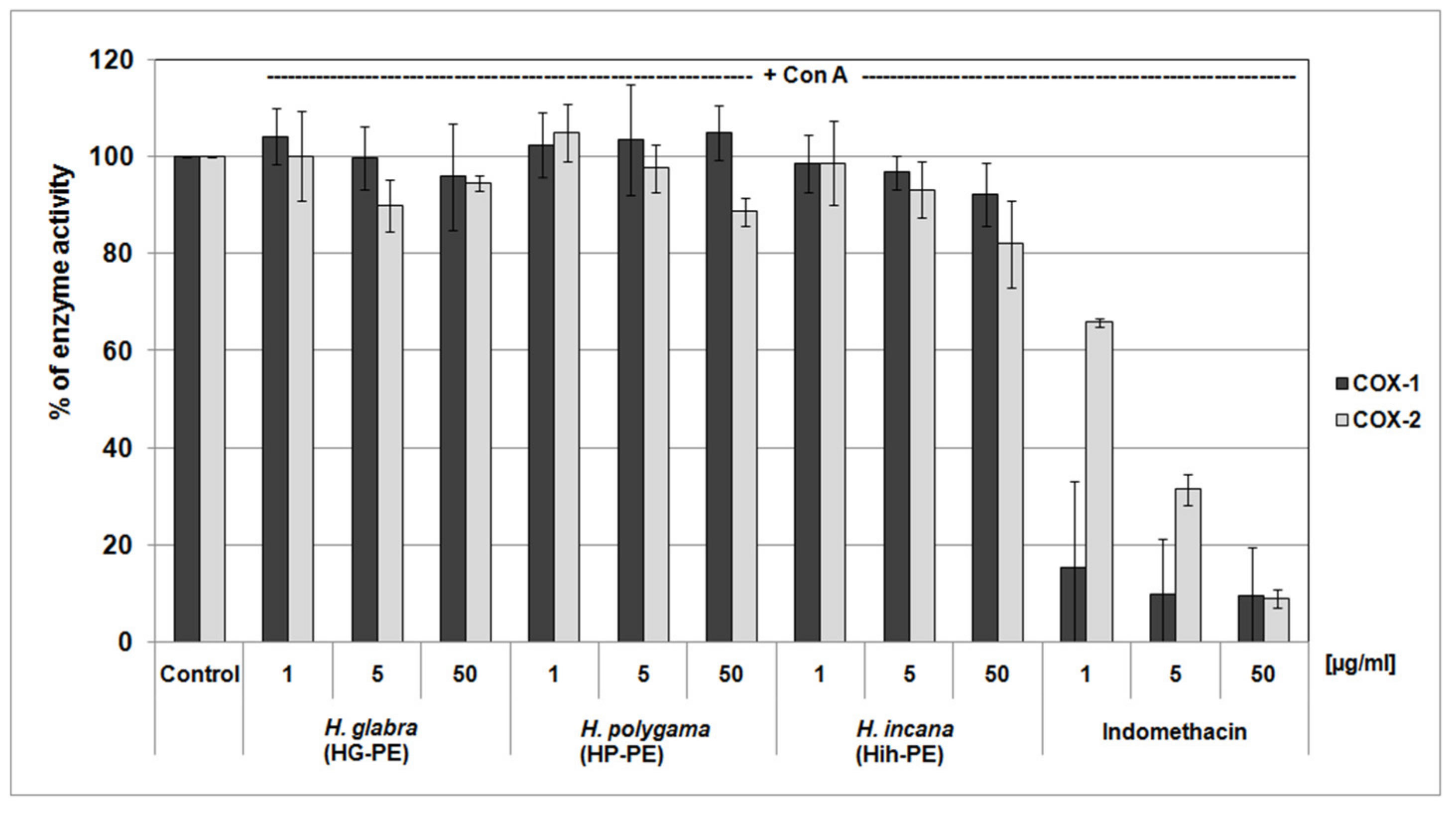
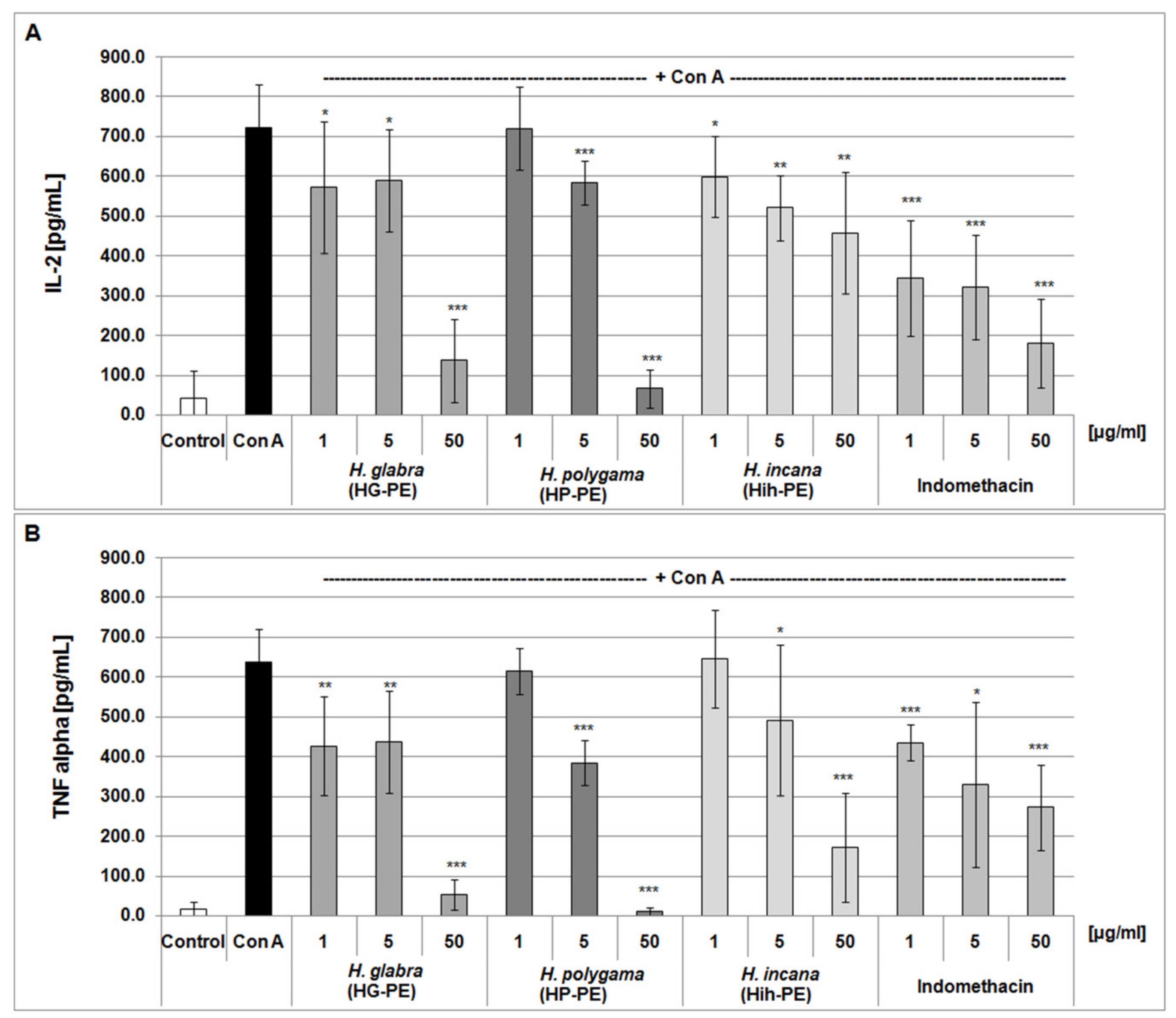
2.3. Anti-Inflammatory Properties of the Tested Herniaria Purified Extracts
2.4. Cytotoxicity Assay
3. Materials and Methods
3.1. Chemicals and Biological Material
3.2. Plant Material
3.3. Extraction
3.4. LC-MS Analysis
3.5. Evaluation of Antioxidant Effects in Blood Plasma
3.6. Evaluation of Anti-Inflammatory Properties
3.7. Cytotoxicity Assays
3.8. Determination of Changes in Fibrinogen Structure by 1D-Electrophoresis
3.9. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Chevallier, A. Encyclopedia of Herbal Medicine. 550 Herbs and Remedies for Common Ailments, 3rd ed.; Dorling Kindersley Ltd.: New York, NY, USA, 2016; ISBN 0241282462. [Google Scholar]
- Kozachok, S.; Pecio, Ł.; Orhan, I.E.; Deniz, F.S.S.; Marchyshyn, S.; Oleszek, W. Reinvestigation of Herniaria glabra L. saponins and their biological activity. Phytochemistry 2020, 169, 112162. [Google Scholar] [CrossRef]
- European Medicines Agency. European Union Herbal Monograph on Herniaria glabra L., H. hirsuta L., H. incana Lam., herba; Committee on Herbal Medicinal Products: Amsterdam, The Netherlands, 2020. [Google Scholar]
- Österreichisches Arzneibuch (Pharmacopoea Austriaca); Verlag Österreich: Wien, Austria, 2017; Volume 217.
- Türk, C.; Neisius, A.; Petrik, A.; Seitz, C.; Skolarikos, A.; Thomas, K.; Davis, N.F.; Donaldson, J.F.; Lombardo, R.; Grivas, N.; et al. European Association of Urology Guidelines on Urolithiasis; European Association of Urology: Arnhem, The Netherlands, 2020. [Google Scholar]
- Trinchieri, A. Epidemiology of urolithiasis: An update Mini-review. Clin. Cases Miner. Bone Metab. 2008, 5, 101–106. [Google Scholar]
- Boonla, C. Oxidative Stress in Urolithiasis. In Reactive Oxygen Species (ROS) in Living Cells; InTech: London, UK, 2018; pp. 129–159. [Google Scholar]
- Wigner, P.; Grębowski, R.; Bijak, M.; Szemraj, J.; Saluk-Bijak, J. The Molecular Aspect of Nephrolithiasis Development. Cells 2021, 10, 1926. [Google Scholar] [CrossRef] [PubMed]
- Ferraro, P.M.; Taylor, E.N.; Gambaro, G.; Curhan, G.C. Soda and other beverages and the risk of kidney stones. Clin. J. Am. Soc. Nephrol. 2013, 8, 1389–1395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ratkalkar, V.N.; Kleinman, J.G. Mechanisms of stone formation. Clin. Rev. Bone Miner. Metab. 2011, 9, 187–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Selvam, R. Calcium oxalate stone disease: Role of lipid peroxidation and antioxidants. Urol. Res. 2002, 30, 35–47. [Google Scholar] [CrossRef]
- Alelign, T.; Petros, B. Kidney Stone Disease: An Update on Current Concepts. Adv. Urol. 2018, 2018, 3068365. [Google Scholar] [CrossRef]
- Rubió, L.; Motilva, M.J.; Romero, M.P. Recent Advances in Biologically Active Compounds in Herbs and Spices: A Review of the Most Effective Antioxidant and Anti-Inflammatory Active Principles. Crit. Rev. Food Sci. Nutr. 2013, 53, 943–953. [Google Scholar] [CrossRef]
- Kozachok, S.; Pecio, Ł.; Kolodziejczyk-czepas, J.; Marchyshyn, S.; Nowak, P.; Mołdoch, J.; Oleszek, W. γ-Pyrone compounds: Flavonoids and maltol glucoside derivatives from Herniaria glabra L. collected in the Ternopil region of the Ukraine. Phytochemistry 2018, 152, 213–222. [Google Scholar] [CrossRef]
- Kolodziejczyk-Czepas, J.; Kozachok, S.; Pecio, Ł.; Marchyshyn, S.; Oleszek, W. Determination of phenolic profiles of Herniaria polygama and Herniaria incana fractions and their In Vitro antioxidant and anti-inflammatory effects. Phytochemistry 2021, 190, 112861. [Google Scholar] [CrossRef] [PubMed]
- Oleszek, W.; Bialy, Z. Chromatographic determination of plant saponins-An update (2002–2005). J. Chromatogr. A 2006, 1112, 78–91. [Google Scholar] [CrossRef]
- Sastre, F.; Ferreira, F.; Pedreschi, F. MALDI-TOF mass spectrometry and reversed-phase HPLC-ELSD chromatography for structural and quantitative studies of major steroid saponins in commercial extracts of Yucca schidigera Roezl. J. Pharm. Biomed. Anal. 2016, 120, 270–282. [Google Scholar] [CrossRef]
