
Evaluation of Total Phenolic and Flavonoid Contents, Antibacterial and Antibiofilm Activities of Hungarian Propolis Ethanolic Extract against Staphylococcus aureus

 , ,
, ,  ,
,
Abstract
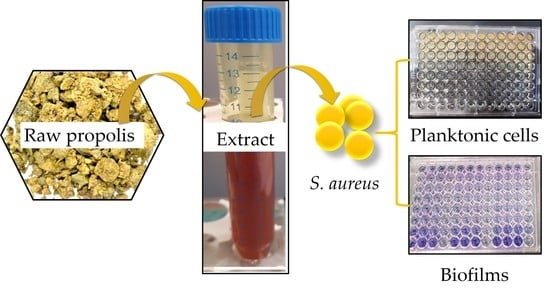
:
1. Introduction
2. Results and Discussion
2.1. Determination of the Total Phenolic and Flavonoid Contents
2.2. Antibacterial Susceptibility Test
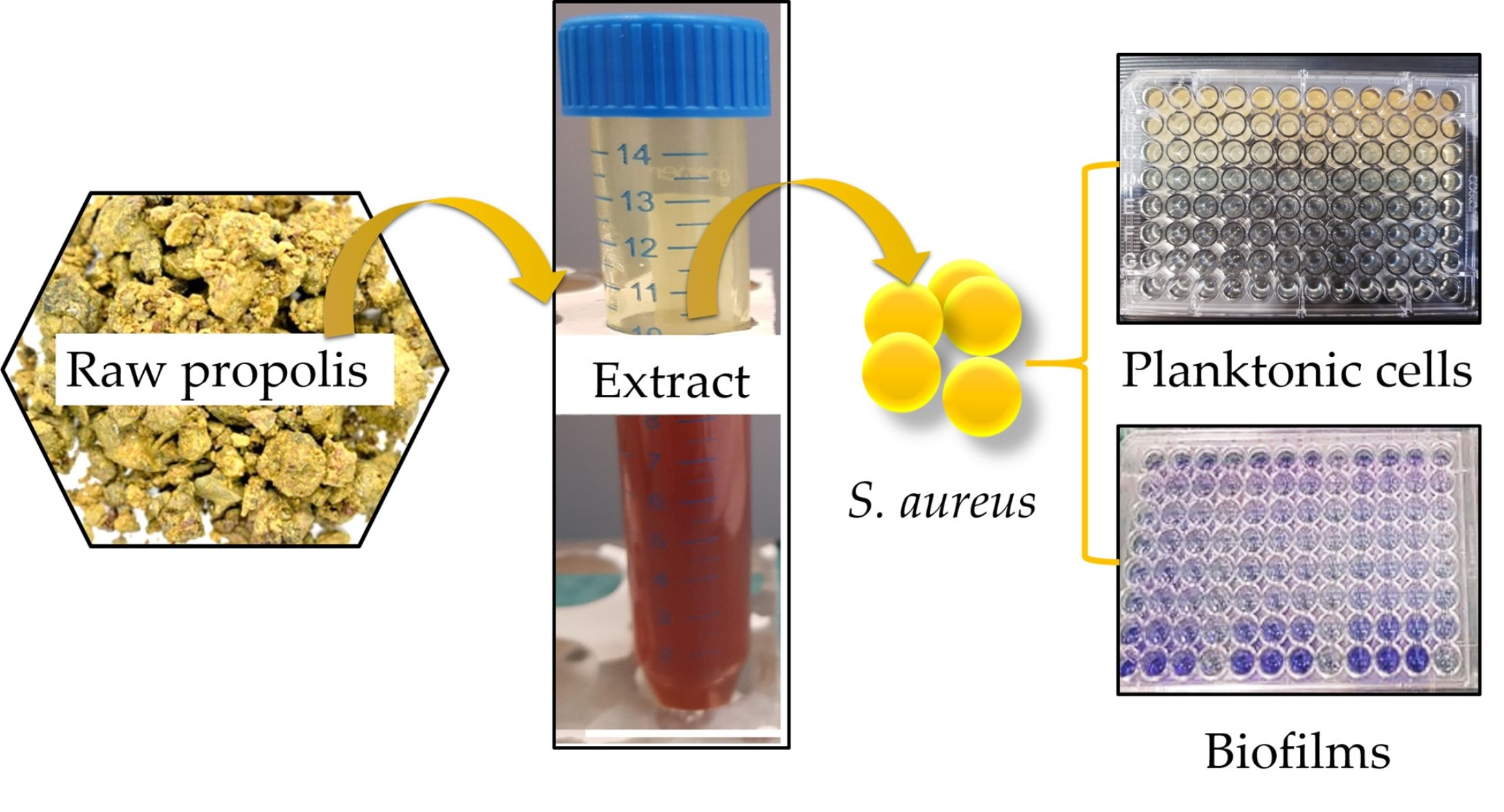
2.3. Killing Effect of EEP1 in Combination with Antibacterial Drugs
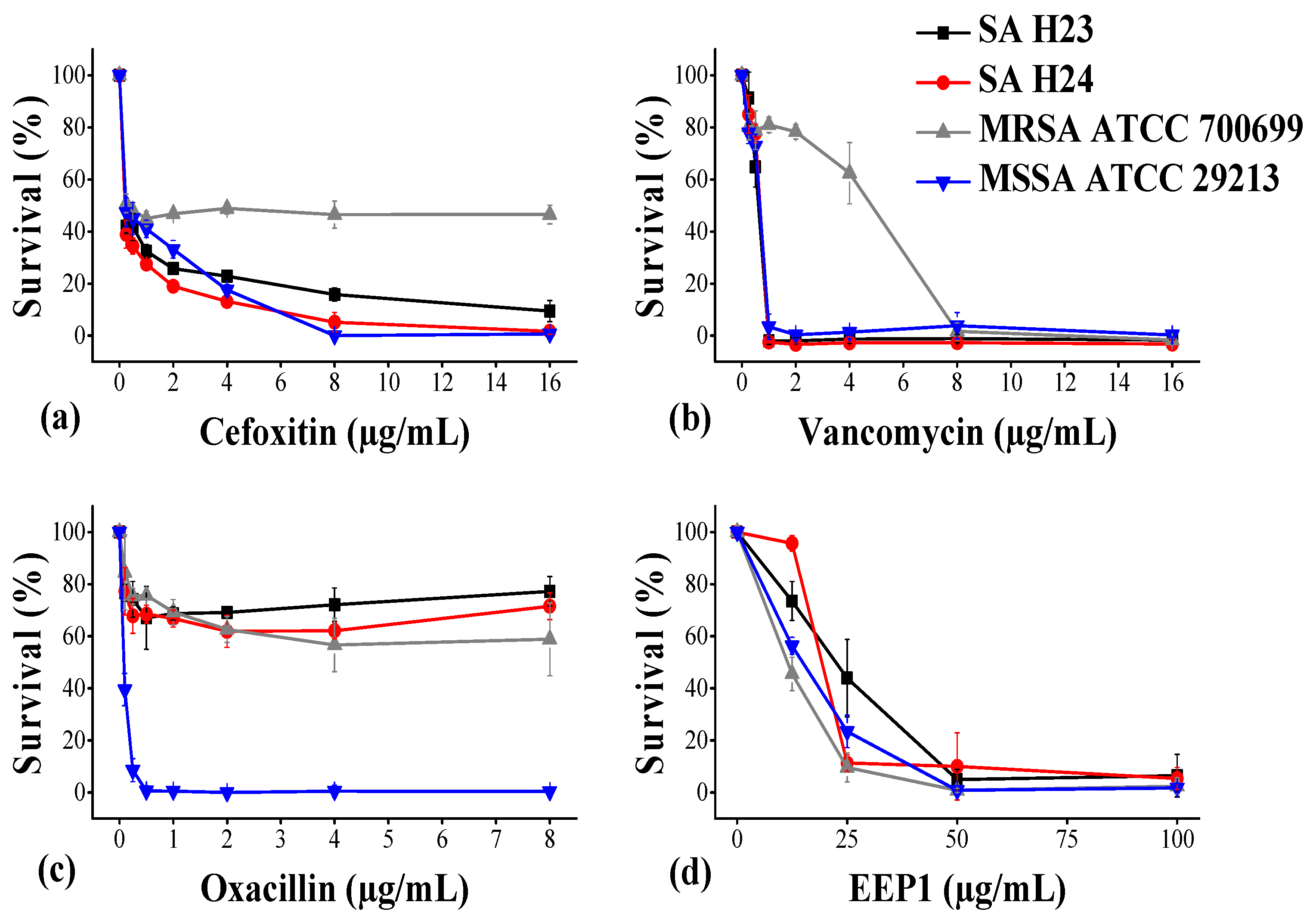
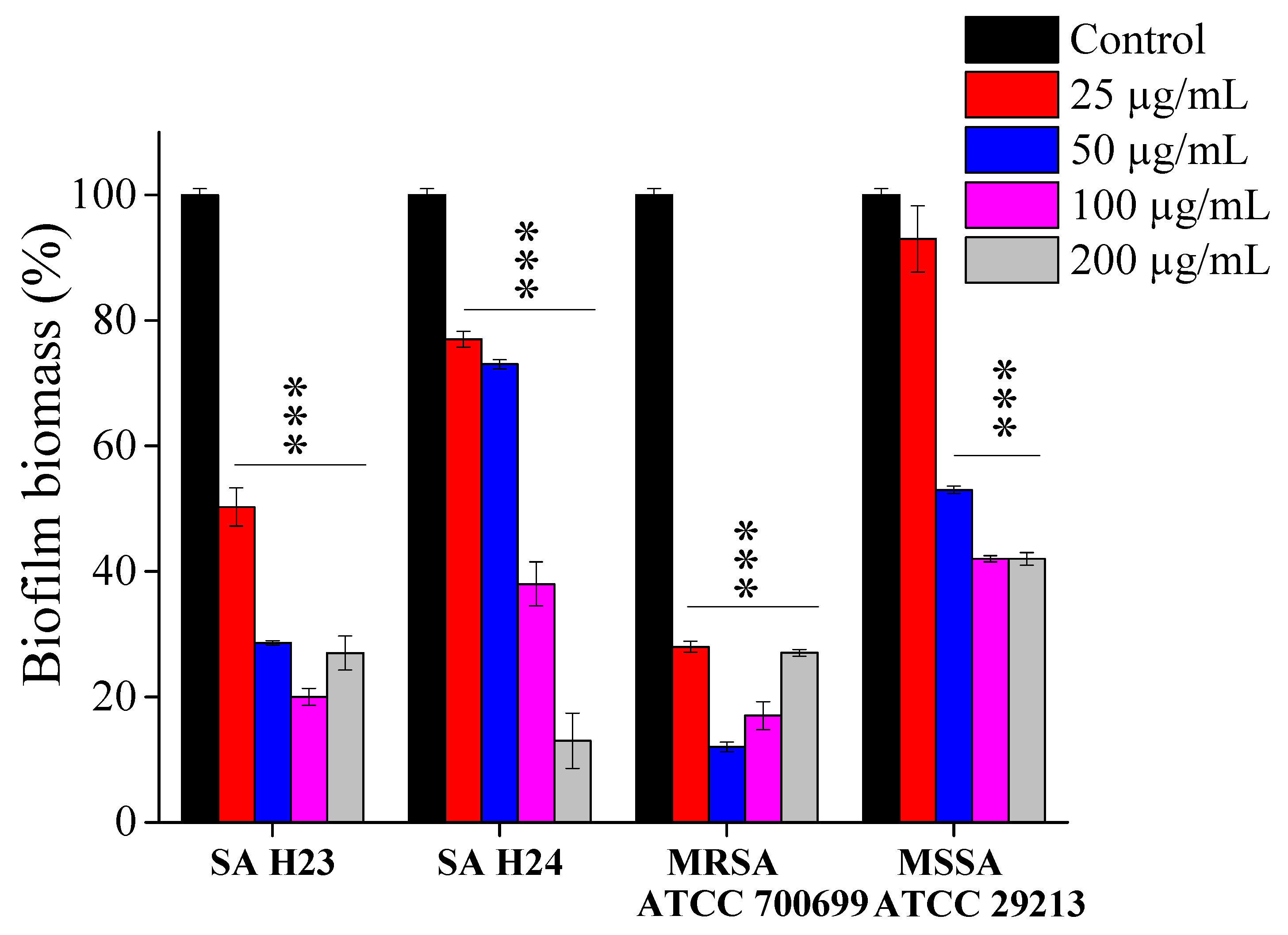
2.4. Effect of EEP1 on Biofilm Formation
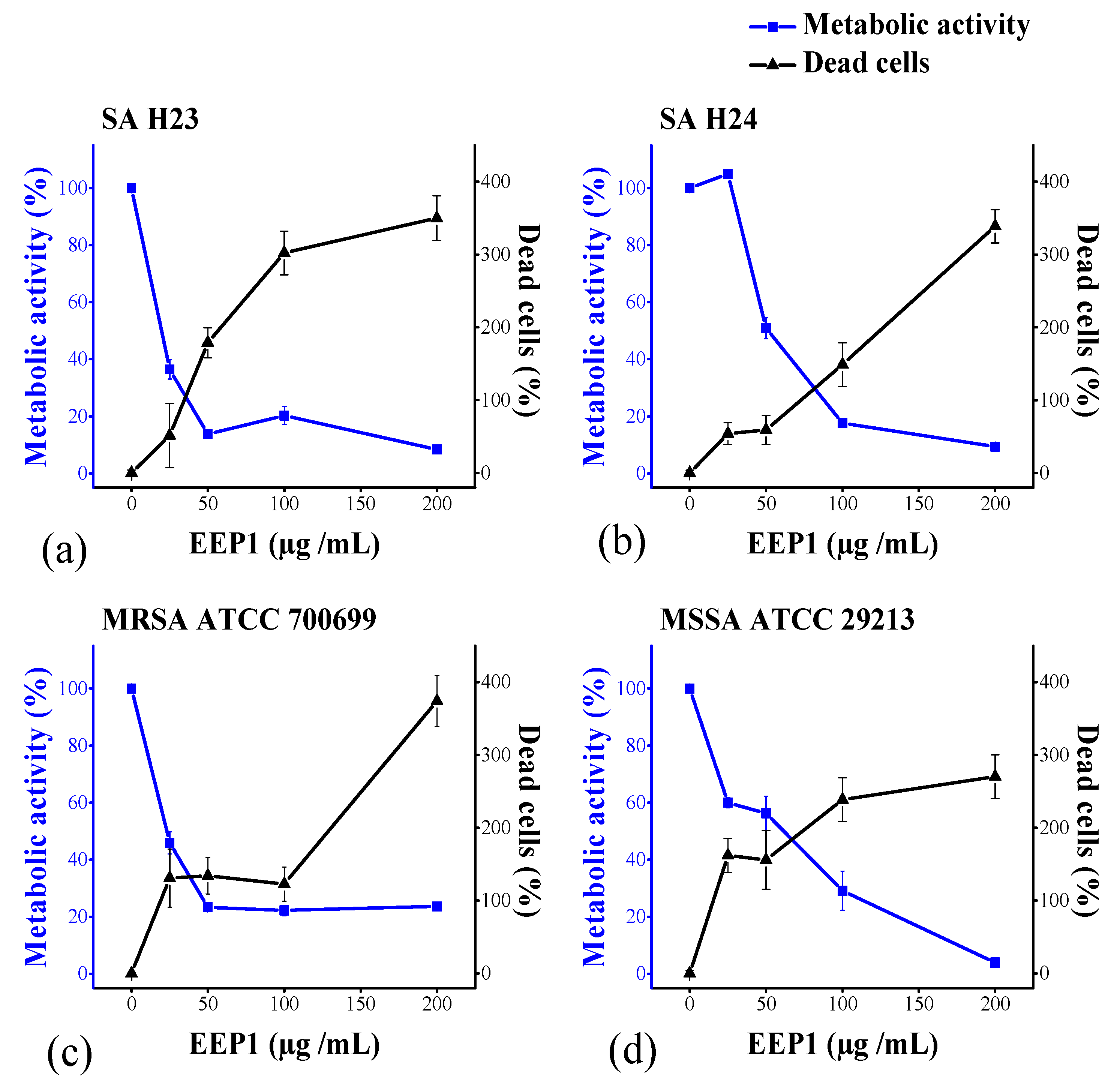
2.5. In Vitro Effect of EEP1 on the Eradication of Mature Biofilm
3. Materials and Methods
3.1. Microorganisms and Culture Conditions
3.2. Propolis Sources and Extraction
3.3. Determination of the Total Phenolic Content (TPC)
3.4. Determination of Total Flavonoids Content (TFC)
3.5. Antibacterial Susceptibility Test
3.5.1. MIC Value Determination
3.5.2. Chequerboard Broth Microdilution Method
3.6. Biofilm Formation and Quantification Assay
3.7. Biofilm Eradication Assay
3.8. Statistical Analyses
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Archer, N.; Mazaitis, M.J.; Costerton, J.W.; Leid, J.G.; Powers, M.E.; Shirtliff, M.E. Staphylococcus aureus biofilms: Properties, regulation, and roles in human disease. Virulence 2011, 2, 445–459. [Google Scholar] [CrossRef] [Green Version]
- Krismer, B.; Weidenmaier, C.; Zipperer, A.; Peschel, A. The commensal lifestyle of Staphylococcus aureus and its interactions with the nasal microbiota. Nat. Rev. Microbiol. 2017, 15, 675–687. [Google Scholar] [CrossRef] [PubMed]
