
The Role of Diacylglycerol Kinase in the Amelioration of Diabetic Nephropathy


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Diabetes and Diabetic Nephropathy
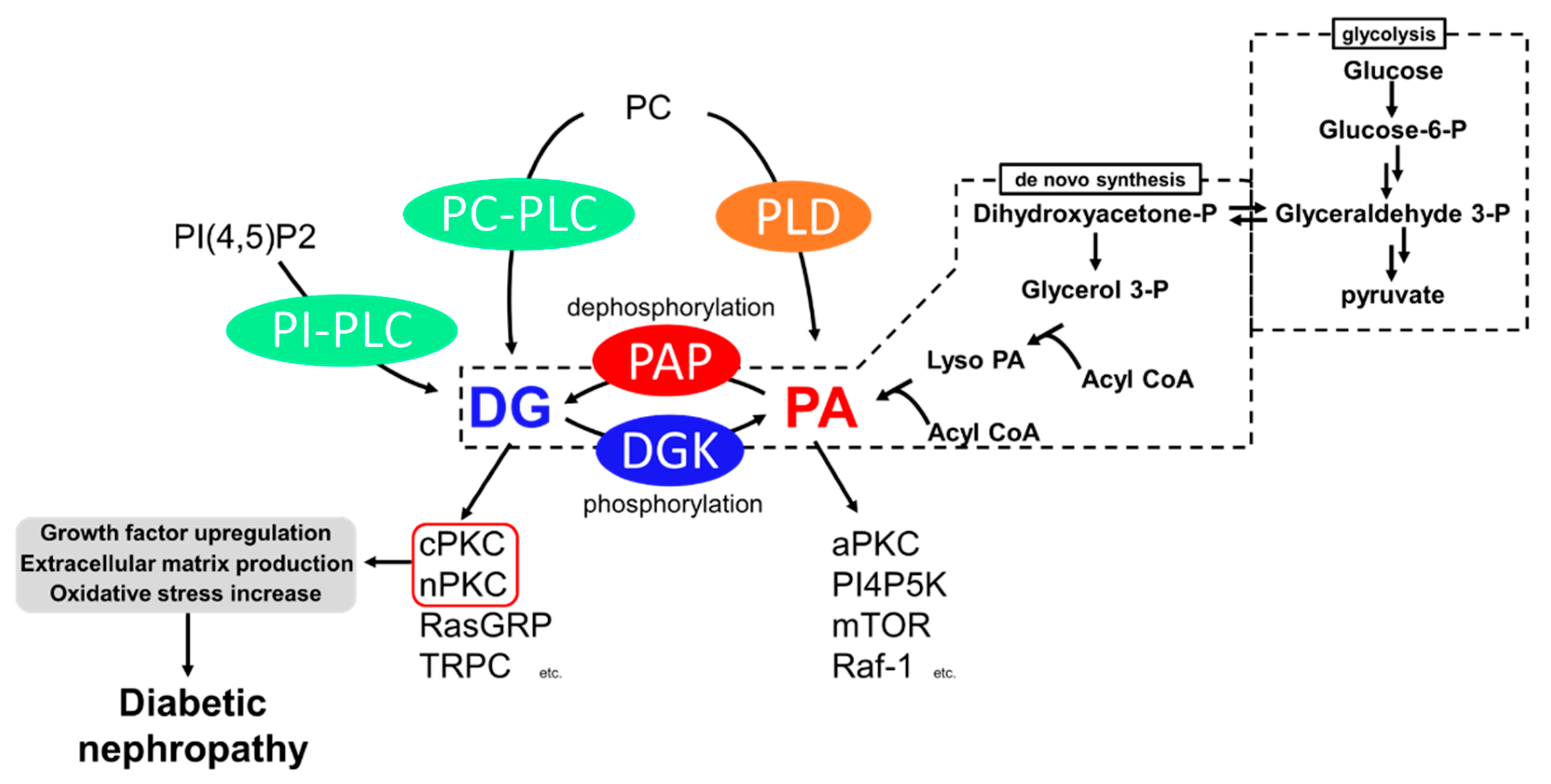
2. Hyperglycemia Impairs Lipid Signaling
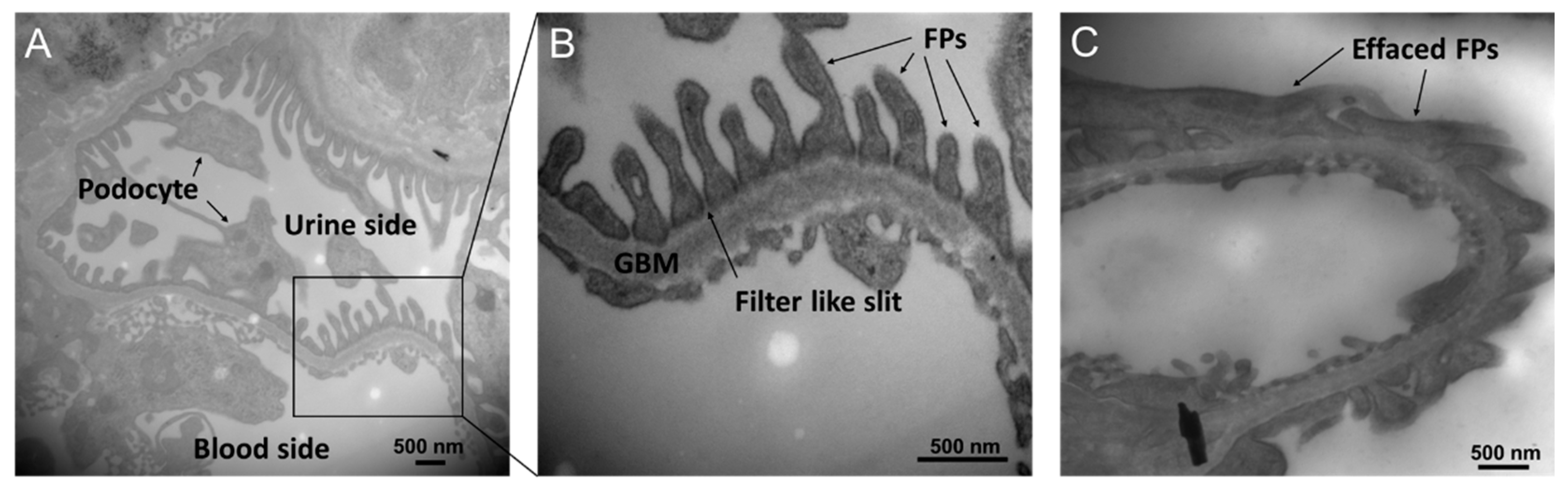
3. Diabetic Nephropathy and PKC
4. Diabetic Nephropathy and DGK
5. The Mechanisms of the Amelioration of DN by DGKα Activation
6. The Mechanisms of DGKα Activation
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sun, H.; Saeedi, P.; Karuranga, S.; Pinkepank, M.; Ogurtsova, K.; Duncan, B.B.; Stein, C.; Basit, A.; Chan, J.C.N.; Mbanya, J.C.; et al. IDF Diabetes Atlas: Global, Regional and Country-Level Diabetes Prevalence Estimates for 2021 and Projections for 2045. Diabetes Res. Clin. Pract. 2022, 183, 109119. [Google Scholar] [CrossRef] [PubMed]
- Sarwar, N.; Gao, P.; Kondapally Seshasai, S.R.; Gobin, R.; Kaptoge, S.; Di Angelantonio, E.; Ingelsson, E.; Lawlor, D.A.; Selvin, E.; Stampfer, M.; et al. Diabetes Mellitus, Fasting Blood Glucose Concentration, and Risk of Vascular Disease: A Collaborative Meta-Analysis of 102 Prospective Studies. Lancet 2010, 375, 2215–2222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carey, I.M.; Critchley, J.A.; Dewilde, S.; Harris, T.; Hosking, F.J.; Cook, D.G. Risk of Infection in Type 1 and Type 2 Diabetes Compared with the General Population: A Matched Cohort Study. Diabetes Care 2018, 41, 513–521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hovind, P.; Tarnow, L.; Rossing, P.; Jensen, B.R.; Graae, M.; Torp, I.; Binder, C.; Parving, H.H. Predictors for the Development of Microalbuminuria and Macroalbuminuria in Patients with Type 1 Diabetes: Inception Cohort Study. Br. Med. J. 2004, 328, 1105–1108. [Google Scholar] [CrossRef] [Green Version]
- Samsu, N. Diabetic Nephropathy: Challenges in Pathogenesis, Diagnosis, and Treatment. Biomed. Res. Int. 2021, 2021, 1497449. [Google Scholar] [CrossRef] [PubMed]
- Secrist, J.P.; Karnitz, L.; Abraham, R.T. T-Cell Antigen Receptor Ligation Induces Tyrosine Phosphorylation of Phospholipase C-Γ1. J. Biol. Chem. 1991, 266, 12135–12139. [Google Scholar] [CrossRef]
- Rhee, S.G.; Bae, Y.S. Regulation of Phosphoinositide-Specific Phospholipase C Isozymes. J. Biol. Chem. 1997, 272, 15045–15048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monick, M.M.; Carter, A.B.; Gudmundsson, G.; Mallampalli, R.; Powers, L.S.; Hunninghake, G.W. A Phosphatidylcholine-Specific Phospholipase C Regulates Activation of P42/44 Mitogen-Activated Protein Kinases in Lipopolysaccharide-Stimulated Human Alveolar Macrophages. J. Immunol. 1999, 162, 3005–3012. [Google Scholar]
- Carman, G.M.; Han, G.S. Phosphatidic Acid Phosphatase, a Key Enzyme in the Regulation of Lipid Synthesis. J. Biol. Chem. 2009, 284, 2593–2597. [Google Scholar] [CrossRef] [Green Version]