- Weber, B.; Herrmann, M.; Hartmann, B.; Joppe, H.; Schmidt, C.O.; Bertram, H.J. HPLC/MS and HPLC/NMR as hyphenated techniques for accelerated characterization of the main constituents in Chamomile (Chamomilla recutita [L.] Rauschert). Eur. Food Res. Technol. 2008, 226, 755–760. [Google Scholar] [CrossRef]
- Ablikim, G.; Bobakulov, K.; Li, J.; Yadikar, N.; Aisa, H.A. Two new glucoside derivatives of truxinic and cinnamic acids from Lavandula angustifolia mill. Nat. Prod. Res. 2019, 35, 2526–2534. [Google Scholar] [CrossRef]
- Wang, S.S.; Zhang, X.J.; Que, S.; Tu, G.Z.; Wan, D.; Cheng, W.; Liang, H.; Ye, J.; Zhang, Q.Y. 3-Hydroxy-3-methylglutaryl flavonol glycosides from Oxytropis falcata. J. Nat. Prod. 2012, 75, 1359–1364. [Google Scholar] [CrossRef] [PubMed]
- Montoro, P.; Teyeb, H.; Masullo, M.; Mari, A.; Douki, W.; Piacente, S. LC-ESI-MS quali-quantitative determination of phenolic constituents in different parts of wild and cultivated Astragalus gombiformis. J. Pharm. Biomed. Anal. 2013, 72, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Jakimiuk, K.; Wink, M.; Tomczyk, M. Flavonoids of the Caryophyllaceae. Phytochem. Rev. 2021. [Google Scholar] [CrossRef]
- Maier, W.; Peipp, H.; Schmidt, J.; Wray, V.; Strack, D. Cell Levels of a Terpenoid Glycoside (Blumenin) and Wall-Bound Phenolics in Some Cereal Mycorrhizas’. Plant Physiol. 2020, 1, 465–470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krzyzanowska-Kowalczyk, J.; Kolodziejczyk-Czepas, J.; Kowalczyk, M.; Pecio, Ł.; Nowak, P.; Stochmal, A. Yunnaneic Acid B, a Component of Pulmonaria officinalis Extract, Prevents Peroxynitrite-Induced Oxidative Stress In Vitro. J. Agric. Food Chem. 2017, 65, 3827–3834. [Google Scholar] [CrossRef]
- Marchelak, A.; Owczarek, A.; Matczak, M.; Pawlak, A.; Kolodziejczyk-Czepas, J.; Nowak, P.; Olszewska, M.A. Bioactivity Potential of Prunus spinosa L. Flower Extracts: Phytochemical Profiling, Cellular Safety, Pro-inflammatory Enzymes Inhibition and Protective Effects Against Oxidative Stress In Vitro. Front. Pharmacol. 2017, 8, 680. [Google Scholar] [CrossRef] [Green Version]
- Mangge, H.; Becker, K.; Fuchs, D.; Gostner, J. Antioxidants, inflammation and cardiovascular disease. World J. Cardiol. 2014, 6, 462. [Google Scholar] [CrossRef]
- Kattoor, A.J.; Pothineni, N.V.K.; Palagiri, D.; Mehta, J.L. Oxidative Stress in Atherosclerosis. Curr. Atheroscler. Rep. 2017, 19, 42. [Google Scholar] [CrossRef]
- Yang, X.; Li, Y.; Li, Y.; Ren, X.; Zhang, X.; Hu, D.; Gao, Y.; Xing, Y.; Shang, H. Oxidative Stress-Mediated Atherosclerosis: Mechanisms and Therapies. Front. Physiol. 2017, 8, 600. [Google Scholar] [CrossRef] [Green Version]
- Ochoa, C.D.; Wu, R.F.; Terada, L.S. ROS signaling and ER stress in cardiovascular disease. Mol. Asp. Med. 2018, 63, 18–29. [Google Scholar] [CrossRef] [PubMed]
- Lahlou, M. The Success of Natural Products in Drug Discovery. Pharmacol. Pharm. 2013, 4, 17–31. [Google Scholar] [CrossRef] [Green Version]
- Newman, D.J.; Cragg, G.M. Natural Products as Sources of New Drugs from 1981 to 2014. J. Nat. Prod. 2016, 79, 629–661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- European Medicines Agency. Assessment Report on Herniaria glabra L., H. hirsuta L., H. incana Lam., Herba; EMA/HMPC/554033/2018; Committee on Herbal Medicinal Products: Amsterdam, The Netherlands, 2020. [Google Scholar]
- Bencheikh, N.; Elbouzidi, A.; Kharchoufa, L.; Ouassou, H.; Merrouni, I.A.; Mechchate, H.; Es-Safi, I.; Hano, C.; Addi, M.; Bouhrim, M.; et al. Inventory of Medicinal Plants Used Traditionally to Manage Kidney Diseases in North-Eastern Morocco: Ethnobotanical Fieldwork and Pharmacological Evidence. Plants 2021, 10, 1966. [Google Scholar] [CrossRef]
- Hakim, E.M.; Sivak, K.V.; Kaukhova, I.E. Evaluation of the diuretic effect of crude ethanol and saponin-rich extracts of Herniaria glabra L. in rats. J. Ethnopharmacol. 2021, 273, 113942. [Google Scholar] [CrossRef] [PubMed]