- Otto, M. Staphylococcuscolonization of the skin and antimicrobial peptides. Expert Rev. Dermatol. 2010, 5, 183–195. [Google Scholar] [CrossRef] [Green Version]
- Thammavongsa, V.; Kim, H.K.; Missiakas, D.; Schneewind, O. Staphylococcal manipulation of host immune responses. Nat. Rev. Microbiol. 2015, 13, 529–543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foster, T.J. Antibiotic resistance in Staphylococcus aureus. Current status and future prospects. FEMS Microbiol. Rev. 2017, 41, 430–449. [Google Scholar] [CrossRef]
- Lai, C.-C.; Chen, C.-C.; Chuang, Y.-C.; Tang, H.-J. Combination of cephalosporins with vancomycin or teicoplanin enhances antibacterial effect of glycopeptides against heterogeneous vancomycin-intermediate Staphylococcus aureus (hVISA) and VISA. Sci. Rep. 2017, 7, 41758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boles, B.R.; Thoendel, M.; Roth, A.J.; Horswill, A.R. Identification of Genes Involved in Polysaccharide-Independent Staphylococcus aureus Biofilm Formation. PLoS ONE 2010, 5, e10146. [Google Scholar] [CrossRef] [Green Version]
- Kostakioti, M.; Hadjifrangiskou, M.; Hultgren, S.J. Bacterial Biofilms: Development, Dispersal, and Therapeutic Strategies in the Dawn of the Postantibiotic Era. Cold Spring Harb. Perspect. Med. 2013, 3, a010306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, D.; Misba, L.; Khan, A.U. Antibiotics versus biofilm: An emerging battleground in microbial communities. Antimicrob. Resist. Infect. Control 2019, 8, 76. [Google Scholar] [CrossRef]