- Carman, G.M.; Han, G.-S. Roles of Phosphatidate Phosphatase Enzymes in Lipid Metabolism. Trends Biochem. Sci. 2006, 31, 694–699. [Google Scholar] [CrossRef] [Green Version]
- Exton, J.H. Phospholipase D: Enzymology, Mechanisms of Regulation, and Function. Physiol. Rev. 1997, 77, 303–320. [Google Scholar] [CrossRef] [PubMed]
- Craven, P.A.; Davidson, C.M.; DeRubertis, F.R. Increase in Diacylglycerol Mass in Isolated Glomeruli by Glucose from de Novo Synthesis of Glycerolipids. Diabetes 1990, 39, 667–674. [Google Scholar] [CrossRef] [PubMed]
- Thorens, B.; Mueckler, M. Glucose Transporters in the 21st Century. Am. J. Physiol. Endocrinol. Metab. 2010, 298, 141–145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, X.; Yang, F.; Brailoiu, E.; Jakubowski, H.; Dun, N.J.; Schafer, A.I.; Yang, X.; Durante, W.; Wang, H. Differential Regulation of Homocysteine Transport in Vascular Endothelial and Smooth Muscle Cells. Diabetes 1993, 42, 80–89. [Google Scholar] [CrossRef] [Green Version]
- Nishizuka, Y. Intracellular Signaling by Hydrolysis of Phospholipids and Activation of Protein Kinase, C. Science 1992, 258, 607–614. [Google Scholar] [CrossRef]
- Newton, A.C. Regulation of Protein Kinase, C. Curr. Opin. Cell Biol. 1997, 9, 161–167. [Google Scholar] [CrossRef]
- Ebinu, J.O.; Bottorff, D.A.; Chan, E.Y.W.; Stang, S.L.; Dunn, R.J.; Stone, J.C. RasGRP, a Ras Guanyl Nucleotide- Releasing Protein with Calcium- and Diacylglycerol-Binding Motifs. Science 1998, 280, 1082–1086. [Google Scholar] [CrossRef]
- Lucas, P.; Ukhanov, K.; Leinders-Zufall, T.; Zufall, F. A Diacylglycerol-Gated Cation Channel in Vomeronasal Neuron Dendrites Is Impaired in TRPC2 Mutant Mice: Mechanism of Pheromone Transduction. Neuron 2003, 40, 551–561. [Google Scholar] [CrossRef] [Green Version]
- Limatola, C.; Schaap, D.; Moolenaar, W.H.; van Blitterswijk, W.J. Phosphatidic Acid Activation of Protein Kinase C-Zeta Overexpressed in COS Cells: Comparison with Other Protein Kinase C Isotypes and Other Acidic Lipids. Biochem. J. 1994, 304, 1001–1008. [Google Scholar] [CrossRef]
- Jones, D.R.; Sanjuan, M.A.; Mérida, I. Type Iα Phosphatidylinositol 4-Phosphate 5-Kinase Is a Putative Target for Increased Intracellular Phosphatidic Acid. FEBS Lett. 2000, 476, 160–165. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, S.; Strum, J.C.; Sciorra, V.A.; Daniel, L.; Bell, R.M. Raf-1 Kinase Possesses Distinct Binding Domains for Phosphatidylserine and Phosphatidic Acid: Phosphatidic Acid Regulates the Translocation of Raf-1 in 12-O-Tetradecanoylphorbol-13-Acetate-Stimulated Madin-Darby Canine Kidney Cells. J. Biol. Chem. 1996, 271, 8472–8480. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Vilella-Bach, M.; Bachmann, R.; Flanigan, A.; Chen, J. Phosphatidic Acid-Mediated Mitogenic Activation of MTOR Signaling. Science 2001, 294, 1942–1945. [Google Scholar] [CrossRef] [PubMed]