- Crescenti, A.; Puiggròs, F.; Colomé, A.; Poch, J.A.; Caimari, A.; del Bas, J.M.; Boqué, N.; Arola, L. Antiurolithiasic effect of a plant formulation of Herniaria glabra, Agropyron repens, Equisetum arvense and Sambucus nigra (Herbensurina®) in the prevention of experimentally induced nephrolithiasis in rats. Arch. Españoles Urol. 2015, 68, 739–749. [Google Scholar]
- Elhagali, G.A.M.; Abozeed, A.E.M.; Abd Elnaser, K.A.; Youssif, Y.M. Investigation of bioactive constituents and biological activities of different fractions from Herniaria hemistemon J. Gay. Bull. Sci. 2019, 30, 67–80. [Google Scholar]
- Nielsen, V.G.; Crow, J.P.; Mogal, A.; Zhou, F.; Parks, D.A. Peroxynitrite decreases hemostasis in human plasma In Vitro. Anesth. Analg. 2004, 99, 21–26. [Google Scholar] [CrossRef]
- Wachowicz, B.; Rywaniak, J.Z.; Nowak, P. Apoptotic markers in human blood platelets treated with peroxynitrite. Platelets 2008, 19, 624–635. [Google Scholar] [CrossRef] [PubMed]
- Rubbo, H.; Trostchansky, A.; O’Donnell, V.B. Peroxynitrite-mediated lipid oxidation and nitration: Mechanisms and consequences. Arch. Biochem. Biophys. 2009, 484, 167–172. [Google Scholar] [CrossRef] [PubMed]
- Vara, D.; Pula, G. Reactive oxygen species: Physiological roles in the regulation of vascular cells. Curr. Mol. Med. 2014, 14, 1103–1125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- ILL-Raga, G.; Palomer, E.; Ramos-Fernández, E.; Guix, F.X.; Bosch-Morató, M.; Guivernau, B.; Tajes, M.; Valls-Comamala, V.; Jiménez-Conde, J.; Ois, A.; et al. Fibrinogen nitrotyrosination after ischemic stroke impairs thrombolysis and promotes neuronal death. Biochim. Biophys. Acta 2015, 1852, 421–428. [Google Scholar] [CrossRef] [PubMed]
- Bartesaghi, S.; Radi, R. Fundamentals on the biochemistry of peroxynitrite and protein tyrosine nitration. Redox Biol. 2018, 14, 618–625. [Google Scholar] [CrossRef]
- Beckman, J.S.; Chen, J.; Ischiropoulos, H.; Crow, J.P. Oxidative chemistry of peroxynitrite. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1994; Volume 233, pp. 229–240. ISBN 9780121821340. [Google Scholar]
- Vadseth, C.; Souza, J.M.; Thomson, L.; Seagraves, A.; Nagaswami, C.; Scheiner, T.; Torbet, J.; Vilaire, G.; Bennett, J.S.; Murciano, J.-C.; et al. Pro-thrombotic State Induced by Post-translational Modification of Fibrinogen by Reactive Nitrogen Species. J. Biol. Chem. 2004, 279, 8820–8826. [Google Scholar] [CrossRef] [Green Version]
- Nowak, P.; Zbikowska, H.M.; Ponczek, M.; Kolodziejczyk, J.; Wachowicz, B. Different vulnerability of fibrinogen subunits to oxidative/nitrative modifications induced by peroxynitrite: Functional consequences. Thromb. Res. 2007, 121, 163–174. [Google Scholar] [CrossRef]
- Štikarová, J.; Kotlín, R.; Riedel, T.; Suttnar, J.; Pimková, K.; Chrastinová, L.; Dyr, J.E. The Effect of Reagents Mimicking Oxidative Stress on Ffibrinogen Function. Sci. World J. 2013, 2013, 359621. [Google Scholar] [CrossRef] [Green Version]
- De Vries, J.J.; Snoek, C.J.M.; Rijken, D.C.; De Maat, M.P.M. Effects of Post-Translational Modifications of Fibrinogen on Clot Formation, Clot Structure, and Fibrinolysis: A Systematic Review. Arterioscler. Thromb. Vasc. Biol. 2020, 40, 554–569. [Google Scholar] [CrossRef]
- Nowak, P.; Kołodziejczyk, J.; Wachowicz, B. Peroxynitrite and fibrinolytic system: The effect of peroxynitrite on plasmin activity. Mol. Cell. Biochem. 2004, 267, 141–146. [Google Scholar] [CrossRef]
- Kolodziejczyk-Czepas, J.; Ponczek, M.B.; Nowak, P. Peroxynitrite and fibrinolytic system—The effects of peroxynitrite on t-PA-induced plasmin activity. Int. J. Biol. Macromol. 2015, 81, 212–219. [Google Scholar] [CrossRef]
- Manach, C.; Williamson, G.; Morand, C.; Scalbert, A.; Rémésy, C. Bioavailability and bioefficacy of polyphenols in humans. I. Review of 97 bioavailability studies. Am. J. Clin. Nutr. 2005, 81, 230S–242S. [Google Scholar] [CrossRef] [Green Version]