- Bryan, J.; Redden, P.; Traba, C. The mechanism of action of Russian propolis ethanol extracts against two antibiotic-resistant biofilm-forming bacteria. Lett. Appl. Microbiol. 2015, 62, 192–198. [Google Scholar] [CrossRef]
- Brown, R. Hive Products: Pollen, Propolis and Royal Jelly. Bee World 1989, 70, 109–117. [Google Scholar] [CrossRef]
- Devequi-Nunes, D.; Machado, B.A.S.; de Abreu Barreto, G.; Rebouças Silva, J.; Da Silva, D.F.; Da Rocha, J.L.C.; Brandão, H.N.; Borges, V.M.; Umsza-Guez, M.A. Chemical characterization and biological activity of six different extracts of propolis through conventional methods and supercritical extraction. PLoS ONE 2018, 13, e0207676. [Google Scholar] [CrossRef] [PubMed]
- Przybyłek, I.; Karpiński, T.M. Antibacterial Properties of Propolis. Molecules 2019, 24, 2047. [Google Scholar] [CrossRef] [Green Version]
- Papp, Z.; Bouchelaghem, S.; Szekeres, A.; Meszéna, R.; Gyöngyi, Z.; Papp, G. The Scent of Antifungal Propolis. Sensors 2021, 21, 2334. [Google Scholar] [CrossRef]
- Campos, J.F.; Santos, U.P.D.; Rocha, P.D.S.D.; Damião, M.J.; Balestieri, J.B.P.; Cardoso, C.A.L.; Paredes-Gamero, E.J.; Estevinho, L.M.; de Picoli Souza, K.; dos Santos, E.L. Antimicrobial, Antioxidant, Anti-Inflammatory, and Cytotoxic Activities of Propolis from the Stingless Bee Tetragonisca fiebrigi (Jataí). Evid.-Based Complementary Altern. Med. 2015, 2015, 296186. [Google Scholar] [CrossRef] [Green Version]