- Newton, A.C. Protein Kinase C: Perfectly Balanced. Crit. Rev. Biochem. Mol. Biol. 2018, 53, 208–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parker, P.J.; Coussens, L.; Totty, N.; Rhee, L.; Young, S.; Chen, E.; Stabel, S.; Waterfield, M.D.; Ullrich, A. The Complete Primary Structure of Protein Kinase C—The Major Phorbol Ester Receptor. Science 1986, 233, 853–859. [Google Scholar] [CrossRef] [PubMed]
- Knopf, J.L.; Lee, M.H.; Sultzman, L.A.; Kriz, R.W.; Loomis, C.R.; Hewick, R.M.; Bell, R.M. Cloning and Expression of Multiple Protein Kinase C CDNAs. Cell 1986, 46, 491–502. [Google Scholar] [CrossRef]
- Ohno, S.; Akita, Y.; Konno, Y.; Imajoh, S.; Suzuki, K. A Novel Phorbol Ester Receptor/Protein Kinase, NPKC, Distantly Related to the Protein Kinase C Family. Cell 1988, 53, 731–741. [Google Scholar] [CrossRef]
- Ono, Y.; Fujii, T.; Ogita, K.; Kikkawa, U.; Igarashi, K.; Nishizuka, Y. The Structure, Expression, and Properties of Additional Members of the Protein Kinase C Family. J. Biol. Chem. 1988, 263, 6927–6932. [Google Scholar] [CrossRef]
- Nakanishi, H.; Exton, J.H. Purification and Characterization of the ζ Isoform of Protein Kinase C from Bovine Kidney. J. Biol. Chem. 1992, 267, 16347–16354. [Google Scholar] [CrossRef]
- Craven, P.A.; DeRubertis, F. Protein Kinase C Is Activated in Glomeruli from Streptozotocin Diabetic Rats. Possible Mediation by Glucose. J. Clin. Investig. 1989, 83, 1667–1675. [Google Scholar] [CrossRef]
- Inoguchi, T.; Battan, R.; Handler, E.; Sportsman, J.R.; Heath, W.; King, G.L. Preferential Elevation of Protein Kinase C Isoform Beta II and Diacylglycerol Levels in the Aorta and Heart of Diabetic Rats: Differential Reversibility to Glycemic Control by Islet Cell Transplantation. Proc. Natl. Acad. Sci. USA 1992, 89, 11059–11063. [Google Scholar] [CrossRef] [Green Version]
- Ayo, S.H.; Radnik, R.; Garoni, J.A.; Troyer, D.A.; Kreisberg, J.I. High Glucose Increases Diacylglycerol Mass and Activates Protein Kinase C in Mesangial Cell Cultures. Am. J. Physiol. Physiol. 1991, 261, F571–F577. [Google Scholar] [CrossRef] [PubMed]
- Koya, D.; King, G.L. Protein Kinase C Activation and the Development of Diabetic Complications. Diabetes 1998, 47, 859–866. [Google Scholar] [CrossRef] [PubMed]
- Menne, J.; Park, J.K.; Boehne, M.; Elger, M.; Lindschau, C.; Kirsch, T.; Meier, M.; Gueler, F.; Fiebeler, A.; Bahlmann, F.H.; et al. Diminished Loss of Proteoglycans and Lack of Albuminuria in Protein Kinase C-α-Deficient Diabetic Mice. Diabetes 2004, 53, 2101–2109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohshiro, Y.; Ma, R.C.; Yasuda, Y.; Hiraoka-Yamamoto, J.; Clermont, A.C.; Isshiki, K.; Yagi, K.; Arikawa, E.; Kern, T.S.; King, G.L. Reduction of Diabetes-Induced Oxidative Stress, Fibrotic Cytokine Expression, and Renal Dysfunction in Protein Kinase Cβ-Null Mice. Diabetes 2006, 55, 3112–3120. [Google Scholar] [CrossRef] [Green Version]