- Boyle, S.P.; Dobson, V.L.; Duthie, S.J.; Hinselwood, D.C.; Kyle, J.A.M.; Collins, A.R. Bioavailability and efficiency of rutin as an antioxidant: A human supplementation study. Eur. J. Clin. Nutr. 2000, 54, 774–782. [Google Scholar] [CrossRef]
- Ganeshpurkar, A.; Saluja, A.K. The Pharmacological Potential of Rutin. Saudi Pharm. J. 2017, 25, 149–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Enogieru, A.B.; Haylett, W.; Hiss, D.C.; Bardien, S.; Ekpo, O.E. Rutin as a Potent Antioxidant: Implications for Neurodegenerative Disorders. Oxid. Med. Cell. Longev. 2018, 2018, 6241017. [Google Scholar] [CrossRef] [PubMed]
- Obolskiy, D.; Pischel, I.; Feistel, B.; Glotov, N.; Heinrich, M. Artemisia dracunculus L. (tarragon): A critical review of its traditional use, chemical composition, pharmacology, and safety. J. Agric. Food Chem. 2011, 59, 11367–11384. [Google Scholar] [CrossRef] [Green Version]
- Shimizu, T.; Shibuya, N.; Narukawa, Y.; Oshima, N.; Hada, N.; Kiuchi, F. Synergistic effect of baicalein, wogonin and oroxylin A mixture: Multistep inhibition of the NF-κB signalling pathway contributes to an anti-inflammatory effect of Scutellaria root flavonoids. J. Nat. Med. 2018, 72, 181–191. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Zhao, S.; Li, F.; Zhang, B.; Qu, Y.; Sun, T.; Luo, T.; Li, D. Investigation of antioxidant interactions between radix Astragali and Cimicifuga foetida and identification of synergistic antioxidant compounds. PLoS ONE 2014, 9, e87221. [Google Scholar] [CrossRef]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.C. NF-κB signaling in inflammation. Signal Transduct. Target. Ther. 2017, 2, 17023. [Google Scholar] [CrossRef] [Green Version]
- Choi, S.; Lim, T.G.; Hwang, M.K.; Kim, Y.A.; Kim, J.; Kang, N.J.; Jang, T.S.; Park, J.S.; Yeom, M.H.; Lee, K.W. Rutin inhibits B [a] PDE-induced cyclooxygenase-2 expression by targeting EGFR kinase activity. Biochem. Pharmacol. 2013, 86, 1468–1475. [Google Scholar] [CrossRef]
- Feng, R.; Lu, Y.; Bowman, L.L.; Qian, Y.; Castranova, V.; Ding, M. Inhibition of Activator Protein-1, NF-κB, and MAPKs and Induction of Phase 2 Detoxifying Enzyme Activity by Chlorogenic Acid. J. Biol. Chem. 2005, 280, 27888–27895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, M.; Choi, S.Y.; Lee, P.; Hur, J. Neochlorogenic Acid Inhibits Lipopolysaccharide-Induced Activation and Pro-inflammatory Responses in BV2 Microglial Cells. Neurochem. Res. 2015, 40, 1792–1798. [Google Scholar] [CrossRef]
- Molimau-Samasoni, S.; Woolner, V.H.; Su’emalo, T.F.; Robichon, K.; Patel, V.; Andreassend, S.K.; Sheridan, J.P.; Kawa, T.T.; Gresham, D.; Miller, D.; et al. Functional genomics and metabolomics advance the ethnobotany of the Samoan traditional medicine “matalafi.”. Proc. Natl. Acad. Sci. USA 2021, 118, e2100880118. [Google Scholar] [CrossRef] [PubMed]
- Naveed, M.; Hejazi, V.; Abbas, M.; Kamboh, A.A.; Khan, G.J.; Shumzaid, M.; Ahmad, F.; Babazadeh, D.; FangFang, X.; Modarresi-Ghazani, F.; et al. Chlorogenic acid (CGA): A pharmacological review and call for further research. Biomed. Pharmacother. 2018, 97, 67–74. [Google Scholar] [CrossRef]
- Hwang, S.J.; Kim, Y.W.; Park, Y.; Lee, H.J.; Kim, K.W. Anti-inflammatory effects of chlorogenic acid in lipopolysaccharide- stimulated RAW 264.7 cells. Inflamm. Res. 2014, 63, 81–90. [Google Scholar] [CrossRef]
- Kadioglu, O.; Nass, J.; Saeed, M.E.M.; Schuler, B.; Efferth, T. Kaempferol Is an Anti-Inflammatory Compound with Activity towards NF-κB Pathway Proteins. Anticancer Res. 2015, 35, 2645–2650. [Google Scholar]
- Yu, L.; Chen, C.; Wang, L.F.; Kuang, X.; Liu, K.; Zhang, H.; Du, J.-R. Neuroprotective Effect of Kaempferol Glycosides against Brain Injury and Neuroinflammation by Inhibiting the Activation of NF-κB and STAT3 in Transient Focal Stroke. PLoS ONE 2013, 8, e55839. [Google Scholar] [CrossRef] [PubMed]
- El-Aasr, M.; Kabbash, A.; Abo El-Seoud, K.A.; Al-Madboly, L.A.; Ikeda, T. Antimicrobial and Immunomodulatory Activities of Flavonol Glycosides Isolated from Atriplex halimus L. Herb. J. Pharm. Sci. Res. 2016, 8, 1159–1168. [Google Scholar]