- Kwiecińska-Piróg, J.; Skowron, K.; Śniegowska, A.; Przekwas, J.; Balcerek, M.; Załuski, D.; Gospodarek-Komkowska, E. The impact of ethanol extract of propolis on biofilm forming by Proteus Mirabilis strains isolated from chronic wounds infections. Nat. Prod. Res. 2018, 33, 3293–3297. [Google Scholar] [CrossRef] [PubMed]
- Pobiega, K.; Kraśniewska, K.; Przybył, J.L.; Bączek, K.; Żubernik, J.; Witrowa-Rajchert, D.; Gniewosz, M. Growth Biocontrol of Foodborne Pathogens and Spoilage Microorganisms of Food by Polish Propolis Extracts. Molecules 2019, 24, 2965. [Google Scholar] [CrossRef] [Green Version]
- Pobiega, K.; Igielska, M.; Włodarczyk, P.; Gniewosz, M. The use of pullulan coatings with propolis extract to extend the shelf life of blueberry (Vaccinium corymbosum) fruit. Int. J. Food Sci. Technol. 2021, 56, 1013–1020. [Google Scholar] [CrossRef]
- Al-Ani, I.; Zimmermann, S.; Reichling, J.; Wink, M. Antimicrobial Activities of European Propolis Collected from Various Geographic Origins Alone and in Combination with Antibiotics. Medicines 2018, 5, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tavafi, H.; Sadrzadeh-Afshar, M.S.; Niroomand, S. In vitro effectiveness of antimicrobial properties of propolis and chlorhexidine on oral pathogens: A comparative study. Biosis Biological Syst. 2020, 1, 116–125. [Google Scholar] [CrossRef]
- López, B.G.-C.; Sawaya, A.C.H.F. A Review of the Plant Origins, Composition and Biological Activity of Red Propolis. In Natural Products: Structure, Bioactivity and Applications; Nova Science Publishers, Inc.: New York, NY, USA, 2012; pp. 83–96. Available online: https://novapublishers.com/wp-content/uploads/2019/10/978-1-62081-728-5_ch4.pdf (accessed on 24 February 2021).