- Tuttle, K.R.; Bakris, G.L.; Toto, R.D.; McGill, J.B.; Hu, K.; Anderson, P.W. The Effect of Ruboxistaurin on Nephropathy in Type 2 Diabetes. Diabetes Care 2005, 28, 2686–2690. [Google Scholar] [CrossRef] [Green Version]
- Shiba, T.; Inoguchi, T.; Sportsman, J.R.; Heath, W.F.; Bursell, S.; King, G.L. Correlation of Diacylglycerol Level and Protein Kinase C Activity in Rat Retina to Retinal Circulation. Am. J. Physiol. 1993, 265, E783–E793. [Google Scholar] [CrossRef]
- Derubertis, F.R.; Craven, P.A. Activation of Protein Kinase C in Glomerular Cells in Diabetes: Mechanisms and Potential Links to the Pathogenesis of Diabetic Glomerulopathy. Diabetes 1994, 43, 1–8. [Google Scholar] [CrossRef]
- Babazono, T.; Kapor-Drezgic, J.; Dlugosz, J.A.; Whiteside, C. Altered Expression and Subcellular Localization of Diacylglycerol- Sensitive Protein Kinase C Isoforms in Diabetic Rat Glomerular Cells. Diabetes 1998, 47, 668–676. [Google Scholar] [CrossRef]
- Chiarelli, F.; Gaspari, S.; Marcovecchio, M.L. Role of Growth Factors in Diabetic Kidney Disease. Horm. Metab. Res. 2009, 41, 585–593. [Google Scholar] [CrossRef]
- Reeves, W.B.; Andreoli, T.E. Transforming Growth Factor β Contributes to Progressive Diabetic Nephropathy. Proc. Natl. Acad. Sci. USA 2000, 97, 7667–7669. [Google Scholar] [CrossRef] [Green Version]
- Arora, M.K.; Singh, U.K. Molecular Mechanisms in the Pathogenesis of Diabetic Nephropathy: An Update. Vascul. Pharmacol. 2013, 58, 259–271. [Google Scholar] [CrossRef] [PubMed]
- Koya, D.; Jirousek, M.R.; Lin, Y.W.; Ishii, H.; Kuboki, K.; King, G.L. Characterization of Protein Kinase C β Isoform Activation on the Gene Expression of Transforming Growth Factor-β, Extracellular Matrix Components, and Prostanoids in the Glomeruli of Diabetic Rats. J. Clin. Investig. 1997, 100, 115–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayashida, T.; Schnaper, H.W. High Ambient Glucose Enhances Sensitivity to TGF-Β1 via Extracellular Signal-Regulated Kinase and Protein Kinase Cδ Activities in Human Mesangial Cells. J. Am. Soc. Nephrol. 2004, 15, 2032–2041. [Google Scholar] [CrossRef] [Green Version]
- Riser, B.L.; Denichilo, M.; Cortes, P.; Baker, C.; Grondin, J.M.; Yee, J.; Narins, R.G. Regulation of Connective Tissue Growth Factor Activity in Cultured Rat Mesangial Cells and Its Expression in Experimental Diabetic Glomerulosclerosis. J. Am. Soc. Nephrol. 2000, 11, 25–38. [Google Scholar] [CrossRef] [PubMed]
- Khamaisi, M.; Schrijvers, B.F.; De Vriese, A.S.; Raz, I.; Flyvbjerg, A. The Emerging Role of VEGF in Diabetic Kidney Disease. Nephrol. Dial. Transplant. 2003, 18, 1427–1430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoshi, S.; Tomari, S.; Hoshi, S.; Nomoto, K.I.; Kuromitsu, J.; Nagata, M. High Glucose Induced VEGF Expression via PKC and ERK in Glomerular Podocytes. Biochem. Biophys. Res. Commun. 2002, 290, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Xia, L.; Wang, H.; Munk, S.; Frecker, H.; Goldberg, H.J.; Fantus, I.G.; Whiteside, C.I. Reactive Oxygen Species, PKC-A1, and PKC-ζ Mediate High-Glucose-Induced Vascular Endothelial Growth Factor Expression in Mesangial Cells. Am. J. Physiol. Endocrinol. Metab. 2007, 293, 1280–1288. [Google Scholar] [CrossRef] [Green Version]