- Chen, L.; Mola, M.; Deng, X.; Mei, Z.; Huang, X.; Shu, G.; Wei, L.; Hou, X.; Lan, Z.; Lin, Q. Dolichos falcata Klein attenuated the inflammation induced by monosodium urate crystals in vivo and In Vitro. J. Ethnopharmacol. 2013, 150, 545–552. [Google Scholar] [CrossRef] [PubMed]
- Vo, N.N.Q.; Nomura, Y.; Muranaka, T.; Fukushima, E.O. Structure-Activity Relationships of Pentacyclic Triterpenoids as Inhibitors of Cyclooxygenase and Lipoxygenase Enzymes. J. Nat. Prod. 2019, 82, 3311–3320. [Google Scholar] [CrossRef] [PubMed]
- Pryor, W.A.; Cueto, R.; Jin, X.; Koppenol, W.H.; Ngu-Schwemlein, M.; Squadrito, G.L.; Uppu, P.L.; Uppu, R.M. A practical method for preparing peroxynitrite solutions of low ionic strength and free of hydrogen peroxide. Free Radic. Biol. Med. 1995, 18, 75–83. [Google Scholar] [CrossRef]
- Doolittle, R.F.; Schubert, D.; Schwartz, S.A. Amino acid sequence studies on artiodactyl fibrinopeptides: I. Dromedary camel, mule deer, and cape buffalo. Arch. Biochem. Biophys. 1967, 118, 456–467. [Google Scholar] [CrossRef]
- Wachowicz, B.; Kustroń, J. Effect of cisplatin on lipid peroxidation in pig blood platelets. Cytobios 1992, 70, 41–47. [Google Scholar]
- Olas, B.; Nowak, P.; Kolodziejczyk, J.; Ponczek, M.; Wachowicz, B. Protective effects of resveratrol against oxidative/nitrative modifications of plasma proteins and lipids exposed to peroxynitrite. J. Nutr. Biochem. 2006, 17, 96–102. [Google Scholar] [CrossRef]
- Rice-Evans, C.A.; Diplock, A.T.; Symons, M.C.R. Detection of protein structural modifications induced by free radicals. In Laboratory Techniques in Biochemistry and Molecular Biology; Burdon, R.H., van Knippenberg, P.H., Eds.; Elsevier: Amsterdam, The Netherlands; pp. 207–235. ISBN 9780444813046.
- Kolodziejczyk-Czepas, J.; Nowak, P.; Kowalska, I.; Stochmal, A. Biological activity of clovers—Free radical scavenging ability and antioxidant action of six Trifolium species. Pharm. Biol. 2014, 52, 1308–1314. [Google Scholar] [CrossRef]
- Strober, W. Trypan Blue Exclusion Test of Cell Viability. In Current Protocols in Immunology; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2001; p. A-3B. ISBN 9780471142737. [Google Scholar]
- Laemmli, U.K. Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| tR (min) | Compound | Neutral Formula | Mode | m/z | Δ * | Mσ ** | Major Fragments (%) | HG,% | HP,% | HIh,% | Ref | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 1.89 | 3-CQA (Neochlorogenic acid) | C16H18O9 | [M − H]− [2M − H]− | 353.0872 707.1820 | 1.6 0.9 | 1.5 1.3 | 191.0558 (100) (C7H11O6), 179.0342 (80.5) (C9H7O4), 173.0342 (3.5) (C7H9O5), 161.0236 (4.8) (C9H5O3), 135.0439 (14.1) (C8H7O2) | _ | _ | 1.27 | [15] |
| 2 | 2.56 | Cis-3-pCoQA | C16H18O8 | [M − H]− [2M − H]− | 337.0923 675.1926 | 1.6 0.74 | 6.8 9.3 | 191.0559 (7.1) (C7H11O6), 173.0453 (4.1) (C7H9O5), 163.0390 (100) (C9H7O3), 155.0336 (2.3) (C7H7O4), 119.0487 (8.6) (C8H7O) | _ | _ | 1.51 | [15] |
| 3 | 2.64 | Trans-3-pCoQA | C16H18O8 | [M − H]− [2M − H]− | 337.0921 675.1919 | 2.5 1.8 | 4.3 8.6 | 191.0558 (37.6) (C7H11O6), 173.0449 (4.4) (C7H9O5), 163.0392 (100) (C9H7O3), 155.0343 (2.0) (C7H7O4), 119.0489 (9.2) (C8H7O) | _ | _ | 0.31 | [15] |
| 4 | 2.81 | a 5-CQA (Chlorogenic acid) | C16H18O9 | [M − H]− [2M − H]− | 353.0871 707.1822 | 0.7 1.0 | 1.9 23.5 | 191.0556 (100) (C7H11O6), 161.0244 (1.53) (C9H5O3) | _ | TA | 1.37 | [15] |
| 5 | 2.99 | pCoA-4-O-hexoside | C15H18O8 | [M − H]− [2M − H]− | 325.0923 651.1925 | 1.8 0.8 | 9.7 11.4 | 163.0393 (100) (C9H7O3), 119.0490 (12.6) (C8H7O) | TA | TA | 0.12 | [15] |
| 6 | 3.05 | a 4-CQA (Cryptochlorogenic acid) | C16H18O9 | [M − H]− [2M − H]− | 353.0874 707.1829 | 1.2 −0.1 | 3.6 15.3 | 191.0552 (40.4) (C7H11O6), 179.0343 (84.5) (C9H7O4), 173.0446 (100) (C7H9O5), 161.0231 (2.7) (C9H5O3), 155.0330 (5.3) (C7H7O4), 135.0441 (13.9) (C8H7O2) | _ | _ | 0.20 | [15] |