- Bouchelaghem, S. Propolis characterization and antimicrobial activities against Staphylococcus aureus and Candida albicans: A review. Saudi J. Biol. Sci. 2021, in press. [CrossRef]
- Molnár, S.; Mikuska, K.; Patonay, K.; Sisa, K.; Daood, H.G.; Némedi, E.; Kiss, A. Comparative studies on polyphenolic profile and antimicrobial activity of propolis samples selected from distinctive geographical areas of Hungary. Food Sci. Technol. Int. 2017, 23, 349–357. [Google Scholar] [CrossRef] [PubMed]
- Machado, C.S.; Mokochinski, J.B.; de Lira, T.O.; Oliveira, F.D.C.E.D.; Cardoso, M.V.; Ferreira, R.G.; Sawaya, A.; Ferreira, A.G.; Pessoa, C.; Cuesta-Rubio, O.; et al. Comparative Study of Chemical Composition and Biological Activity of Yellow, Green, Brown, and Red Brazilian Propolis. Evid.-Based Complement. Altern. Med. 2016, 2016, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zarate, M.S.H.; Juárez, M.D.R.A.; García, A.C.; López, C.O.; Chávez, A.J.G.; Garfias, J.J.N.S.; Ramos, F.A. Flavonoids, phenolic content, and antioxidant activity of propolis from various areas of Guanajuato, Mexico. Food Sci. Technol. 2018, 38, 210–215. [Google Scholar] [CrossRef] [Green Version]
- Gatea, F.; Teodor, E.D.; Radu, G.L.; Gille, E. Total polyphenols, flavonoids and antioxidant activity of Romanian propolis. An. Stiint. Univ. Al. I. Cuza Iasi Biol. Veg. Ser. Noua Sect. II A 2016, 62, 92–93. [Google Scholar]
- Nina, N.; Quispe, C.; Jiménez-Aspee, F.; Theoduloz, C.; Giménez, A.; Schmeda-Hirschmann, G. Chemical profiling and antioxidant activity of Bolivian propolis. J. Sci. Food Agric. 2016, 96, 2142–2153. [Google Scholar] [CrossRef]
- Asem, N.; Gapar, N.A.A.; Hapit, N.H.A.; Omar, E.A. Correlation between total phenolic and flavonoid contents with antioxidant activity of Malaysian stingless bee propolis extract. J. Apic. Res. 2020, 59, 437–442. [Google Scholar] [CrossRef]
- Gezgin, Y.; Kazan, A.; Ulucan, F.; Yesil-Celiktas, O. Antimicrobial activity of propolis and gentamycin against methicillin-resistant Staphylococcus aureus in a 3D thermo-sensitive hydrogel. Ind. Crop. Prod. 2019, 139, 111588. [Google Scholar] [CrossRef]
- Ozdal, T.; Ceylan, F.D.; Eroglu, N.; Kaplan, M.; Olgun, E.O.; Capanoglu, E. Investigation of antioxidant capacity, bioaccessibility and LC-MS/MS phenolic profile of Turkish propolis. Food Res. Int. 2019, 122, 528–536. [Google Scholar] [CrossRef]
- Mohtar, L.G.; Messina, G.A.; Bertolino, F.A.; Pereira, S.V.; Raba, J.; Nazareno, M.A. Comparative study of different methodologies for the determination the antioxidant activity of Venezuelan propolis. Microchem. J. 2020, 158, 105244. [Google Scholar] [CrossRef]
- Torres, A.; Sandjo, L.; Friedemann, M.; Tomazzoli, M.; Maraschin, M.; Mello, C.; Santos, A. Chemical characterization, antioxidant and antimicrobial activity of propolis obtained from Melipona quadrifasciata and Tetragonisca angustula stingless bees. Braz. J. Med Biol. Res. 2018, 51, e7118. [Google Scholar] [CrossRef] [PubMed]
- Narimane, S.; Demircan, E.; Salah, A.; Ozcelik, B.; Salah, R. Correlation between antioxidant activity and phenolic acids profile and content of Algerian propolis: Influence of solvent. Pak. J. Pharm. Sci. 2017, 30, 1417–1423. [Google Scholar] [PubMed]
- Pobiega, K.; Kraśniewska, K.; Derewiaka, D.; Gniewosz, M. Comparison of the antimicrobial activity of propolis extracts obtained by means of various extraction methods. J. Food Sci. Technol. 2019, 56, 5386–5395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wieczynska, A.; Wezgowiec, J.; Wieckiewicz, W.; Czarny, A.; Kulbacka, J.; Nowakowska, D.; Gancarz, R.; Wilk, K.A. Antimicrobial activity, cytotoxicity and total phenolic content of different extracts of propolis from the west Pomeranian region in Poland. Acta Pol. Pharm. 2017, 74, 715–722. [Google Scholar]
- Tlak, G.I.; Iva, P.; Mirza, B.; Ivan, K.; Siniša, S.; Toni, V.; Josipa, V. Components responsible for antimicrobial activity of propolis from continental and Mediterranean regions in Croatian. Czech J. Food Sci. 2017, 35, 376–385. [Google Scholar] [CrossRef] [Green Version]
- Daraghmeh, J.; Imtara, H. In Vitro Evaluation of Palestinian Propolis as a Natural Product with Antioxidant Properties and Antimicrobial Activity against Multidrug-Resistant Clinical Isolates. J. Food Qual. 2020, 2020, 8861395. [Google Scholar] [CrossRef]
- Ozdal, T.; Sari-Kaplan, G.; Altundag, E.M.; Boyacioglu, D.; Capanoglu, E. Evaluation of Turkish propolis for its chemical composition, antioxidant capacity, anti-proliferative effect on several human breast cancer cell lines and proliferative effect on fibroblasts and mouse mesenchymal stem cell line. J. Apic. Res. 2018, 57, 627–638. [Google Scholar] [CrossRef]