- Thallas-Bonke, V.; Thorpe, S.R.; Coughlan, M.T.; Fukami, K.; Yap, F.Y.T.; Sourris, K.C.; Penfold, S.A.; Bach, L.A.; Cooper, M.E.; Forbes, J.M. Inhibition of NADPH Oxidase Prevents Advanced Glycation End Product-Mediated Damage in Diabetic Nephropathy through a Protein Kinase C-α-Dependent Pathway. Diabetes 2008, 57, 460–469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishii, H.; Jirousek, M.R.; Koya, D.; Takagi, C.; Xia, P.; Clermont, A.; Bursell, S.E.; Kern, T.S.; Ballas, L.M.; Heath, W.F.; et al. Amelioration of Vascular Dysfunctions in Diabetic Rats by an Oral PKC β Inhibitor. Science 1996, 272, 728–731. [Google Scholar] [CrossRef]
- Koya, D.; Haneda, M.; Nakagawa, H.; Isshiki, K.; Sato, H.; Maeda, S.; Sugimoto, T.; Yasuda, H.; Kashiwagi, A.; Ways, D.K.; et al. Amelioration of Accelerated Diabetic Mesangial Expansion by Treatment with a PKC β Inhibitor in Diabetic Db/Db Mice, a Rodent Model for Type 2 Diabetes. FASEB J. 2000, 14, 439–447. [Google Scholar] [CrossRef] [Green Version]
- Sheetz, M.J.; Aiello, L.P.; Davis, M.D.; Danis, R.; Bek, T.; Cunha-Vaz, J.; Shahri, N.; Berg, P.H. The Effect of the Oral PKC β Inhibitor Ruboxistaurin on Vision Loss in Two Phase 3 Studies. Investig. Ophthalmol. Vis. Sci. 2013, 54, 1750–1757. [Google Scholar] [CrossRef] [PubMed]
- Meier, M.; Park, J.K.; Overheu, D.; Kirsch, T.; Lindschau, C.; Gueler, F.; Leitges, M.; Menne, J.; Haller, H. Deletion of Protein Kinase C-β Isoform in Vivo Reduces Renal Hypertrophy but Not Albuminuria in the Streptozotocin-Induced Diabetic Mouse Model. Diabetes 2007, 56, 346–354. [Google Scholar] [CrossRef] [Green Version]
- Kanoh, H.; Yamada, K.; Sakane, F. Diacylglycerol Kinase: A Key Modulator of Signal Transduction? Trends Biochem. Sci. 1990, 15, 47–50. [Google Scholar] [CrossRef]
- Sakane, F.; Imai, S.I.; Kai, M.; Yasuda, S.; Kanoh, H. Diacylglycerol Kinases: Why so Many of Them? Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2007, 1771, 793–806. [Google Scholar] [CrossRef] [PubMed]
- Topham, M.K.; Prescott, S.M. Mammalian diacylglycerol kinases, a Family of Lipid Kinases with Signaling Functions. J. Biol. Chem. 1999, 274, 11447–11450. [Google Scholar] [CrossRef] [Green Version]
- Van Blitterswijk, W.J.; Houssa, B. Properties and Functions of Diacylglycerol Kinases. Cell. Signal. 2000, 12, 595–605. [Google Scholar] [CrossRef]
- Kanoh, H.; Yamada, K.; Sakane, F. Diacylglycerol Kinases: Emerging Downstream Regulators in Cell Signaling Systems. J. Biochem. 2002, 131, 629–633. [Google Scholar] [CrossRef]
- Sakane, F.; Yamada, K.; Kanoh, H.; Yokoyama, C.; Tanabe, T. Porcine Diacylglycerol Kinase Sequence Has Zinc Finger and E-F Hand Motifs. Nature 1990, 344, 345–348. [Google Scholar] [CrossRef]