| 7 | 3.21 | Cis-3-FerQA | C17H20O9 | [M − H]− [2M − H]− | 367.1025 735.2128 | 2.6 1.9 | 4.0 15.7 | 193.0500 (100) (C10H9O4), 173.0450 (4.0) (C7H9O5), 155.0343 (1.2) (C7H7O4), 149.0597 (2.0) (C9H9O2), 134.0363 (C8H6O2●) (6.1) | TA | TA | 1.24 | [15] |
| 8 | 3.31 | Trans-3-FerQA | C17H20O9 | [M − H]− [2M − H]− | 367.1030 735.2138 | 1.2 0.6 | 6.7 36.2 | 193.0502 (100) (C10H9O4), 173.0445 (5.5) (C7H9O5), 155.0338 (1.0) (C7H7O4), 149.0599 (4.3) (C9H9O2), 134.0365 (11.8) (C8H6O2●) | _ | _ | 0.16 | [15] |
| 9 | 3.72 | # Licoagroside B (maltol-3-O-[HMG-(1→6)]-β-glucoside) | C18H24O12 | [M − H]− [2M − H]− | 431.1189 863.2463 | 1.3 0.9 | 4.3 7.2 | 161.0448 (10) (C6H9O5), 125.0232 (100) (C6H5O3) | 1.80 | 1.33 | 1.10 | [14] |
| 10 | 3.81 | # (2R,3R)-Benzoyl-tartaric acid | C11H10O7 | [M − H]− | 253.0340 | 5.4 | 6.8 | 130.9969 (26.3) (C4H3O5), 121.0277 (100) (C7H5O2) | _ | 1.01 | _ | [15] |
| 11 | 3.87 | #Cis-2-hydroxy-4-methoxycinnamic acid 2-O-β-glucoside (cis-GMCA) | C16H20O9 | [M − H]− | 355.1017 | 5.2 | 4.9 | 193.0494 (100) (C10H9O4), 149.0591 (75.4) (C9H9O2), 134.0367 (5.5) (C8H6O2●) | _ | 5.76 | _ | [15] |
| 12 | 4.70 | Trans-4-FerQA | C17H20O9 | [M − H]− | 367.1028 | 1.9 | 4.1 | 193.0501 (21.3) (C10H9O4), 191.0558 (40.6) (C7H11O6), 173.0455 (100) (C7H9O5), 155.0349 (4.0) (C7H7O4), 134.0372 (2.5) (C8H6O2●) | TA | TA | 0.22 | [15] |
| 13 | 4.77 | # 3-(2-glucosyloxy-4- methoxyphenyl)propanoic acid (GMPPA) | C16H22O9 | [M − H]− | 357.1174 | 4.9 | 0.7 | 195.0650 (100) (C10H11O4), 177.0541 (2.8) (C10H9O3), 151.0748 (10.7) (C9H11O2), 136.0512 (1.1) (C8H8O2●) | _ | 1.07 | _ | [15] |
| 14 | 5.41 | N-Malonyl-tryptophan | C14H14N2O5 | [M + H]+ | 291.0969 | 2.3 | 4.6 | 273.0868 (19.0) (C14H13N2O4), 245.0916 (100) (C13H13N2O3), 227.0807 (24.9) (C13H11N2O2), 188.0702 (21.9) (C11H10NO2), 130.0647 (18.7) (C9H8N) | _ | _ | NC | [15] |
| 15 | 5.52 | #Trans-2-hydroxy-4- methoxycinnamic acid 2-O-β-glucoside (trans-GMCA) | C16H20O9 | [M − H]− | 355.1016 | 5.3 | 0.7 | 193.0494 (100) (C10H9O4), 149.0591 (36.5) (C9H9O2), 134.0357 (4.7) (C8H6O2●) | _ | 1.58 | _ | [15] |
| 16 | 5.60 | # Apiorutin (quercetin-3-O-[β-Api-(1→2)]-[α-Rha-(1→6)]-β-glucoside) | C32H38O20 | [M − H]− | 741.1868 | 2.1 | 5.9 | 609.1461 (0.5) (C27H29O16), 591.1362 (0.3) (C27H27O15), 300.0269 (100) (C15H8O7●), 271.0244 (4.5) (C14H7O6), 255.0299 (2.0) (C14H7O5), 178.9986 (2.0) (C8H3O5), 151.0028 (0.9) (C7H3O4) | 1.50 | 0.33 | _ | [14] |
| 17 | 5.83 | Kaempferol-3-O-(dHex-(1→2))-[dHex-(1→2)]- hexoside | C33H40O19 | [M − H]− | 739.2080 | 1.5 | 29.0 | 575.1392 (0.6) (C27H27O14), 284.0316 (100) (C15H8O6●), 255.0291 (7.4) (C14H7O5), 227.0341 (4.5) (C13H7O4), 178.9973 (1.2) (C8H3O5), 151.0024 (1.5) (C7H3O4) | _ | _ | 0.16 | [15] |
| 18 | 6.18 | # Rutin (quercetin-3-O-[α-Rha-(1→6)]-β-glucoside) | C27H30O16 | [M − H]− | 609.1453 | 1.5 | 8.3 | 343.0459 (1.7) (C17H11O8), 300.0268 (100) (C15H8O7●), 255.0299 (1.0) (C14H7O5), 178.9986 (2.9) (C8H3O5), 151.0037 (1.8) (C7H3O4) | 1.87 | 5.27 | 0.82 | [14] |
| 19 | 6.33 | Quercetin-3-O- hexoside | C21H20O12 | [M − H]− | 463.0874 | 1.6 | 4.8 | 343.0459 (1.4) (C17H11O8), 300.0267 (100) (C15H8O7●), 255.0299 (1.1) (C14H7O5), 178.9965 (1.8) (C8H3O5), 151.0035 (1.5) (C7H3O4) | TA | 0.22 | 0.18 | |
| 20 | 6.76 | Kaempferol-3-O-[dHex-(1→6)]- hexoside | C27H30O15 | [M − H]− | 593.1499 | 1.3 | 2.1 | 327.0502 (3.2) (C17H11O7), 285.0389 (53.9) (C15H9O6), 284.0317 (100) (C15H8O6●), 255.0300 (2.9) (C14H7O5), 227.0343 (1.7) (C13H7O4), 151.0035 (2.1) (C7H3O4) | _ | _ | 0.19 | [15] |
| 21 | 6.95 | Kaempferol-3-O-hexoside | C21H20O11 | [M − H]− | 447.0902 | 2.0 | 6.5 | 284.0322 (47.4) (C15H8O6●) | _ | _ | TA | [15] |
| 22 | 6.96 | Quercetin- 3-O-[HMG]- hexoside | C27H28O16 | [M − H]− | 607.1219 | 2.4 | 7.2 | 545.1280 (8.0) (C26H25O13), 505.0990 (37.2) (C23H21O13), 463.0874 (47.5) (C21H19O12), 300.0268 (100) (C15H8O7●), 271.0235 (2.9) (C14H7O6), 255.0292 (1.5) (C14H7O5), 178.9978 (3.1) (C8H3O5), 151.0028 (2.6) (C7H3O4) | _ | _ | TA | [15] |