- Ristivojevic, P.; Trifkovic, J.; Andrić, F.; Milojković-Opsenica, D. Poplar-type Propolis: Chemical Composition, Botanical Origin and Biological Activity. Nat. Prod. Commun. 2015, 10, 1869–1876. [Google Scholar] [CrossRef] [Green Version]
- Chovanová, R.; Mikulášová, M.; Vaverková, Š. Modulation of mecA Gene Expression by Essential Oil from Salvia sclarea and Synergism with Oxacillin in Methicillin Resistant Staphylococcus epidermidis Carrying Different Types of Staphylococcal Chromosomal Cassettemec. Int. J. Microbiol. 2016, 2016, 6475837. [Google Scholar] [CrossRef] [Green Version]
- Howden, B.P.; Davies, J.K.; Johnson, P.D.R.; Stinear, T.P.; Grayson, M.L. Reduced Vancomycin Susceptibility in Staphylococcus aureus, Including Vancomycin-Intermediate and Heterogeneous Vancomycin-Intermediate Strains: Resistance Mechanisms, Laboratory Detection, and Clinical Implications. Clin. Microbiol. Rev. 2010, 23, 99–139. [Google Scholar] [CrossRef] [Green Version]
- Bueno-Silva, B.; Marsola, A.; Ikegaki, M.; Alencar, S.M.; Rosalen, P.L. The effect of seasons on Brazilian red propolis and its botanical source: Chemical composition and antibacterial activity. Nat. Prod. Res. 2016, 31, 1318–1324. [Google Scholar] [CrossRef] [PubMed]
- Regueira, M.; Tintino, S.R.; da Silva, A.R.P.; Costa, M.D.S.; Boligon, A.A.; Matias, E.F.; Balbino, V.D.Q.; Menezes, I.R.; Coutinho, H. Seasonal variation of Brazilian red propolis: Antibacterial activity, synergistic effect and phytochemical screening. Food Chem. Toxicol. 2017, 107, 572–580. [Google Scholar] [CrossRef] [PubMed]
- Nazeri, R.; Ghaiour, M.; Abbasi, S. Evaluation of Antibacterial Effect of Propolis and its Application in Mouthwash Production. Front. Dent. 2019, 16, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Al-Waili, N.; Al-Ghamdi, A.; Ansari, M.J.; Al-Attal, Y.; Salom, K. Synergistic Effects of Honey and Propolis toward Drug Multi-Resistant Staphylococcus aureus, Escherichia Coli and Candida albicans Isolates in Single and Polymicrobial Cultures. Int. J. Med Sci. 2012, 9, 793–800. [Google Scholar] [CrossRef] [Green Version]
- Vică, M.L.; Glevitzky, M.; Tit, D.M.; Behl, T.; Heghedűş-Mîndru, R.C.; Zaha, D.C.; Ursu, F.; Popa, M.; Glevitzky, I.; Bungău, S. The antimicrobial activity of honey and propolis extracts from the central region of Romania. Food Biosci. 2021, 41, 101014. [Google Scholar] [CrossRef]
- Surek, M.; Fachi, M.M.; Cobre, A.D.F.; de Oliveira, F.F.; Pontarolo, R.; Crisma, A.R.; de Souza, W.M.; Felipe, K.B. Chemical composition, cytotoxicity, and antibacterial activity of propolis from Africanized honeybees and three different Meliponini species. J. Ethnopharmacol. 2021, 269, 113662. [Google Scholar] [CrossRef]
- Wang, F.; Liu, H.; Li, J.; Zhang, W.; Jiang, B.; Xuan, H. Australian propolis ethanol extract exerts antibacterial activity against methicillin-resistant Staphylococcus aureus by mechanisms of disrupting cell structure, reversing resistance, and resisting biofilm. Braz. J. Microbiol. 2021, 52, 1651–1664. [Google Scholar] [CrossRef]
- Kashi, T.S.J.; Kermanshahi, R.K.; Erfan, M.; Dastjerdi, E.V.; Rezaei, Y.; Tabatabaei, F.S. Evaluating the In-vitro Antibacterial Effect of Iranian Propolis on Oral Microorganisms. Iran J. Pharm. Res. 2011, 10, 363–368. [Google Scholar]
- Lu, L.-C.; Chen, Y.-W.; Chou, C.-C. Antibacterial activity of propolis against Staphylococcus aureus. Int. J. Food Microbiol. 2005, 102, 213–220. [Google Scholar] [CrossRef]
- Popova, M.; Dimitrova, R.; Al-Lawati, H.T.; Tsvetkova, I.; Najdenski, H.; Bankova, V. Omani propolis: Chemical profiling, antibacterial activity and new propolis plant sources. Chem. Central J. 2013, 7, 158. [Google Scholar] [CrossRef] [Green Version]
- Uzel, A.; Sorkun, K.; Önçağ, Ö.; Çoğulu, D.; Gençay, Ö.; Sali˙h, B. Chemical compositions and antimicrobial activities of four different Anatolian propolis samples. Microbiol. Res. 2005, 160, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Miklasińska-Majdanik, M.; Kępa, M.; Wojtyczka, R.D.; Idzik, D.; Wąsik, T.J. Phenolic Compounds Diminish Antibiotic Resistance of Staphylococcus aureus Clinical Strains. Int. J. Environ. Res. Public Heal. 2018, 15, 2321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wojtyczka, R.D.; Dziedzic, A.; Idzik, D.; Kępa, M.; Kubina, R.; Kabała-Dzik, A.; Smoleń-Dzirba, J.; Stojko, J.; Sajewicz, M.; Wąsik, T.J. Susceptibility of Staphylococcus aureus Clinical Isolates to Propolis Extract Alone or in Combination with Antimicrobial Drugs. Molecules 2013, 18, 9623–9640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- LaPlante, K.L.; Rybak, M.J. Impact of High-Inoculum Staphylococcus aureus on the Activities of Nafcillin, Vancomycin, Linezolid, and Daptomycin, Alone and in Combination with Gentamicin, in an In Vitro Pharmacodynamic Model. Antimicrob. Agents Chemother. 2004, 48, 4665–4672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mizunaga, S.; Kamiyama, T.; Fukuda, Y.; Takahata, M.; Mitsuyama, J. Influence of inoculum size of Staphylococcus aureus and Pseudomonas aeruginosa on in vitro activities and in vivo efficacy of fluoroquinolones and carbapenems. J. Antimicrob. Chemother. 2005, 56, 91–96. [Google Scholar] [CrossRef] [Green Version]