- Klauck, T.M.; Xu, X.; Mousseau, B.; Jaken, S. Cloning and Characterization of a Glucocorticoid-Induced Diacylglycerol Kinase. J. Biol. Chem. 1996, 271, 19781–19788. [Google Scholar] [CrossRef] [Green Version]
- Letwin, K.; Yee, S.P.; Pawson, T. Novel Protein-Tyrosine Kinase CDNAs Related to Fps/Fes and Eph Cloned Using Anti-Phosphotyrosine Antibody. Oncogene 1988, 3, 621–627. [Google Scholar]
- Imai, S.; Kai, M.; Yasuda, S.; Kanoh, H.; Sakane, F. Identification and Characterization of a Novel Human Type II Diacylglycerol Kinase, DGK Kappa. J. Biol. Chem. 2005, 280, 39870–39881. [Google Scholar] [CrossRef] [Green Version]
- Tang, W.; Bunting, M.; Zimmerman, G.A.; McIntyre, T.M.; Prescott, S.M. Molecular Cloning of a Novel Human Diacylglycerol Kinase Highly Selective for Arachidonate-Containing Substrates. J. Biol. Chem. 1996, 271, 10237–10241. [Google Scholar] [CrossRef]
- Bunting, M.; Tang, W.; Zimmerman, G.A.; McIntyre, T.M.; Prescott, S.M. Molecular Cloning and Characterization of a Novel Human Diacylglycerol Kinase Zeta. J. Biol. Chem. 1996, 271, 10230–10236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goto, K.; Kondo, H. A 104-KDa Diacylglycerol Kinase Containing Ankyrin-like Repeats Localizes in the Cell Nucleus. Proc. Natl. Acad. Sci. USA 1996, 93, 11196–11201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, L.; Traer, E.; McIntyre, T.M.; Zimmerman, G.A.; Prescott, S.M. The Cloning and Characterization of a Novel Human Diacylglycerol Kinase, DGKiota. J. Biol. Chem. 1998, 273, 32746–32752. [Google Scholar] [PubMed] [Green Version]
- Houssa, B.; Schaap, D.; Van Der Wal, J.; Goto, K.; Kondo, H.; Yamakawa, A.; Shibata, M.; Takenawa, T.; Van Blitterswijk, W.J. Cloning of a Novel Human Diacylglycerol Kinase (DGKθ) Containing Three Cysteine-Rich Domains, a Proline-Rich Region, and a Pleckstrin Homology Domain with an Overlapping Ras-Associating Domain. J. Biol. Chem. 1997, 272, 10422–10428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rigotti, A. Absorption, Transport, and Tissue Delivery of Vitamin E. Mol. Asp. Med. 2007, 28, 423–436. [Google Scholar] [CrossRef]
- Traber, M.G.; Atkinson, J. Vitamin E, Antioxidant and Nothing More. Free Radic. Biol. Med. 2007, 43, 4–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koya, D.; Lee, I.K.; Ishii, H.; Kanoh, H.; King, G.L. Prevention of Glomerular Dysfunction in Diabetic Rats by Treatment with D-Alpha-Tocopherol. J. Am. Soc. Nephrol. 1997, 8, 426–435. [Google Scholar]
- Tada, H.; Ishii, H.; Isogai, S. Protective Effect of D-α-Tocopherol on the Function of Human Mesangial Cells Exposed to High Glucose Concentrations. Metabolism 1997, 46, 779–784. [Google Scholar] [CrossRef]
- Kakehi, T.; Yagi, K.; Saito, N.; Shirai, Y. Effects of Vitamin E and Its Derivatives on Diabetic Nephropathy in Rats and Identification of Diacylglycerol Kinase Subtype Involved in the Improvement of Diabetic Nephropathy. Funct. Foods Health Dis. 2017, 7, 816–832. [Google Scholar] [CrossRef]