| 23 | 7.00 | Blumenin isomer (sesquiterpenoid cyclohexanone-hexuronosyl-hexoside) | C25H40O13 | [M − H]− | 547.2383 | 1.3 | 12.3 | 347.2062 (3.7) (C19H31O7), 209.1529 (1.7) (C13H21O2), 175.0239 (4.3) (C6H7O6), 161.0444 (1.5) (C6H9O5) | _ | _ | TA | [23] |
| 24 | 7.17 | Kaempferol-3-O-[HMG-(1→3/4)]-[dHex-(1→2)]-hexoside | C33H38O19 | [M − H]− | 737.1919 | 2.1 | 1.5 | 675.1922 (1.4) (C32H35O16), 635.1602 (5.7) (C29H31O16), 593.1528 (8.1) (C27H29O15), 429.0819 (0.7) (C21H17O10), 284.0317 (100) (C15H8O6●), 255.0293 (4.4) (C14H7O5), 227.0340 (2.0) (C13H7O4), 178.9974 (0.8) (C8H3O5), 151.0027 (0.9) (C7H3O4) | _ | _ | 1.22 $ | [15] |
| 25 | 7.19 | Nicotiflorin (kaempferol-3-O-[Rha-(1→6)]-glucoside) | C27H30O15 | [M − H]− | 593.1501 | 1.8 | 4.4 | 327.0498 (2.4) (C17H11O7), 285.0392 (100) (C15H9O6), 255.0291 (1.9) (C14H7O5), 227.0344 (0.9) (C13H7O4), 151.0027 (1.2) (C7H3O4) | 0.44 | 0.44 | [14] | |
| 26 | 7.58 | # Narcissin (isorhamnetin-3-O-[α-Rhap-(1→6)]-β-glucoside) | C28H32O16 | [M − H]− | 623.1603 | 2.3 | 5.1 | 357.0609 (1.5) (C18H13O8), 315.0499 (100) (C16H11O7), 300.0266 (5.9) (C15H8O7●), 271.0244 (1.3) (C14H7O6), 151.0028 (0.7) (C7H3O4) | 7.34 | 5.43 | 0.96 | [14] |
| 27 | 7.77 | 4,4′-Dimethoxy-2,2′-di-O-hexoside-truxinate (cyclodimer of GMCA) | C32H40O18 | [M − H]− | 711.2128 | 1.9 | 6. 9 | 549.1606 (12.9) (C26H29O13), 531.1497 (8.5) (C26H27O12), 369.0973 (51.4) (C20H17O7), 343.1181 (41.6) (C19H19O6), 299.1284 (23.7) (C18H19O4), 271.0970 (92.1), 256.0737 (2.7) (C15H12O4●), 193.0502 (100) (C10H9O4), 149.0599 (41.8) (C9H9O2), 134.0361 (8.2) (C8H6O2●) | _ | TA | _ | [15] |
| 28 | 7.80 | Kaempferol-3-O-[HMG-(1→3/4)]-hexoside | C27H28O15 | [M − H]− | 591.1344 | 1.9 | 5.6 | 529.1343 (13.8) (C26H25O12), 489.1031 (39.2) (C23H21O12), 447.0920 (31.8) (C21H19O11), 327.0499 (1.8) (C17H11O7), 284.0318 (100) (C15H8O6●), 255.0282 (1.4) (C14H7O5), 229.0503 (1.1) (C13H9O4), 151.0025 (0.9) (C7H3O4) | _ | _ | 0.25 | |
| 29 | 7.81 | Isorhamnetin-3-O-hexoside | C22H22O12 | [M − H]− | 477.1030 | 1.7 | 12.8 | 357.0604 (3.0) (C18H13O8), 314.0422 (100) (C16H10O7●), 299.0195 (1.46) (C15H7O7), 285.0397 (1.3) (C15H9O6), 271.0220 (1.6) (C14H7O6), 151.0034 (2.0) (C7H3O4) | TA | TA | TA | |
| 30 | 9.10 | a Herniarin (7-methoxy- coumarin) | C10H8O3 | [M + H]+ | 177.0557 | −5.8 | 1.9 | 177.0537 (100), 133.0659 (4. 3), 121.0658 (2.9) | _ | 1.59 | _ | [15] |
| 31 | 9.94 | # 7-O-Methylquercetin-3-O-[HMG-(1→6)]-[α-Rha-(1→2)]-β-galactoside | C34H40O20 | [M − H]− | 767.2035 | 0.7 | 17.0 | 705.2059 (0.4) (C33H37O17), 665.1726 (1.8) (C30H33O17), 623.1619 (4.8) (C28H31O16), 314.0429 (100) (C16H10O7●); 299.0196 (13.1) (C15H7O7), 271.0246 (2.4) (C14H7O6), 193.0145 (0.8) (C9H5O5), 165.0189 (1.5) (C8H5O4) | _ | _ | 0.31 | [15] |
| 32 | 10.20 | 7-O-Methylkaempferol-3-O-[dHex-(1→2)]-hexoside | C28H32O15 | [M − H]− | 607.1672 | −0.6 | 1.6 | 298.0484 (100) (C16H10O6●), 283.0248 (10.2) (C15H7O6), 271.0610 (0.9) (C15H11O5), 255.0298 (1.1) (C14H7O5), 165.0190 (0.7) (C8H5O4) | _ | _ | 0.51 | [15] |
| 33 | 10.72 | # Oxytroflavoside A (7-O-methylkaempferol-3-O-[HMG-(1→6)]-[α-Rha-(1→2)]-β- galactoside) | C34H40O19 | [M − H]− | 751.2089 | 0.3 | 14.8 | 649.1772 (2.2) (C30H33O16), 607.1666 (4.4) (C28H31O15), 298.0481 (100) (C16H10O6●), 283.0247 (13.5) (C15H7O6); 271.0610 (0.9) (C15H11O5), 255.0298 (1.7) (C14H7O5), 165.0186 (0.7) (C8H5O4) | _ | _ | 14.73 | [15] |
| 34 | 11.05 | # 7-O-Methylkaempferol-3-O-[HMG-(1→6)]-[α-Rha-(1→2)]-β-glucoside | C34H40O19 | [M − H]− | 751.2090 | 0.2 | 4.4 | 649.1788 (1.4) (C30H33O16), 607.1670 (5.5) (C28H31O15), 298.0483 (100) (C16H10O6●), 283.0247 (13.3) (C15H7O6), 271.0616 (0.8) (C15H11O5), 255.0301 (1.5) (C14H7O5), 165.0188 (0.8) (C8H5O4) | _ | _ | 0.91 | [15] |
| 35 | 11.26 | # Oxytroflavoside C (main) (7-O-methylkaempferol-3-O-[HMG-(1→4)]-[α-Rha-(1→2)]-β-galactoside]) Oxytroflavoside B (minor) (7-O-methylkaempferol-3-O-[HMG-(1→3)]-[α-Rha-(1→2)]-β-galactoside) | C34H40O19 | [M − H]− | 751.2086 | 0.6 | 8.1 | 649.1779 (1.6) (C30H33O16), 607.1676 (8.8) (C28H31O15), 298.0483 (100) (C16H10O6●), 283.0249 (11.2) (C15H7O6), 271.0614 (1.0) (C15H11O5), 255.0303 (1.2) (C14H7O5), 165.0189 (0.7) (C8H5O4) | _ | _ | 1.38 $ | [15] |