- Grecka, K.; Kuś, P.M.; Okińczyc, P.; Worobo, R.W.; Walkusz, J.; Szweda, P. The Anti-Staphylococcal Potential of Ethanolic Polish Propolis Extracts. Molecules 2019, 24, 1732. [Google Scholar] [CrossRef] [Green Version]
- Krol, W.; Scheller, S.; Shani, J.; Pietsz, G.; Czuba, Z. Synergistic effect of ethanolic extract of propolis and antibiotics on the growth of Staphylococcus aureus. Arzneimittelforschung 1993, 43, 607–609. [Google Scholar]
- Júnior, A.F.; Balestrin, E.C.; Betoni, J.E.C.; Orsi, R.D.O.; Cunha, M.D.L.R.D.S.D.; Montelli, A.C. Propolis: Anti-Staphylococcus aureus activity and synergism with antimicrobial drugs. Mem. Inst. Oswaldo Cruz 2005, 100, 563–566. [Google Scholar] [CrossRef] [Green Version]
- Reichmann, N.T.; Pinho, M.G. Role of SCCmec type in resistance to the synergistic activity of oxacillin and cefoxitin in MRSA. Sci. Rep. 2017, 7, 6154. [Google Scholar] [CrossRef] [Green Version]
- Naorem, R.S.; Urban, P.; Goswami, G.; Fekete, C. Characterization of methicillin-resistant Staphylococcus aureus through genomics approach. 3 Biotech 2020, 10, 401. [Google Scholar] [CrossRef]
- Yoshii, Y.; Okuda, K.-I.; Yamada, S.; Nagakura, M.; Sugimoto, S.; Nagano, T.; Okabe, T.; Kojima, H.; Iwamoto, T.; Kuwano, K.; et al. Norgestimate inhibits staphylococcal biofilm formation and resensitizes methicillin-resistant Staphylococcus aureus to β-lactam antibiotics. NPJ Biofilms Microbiomes 2017, 3, 1–9. [Google Scholar] [CrossRef]
- Ghasemian, A.; Peerayeh, S.N.; Bakhshi, B.; Mirzaee, M. Comparison of Biofilm Formation between Methicillin-Resistant and Methicillin-Susceptible Isolates of Staphylococcus aureus. Iran. Biomed. J. 2016, 20, 175–181. [Google Scholar] [CrossRef]
- Miao, J.; Lin, S.; Soteyome, T.; Peters, B.M.; Li, Y.; Chen, H.; Su, J.; Li, L.; Li, B.; Xu, Z.; et al. Biofilm Formation of Staphylococcus aureus under Food Heat Processing Conditions: First Report on CML Production within Biofilm. Sci. Rep. 2019, 9, 1312. [Google Scholar] [CrossRef] [PubMed]
- Piechota, M.; Kot, B.; Frankowska-Maciejewska, A.; Grużewska, A.; Woźniak-Kosek, A. Biofilm Formation by Methicillin-Resistant and Methicillin-Sensitive Staphylococcus aureus Strains from Hospitalized Patients in Poland. BioMed Res. Int. 2018, 2018, 4657396. [Google Scholar] [CrossRef] [Green Version]
- Wojtyczka, R.D.; Kępa, M.; Idzik, D.; Kubina, R.; Kabała-Dzik, A.; Dziedzic, A.; Wąsik, T.J. In VitroAntimicrobial Activity of Ethanolic Extract of Polish Propolis against Biofilm Forming Staphylococcus epidermidis Strains. Evid.-Based Complement. Altern. Med. 2013, 2013, 590703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Marco, S.; Piccioni, M.; Pagiotti, R.; Pietrella, D. Antibiofilm and Antioxidant Activity of Propolis and Bud Poplar Resins versus Pseudomonas aeruginosa. Evid.-Based Complement. Altern. Med. 2017, 2017, 5163575. [Google Scholar] [CrossRef] [Green Version]
- Alfarrayeh, I.; Fekete, C.; Gazdag, Z.; Papp, G. Propolis ethanolic extract has double-face in vitro effect on the planktonic growth and biofilm formation of some commercial probiotics. Saudi J. Biol. Sci. 2021, 28, 1033–1039. [Google Scholar] [CrossRef] [PubMed]
- Budzyńska, A.; Wieckowska-Szakiel, M.; Sadowska, B.; Kalemba, D.; Rózalska, B. Antibiofilm activity of selected plant essential oils and their major components. Pol. J. Microbiol. 2011, 60, 35–41. [Google Scholar] [CrossRef]
- de Azevedo, M.N.; Marques, N.T.; Fonseca, M.F.L.; Schuch, L.F.; de Arruda, J.A.A.; Santos, V.R.; Mesquita, R.A.; Moreno, A. Disinfectant effects of Brazilian green propolis alcohol solutions on the Staphylococcus aureus biofilm of maxillofacial prosthesis polymers. J. Prosthet. Dent. 2021, in press. [CrossRef]
- Daikh, A.; Segueni, N.; Dogan, N.M.; Arslan, S.; Mutlu, D.; Kivrak, I.; Akkal, S.; Rhouati, S. Comparative study of antibiofilm, cytotoxic activity and chemical composition of Algerian propolis. J. Apic. Res. 2020, 59, 160–169. [Google Scholar] [CrossRef]
- Hazem, A.; Popescu, C.V.; Crișan, I.; Popa, M.; Chifiriuc, M.C.; Pircalabioru, S.; Lupuliasa, D. Antibacterial efficiency of five propolis extracts on planktonic and adherent microbial strains. Farmacia 2017, 65, 813–818. [Google Scholar]