- Shirai, Y.; Segawa, S.; Kuriyama, M.; Goto, K.; Sakai, N.; Naoaki, S. Subtype-Specific Translocation of Diacylglycerol Kinase α and γ and Its Correlation with Protein Kinase C. J. Biol. Chem. 2000, 275, 24760–24766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
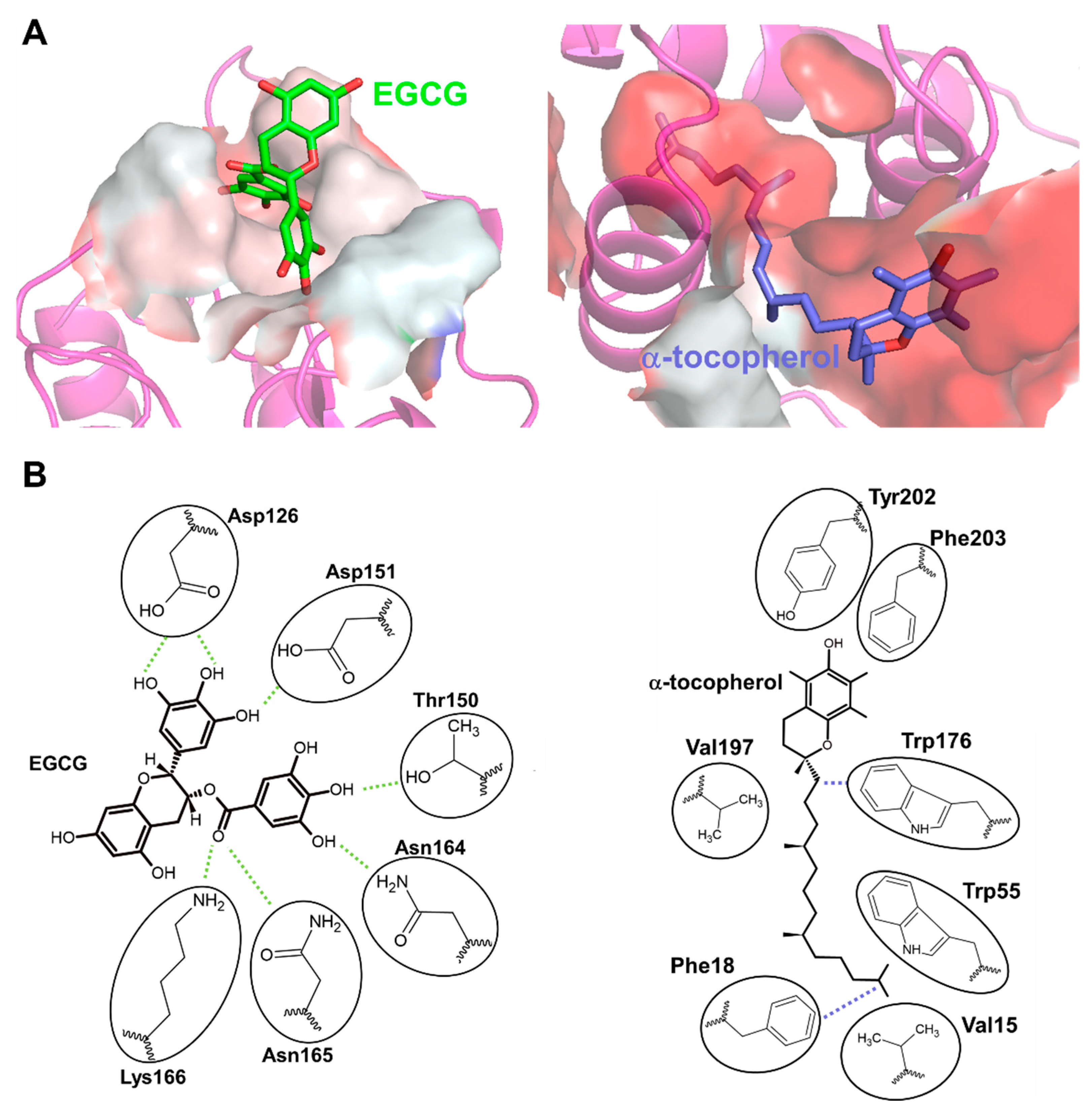
- Fukunaga-Takenaka, R.; Shirai, Y.; Yagi, K.; Adachi, N.; Sakai, N.; Merino, E.; Merida, I.; Saito, N. Importance of Chroman Ring and Tyrosine Phosphorylation in the Subtype-Specific Translocation and Activation of Diacylglycerol Kinase α by D-α-Tocopherol. Genes Cells 2005, 10, 311–319. [Google Scholar] [CrossRef] [PubMed]
- Tasinato, A.; Boscoboinik, D.; Bartoli, G.M.; Maroni, P.; Azzi, A. D-α-Tocopherol Inhibition of Vascular Smooth Muscle Cell Proliferation Occurs at Physiological Concentrations, Correlates with Protein Kinase C Inhibition, and Is Independent of Its Antioxidant Properties. Proc. Natl. Acad. Sci. USA 1995, 92, 12190–12194. [Google Scholar] [CrossRef] [Green Version]
- Pryor, W.A.; Cornicelli, J.A.; Devall, L.J.; Tait, B.; Trivedi, B.K.; Witiak, D.T.; Wu, M. A Rapid Screening Test To Determine the Antioxidant Potencies of Natural and Synthetic Antioxidants. J. Org. Chem. 1993, 58, 3521–3532. [Google Scholar] [CrossRef]
- Atsumi, H.; Kitada, M.; Kanasaki, K.; Koya, D. Reversal of Redox-Dependent Inhibition of Diacylglycerol Kinase by Antioxidants in Mesangial Cells Exposed to High Glucose. Mol. Med. Rep. 2011, 4, 923–927. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, D.; Yagi, K.; Song, C.; Ueda, S.; Yamanoue, M.; Topham, M.; Suzaki, T.; Saito, N.; Emoto, N.; Shirai, Y. Diacylglycerol Kinase Alpha Is Involved in the Vitamin E-Induced Amelioration of Diabetic Nephropathy in Mice. Sci. Rep. 2017, 7, 2597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vasavada, N.; Agarwal, R. Role of Oxidative Stress in Diabetic Nephropathy. Adv. Chronic Kidney Dis. 2005, 12, 146–154. [Google Scholar] [CrossRef]
- Kaneko, Y.K.; Kobayashi, Y.; Motoki, K.; Nakata, K.; Miyagawa, S.; Yamamoto, M.; Hayashi, D.; Shirai, Y.; Sakane, F.; Ishikawa, T. Depression of Type i Diacylglycerol Kinases in Pancreatic β-Cells from Male Mice Results in Impaired Insulin Secretion. Endocrinology 2013, 154, 4089–4098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chibalin, A.V.; Leng, Y.; Vieira, E.; Krook, A.; Björnholm, M.; Long, Y.C.; Kotova, O.; Zhong, Z.; Sakane, F.; Steiler, T.; et al. Downregulation of Diacylglycerol Kinase Delta Contributes to Hyperglycemia-Induced Insulin Resistance. Cell 2008, 132, 375–386. [Google Scholar] [CrossRef] [Green Version]