| 36 | ||||||||||||
| 37 | 11.54 | 7-O-Methyl-kaempferol-3-O-[HMG-(1→3/4)]-hexoside | C28H30O15 | [M − H]− | 605.1515 | −0.5 | 11.4 | 543.1516 (8.0) (C27H27O12), 503.1198 (27.1) (C24H23O12), 461.1093 (22.9) (C22H21O11), 341.0663 (1.8) (C18H13O7), 298.0482 (100) (C16H10O6●), 283.0247 (6.2) (C15H7O6); 255.0302 (1.0) (C14H7O5), 165.0192 (1.3) (C8H5O4) | _ | _ | 0.32 | [15] |
| 38 | 11.77 | 7-O-Methyl-kaempferol-3-O-[dHex-(1→2)]-pentoside | C27H30O14 | [M − H]− | 577.1565 | −0.5 | 13.5 | 413.0888 (1.0) (C21H17O9), 298.0490 (100) (C16H10O6●), 283.0250 (7.3) (C15H7O6), 255.0302 (1.0) (C14H7O5), 165.0188 (0.9) (C8H5O4) | _ | _ | 0.12 | [15] |
| Sum | 12.95 | 24.03 | 29.56 |
| (µg/ mL) | TBARS (nmol/mL of Plasma) (n = 9) | Protein-SH Groups (µmol/mL of Plasma) (n = 8) | FRAP (mM Fe2+) (n = 9) | |
|---|---|---|---|---|
| Control (untreated) plasma | 0 | 0.078 ± 0.011 | 0.335 ± 0.019 | 0.460 ± 0.035 |
| Plasma treated with ONOO− | 0 | 0.122 ± 0.015 # | 0.229 ± 0.029 ## | 0.380 ± 0.027 ## |
| Trolox | 1 | 0.104 ± 0.027 | 0.267 ± 0.025 * | 0.422 ± 0.031 |
| 5 | 0.108 ± 0.027 | 0.274 ± 0.029 ** | 0.462 ± 0.075 ** | |
| 50 | 0.106 ± 0.025 | 0.288 ± 0.038 ** | 0.603 ± 0.060 ** | |
| HG-PE | 1 | 0.100 ± 0.022 * | 0.260 ± 0.026 * | 0.418 ± 0.033 |
| 5 | 0.099 ± 0.023 ** | 0.268 ± 0.022 ** | 0.429 ± 0.028 * | |
| 50 | 0.102 ± 0.022 ** | 0.262 ± 0.024 ** | 0.447 ± 0.040 * | |
| HP-PE | 1 | 0.101 ± 0.022 * | 0.261 ± 0.023 * | 0.428 ± 0.029 |
| 5 | 0.093 ± 0.019 ** | 0.264 ± 0.029 * | 0.442 ± 0.033 ** | |
| 50 | 0.100 ± 0.026 * | 0.258 ± 0.027 * | 0.458 ± 0.032 ** | |
| HIh-PE | 1 | 0.096 ± 0.026 * | 0.258 ± 0.031 * | 0.425 ± 0.033 |
| 5 | 0.101 ± 0.024 | 0.262 ± 0.024 ** | 0.440 ± 0.028 * | |
| 50 | 0.099 ± 0.022 * | 0.264 ± 0.025 ** | 0.485 ± 0.091 ** |
| The Examined Extracts | (μg/mL) | PBMCs Viability (%) | |
|---|---|---|---|
| the Resazurin-Based Assay (n = 7) | the Trypan Blue Excluding Test (n = 11) | ||
| HG-PE | 5 | 93.47 ± 6.83 | 102.73 ± 11.15 |
| 25 | 95.42 ± 6.82 | 97.49 ± 8.25 | |
| 50 | 93.82 ± 12.51 | 89.22 ± 14.37 | |
| HP-PE | 5 | 93.11 ± 10.27 | 99.06 ± 11.36 |
| 25 | 95.17 ± 8.46 | 97.83 ± 10.97 | |
| 50 | 68.49 ± 17.29 ** | 87.69 ± 12.08 | |
| HIh-PE | 5 | 93.52 ± 10.47 | 101.18 ± 13.86 |
| 25 | 96.85 ± 9.89 | 101.76 ± 13.29 | |
| 50 | 99.98 ± 13.60 | 92.84 ± 14.54 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kozachok, S.; Kolodziejczyk-Czepas, J.; Marchyshyn, S.; Wojtanowski, K.K.; Zgórka, G.; Oleszek, W. Comparison of Phenolic Metabolites in Purified Extracts of Three Wild-Growing Herniaria L. Species and Their Antioxidant and Anti-Inflammatory Activities In Vitro. Molecules 2022, 27, 530. https://doi.org/10.3390/molecules27020530
Kozachok S, Kolodziejczyk-Czepas J, Marchyshyn S, Wojtanowski KK, Zgórka G, Oleszek W. Comparison of Phenolic Metabolites in Purified Extracts of Three Wild-Growing Herniaria L. Species and Their Antioxidant and Anti-Inflammatory Activities In Vitro. Molecules. 2022; 27(2):530. https://doi.org/10.3390/molecules27020530
Chicago/Turabian StyleKozachok, Solomiia, Joanna Kolodziejczyk-Czepas, Svitlana Marchyshyn, Krzysztof Kamil Wojtanowski, Grażyna Zgórka, and Wieslaw Oleszek. 2022. "Comparison of Phenolic Metabolites in Purified Extracts of Three Wild-Growing Herniaria L. Species and Their Antioxidant and Anti-Inflammatory Activities In Vitro" Molecules 27, no. 2: 530. https://doi.org/10.3390/molecules27020530
APA StyleKozachok, S., Kolodziejczyk-Czepas, J., Marchyshyn, S., Wojtanowski, K. K., Zgórka, G., & Oleszek, W. (2022). Comparison of Phenolic Metabolites in Purified Extracts of Three Wild-Growing Herniaria L. Species and Their Antioxidant and Anti-Inflammatory Activities In Vitro. Molecules, 27(2), 530. https://doi.org/10.3390/molecules27020530