- Ambi, A.; Bryan, J.; Borbon, K.; Centeno, D.; Liu, T.; Chen, T.P.; Cattabiani, T.; Traba, C. Are Russian propolis ethanol extracts the future for the prevention of medical and biomedical implant contaminations? Phytomedicine 2017, 30, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Grecka, K.; Xiong, Z.R.; Chen, H.; Pełka, K.; Worobo, R.W.; Szweda, P. Effect of Ethanol Extracts of Propolis (EEPs) against Staphylococcal Biofilm—Microscopic Studies. Pathogens 2020, 9, 646. [Google Scholar] [CrossRef] [PubMed]
- Dembogurski, D.S.D.O.; Trentin, D.S.; Boaretto, A.G.; Rigo, G.V.; Campos-Silva, R.; Tasca, T.; Macedo, A.J.; Carollo, C.A.; Silva, D.B. Brown propolis-metabolomic innovative approach to determine compounds capable of killing Staphylococcus aureus biofilm and Trichomonas vaginalis. Food Res. Int. 2018, 111, 661–673. [Google Scholar] [CrossRef] [PubMed]
- Alencar, S.; Oldoni, T.; Castro, M.; Cabral, I.; Costa-Neto, C.; Cury, J.; Rosalen, P.L.; Ikegaki, M. Chemical composition and biological activity of a new type of Brazilian propolis: Red propolis. J. Ethnopharmacol. 2007, 113, 278–283. [Google Scholar] [CrossRef] [PubMed]
- Moreira, L.F.; Dias, L.; Pereira, J.A.; Estevinho, M.L.M.F. Antioxidant properties, total phenols and pollen analysis of propolis samples from Portugal. Food Chem. Toxicol. 2008, 46, 3482–3485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dias, L.; Pereira, A.P.; Estevinho, M.L.M.F. Comparative study of different Portuguese samples of propolis: Pollinic, sensorial, physicochemical, microbiological characterization and antibacterial activity. Food Chem. Toxicol. 2012, 50, 4246–4253. [Google Scholar] [CrossRef] [PubMed]
- CLSI. Methods for Antimicrobial Susceptibility Testing of Anaerobic Bacteria: Approved Standard, 8th ed.; CLSI Document M11-A8; Clinical and Laboratory Standards Inst.: Wayne, PA, USA, 2012; ISBN1 1-56238-789-8 (Print). ISBN2 1-56238-789-8 (Electronic). [Google Scholar]
- Stepanović, S.; Vuković, D.; Hola, V.; DI Bonaventura, G.; Djukić, S.; Ćirković, I.; Ruzicka, F. Quantification of biofilm in microtiter plates: Overview of testing conditions and practical recommendations for assessment of biofilm production by staphylococci. APMIS 2007, 115, 891–899. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Propolis Samples | TPC (mg GAE/g DW) | TFC (mg CAE/g DW) |
|---|---|---|
| EEP1 | 71.1 ± 4.3 a | 273.2 ± 10.2 a |
| EEP2 | 55.8 ± 2.0 b | 172.8 ± 11.5 b |
| EEP3 | 47.9 ± 0.2 c | 164.1 ± 2.7 b,c |
| EEP4 | 44.0 ± 0.5 c | 142.5 ± 4.2 c,d |
| EEP5 | 34.6 ± 1.0 d | 147.3 ± 12.9 c,d |
| EEP6 | 10.4 ± 1.4 e | 33.8 ± 3.0 e |
| S. aureus Strains | Drugs | MIC Value (µg/mL) | FICI (Type of Interaction) | |
|---|---|---|---|---|
| Alone | In Combination | |||
| SA H23 | EEP1 | 200 | <200 | N |
| Oxacillin | <4 | <4 | ||
| EEP1 | 200 | 200 | N | |
| Cefoxitin | <8 | 0.13 | ||
| EEP1 | 200 | 3.13 | 0.03 (synergistic) | |
| Vancomycin | 2 | <200 | ||
| SA H24 | EEP1 | 200 | 200 | N |
| Oxacillin | <4 | 0.06 | ||
| EEP1 | 200 | 200 | N | |
| Cefoxitin | <8 | 0.13 | ||
| EEP1 | 200 | 3.13 | 0.03 (synergistic) | |
| Vancomycin | 2 | 0.03 | ||
| MSSA | EEP1 | 100 | 25 | 0.27 (synergistic) |
| Oxacillin | 4 | 0.06 | ||
| EEP1 | 100 | 3.13 | 0.05 (synergistic) | |
| Cefoxitin | 2 | 0.03 | ||
| EPP | 100 | 3.13 | 0.05 (synergistic) | |
| Vancomycin | 2 | 0.03 | ||
| MRSA | EEP1 | 400 | 400 | N |
| Oxacillin | <4 | 0.06 | ||
| EEP1 | 400 | 200 | N | |
| Cefoxitin | <8 | 0.13 | ||
| EPP1 | 400 | 6.25 | 0.05 (synergistic) | |
| Vancomycin | 4 | 0.13 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bouchelaghem, S.; Das, S.; Naorem, R.S.; Czuni, L.; Papp, G.; Kocsis, M. Evaluation of Total Phenolic and Flavonoid Contents, Antibacterial and Antibiofilm Activities of Hungarian Propolis Ethanolic Extract against Staphylococcus aureus. Molecules 2022, 27, 574. https://doi.org/10.3390/molecules27020574
Bouchelaghem S, Das S, Naorem RS, Czuni L, Papp G, Kocsis M. Evaluation of Total Phenolic and Flavonoid Contents, Antibacterial and Antibiofilm Activities of Hungarian Propolis Ethanolic Extract against Staphylococcus aureus. Molecules. 2022; 27(2):574. https://doi.org/10.3390/molecules27020574
Chicago/Turabian StyleBouchelaghem, Sarra, Sourav Das, Romen Singh Naorem, Lilla Czuni, Gábor Papp, and Marianna Kocsis. 2022. "Evaluation of Total Phenolic and Flavonoid Contents, Antibacterial and Antibiofilm Activities of Hungarian Propolis Ethanolic Extract against Staphylococcus aureus" Molecules 27, no. 2: 574. https://doi.org/10.3390/molecules27020574
APA StyleBouchelaghem, S., Das, S., Naorem, R. S., Czuni, L., Papp, G., & Kocsis, M. (2022). Evaluation of Total Phenolic and Flavonoid Contents, Antibacterial and Antibiofilm Activities of Hungarian Propolis Ethanolic Extract against Staphylococcus aureus. Molecules, 27(2), 574. https://doi.org/10.3390/molecules27020574