- Lonn, E.; Yusuf, S.; Hoogwerf, B.; Pogue, J.; Yi, Q.; Zinman, B.; Bosch, J.; Dagenais, G.; Mann, J.F.E.; Gerstein, H.C. Effects of Vitamin E on Cardiovascular and Microvascular Outcomes in High-Risk Patients With Diabetes Results of the HOPE Study and micro-hope substudy on behalf of the heart outcomes prevention evaluation (hope) investigators. Diabetes Care 2002, 25, 1919–1927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Vincenzo, A.; Tana, C.; El Hadi, H.; Pagano, C.; Vettor, R.; Rossato, M. Antioxidant, Anti-Inflammatory, and Metabolic Properties of Tocopherols and Tocotrienols: Clinical Implications for Vitamin E Supplementation in Diabetic Kidney Disease. Int. J. Mol. Sci. 2019, 20, 5101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, S.M.Q.; Chiew, Y.; Ahmad, B.; Kadir, K.A. Tocotrienol-Rich Vitamin E from Palm Oil (Tocovid) and Its Effects in Diabetes and Diabetic Nephropathy: A Pilot Phase II Clinical Trial. Nutrients 2018, 10, 1315. [Google Scholar] [CrossRef]
- Yokozawa, T.; Nakagawa, T.; Oya, T.; Okubo, T.; Juneja, L.R. Green Tea Polyphenols and Dietary Fibre Protect against Kidney Damage in Rats with Diabetic Nephropathy. J. Pharm. Pharmacol. 2010, 57, 773–780. [Google Scholar] [CrossRef]
- Yamabe, N.; Yokozawa, T.; Oya, T.; Kim, M. Therapeutic Potential of (-)-Epigallocatechin 3-O-Gallate on Renal Damage in Diabetic Nephropathy Model Rats. J. Pharmacol. Exp. Ther. 2006, 319, 228–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohabbulla Mohib, M.; Fazla Rabby, S.M.; Paran, T.Z.; Mehedee Hasan, M.; Ahmed, I.; Hasan, N.; Abu Taher Sagor, M.; Mohiuddin, S. Protective Role of Green Tea on Diabetic Nephropathy—A Review. Cogent. Biol. 2016, 2, 1248166. [Google Scholar] [CrossRef]
- Hayashi, D.; Ueda, S.; Yamanoue, M.; Saito, N.; Ashida, H.; Shirai, Y. Epigallocatechin-3-Gallate Activates Diacylglycerol Kinase Alpha via a 67 KDa Laminin Receptor: A Possibility of Galloylated Catechins as Functional Food to Prevent and/or Improve Diabetic Renal Dysfunctions. J. Funct. Foods 2015, 15, 561–569. [Google Scholar] [CrossRef]
- Hayashi, D.; Wang, L.; Ueda, S.; Yamanoue, M.; Ashida, H. The Mechanisms of Ameliorating Effect of a Green Tea Polyphenol on Diabetic Nephropathy Based on Diacylglycerol Kinase α. Sci. Rep. 2020, 10, 11790. [Google Scholar] [CrossRef]
- Borges, C.M.; Papadimitriou, A.; Duarte, D.A.; Lopes De Faria, J.M.; Lopes De Faria, J.B. The Use of Green Tea Polyphenols for Treating Residual Albuminuria in Diabetic Nephropathy: A Double-Blind Randomised Clinical Trial. Sci. Rep. 2016, 6, 28282. [Google Scholar] [CrossRef] [Green Version]
- Ueda, M.; Nishiumi, S.; Nagayasu, H.; Fukuda, I.; Yoshida, K.I.; Ashida, H. Epigallocatechin Gallate Promotes GLUT4 Translocation in Skeletal Muscle. Biochem. Biophys. Res. Commun. 2008, 377, 286–290. [Google Scholar] [CrossRef]
- Shahwan, M.; Alhumaydhi, F.; Ashraf, G.M.; Hasan, P.M.Z.; Shamsi, A. Role of Polyphenols in Combating Type 2 Diabetes and Insulin Resistance. Int. J. Biol. Macromol. 2022, 206, 567–579. [Google Scholar] [CrossRef]
- Isbrucker, R.A.; Edwards, J.A.; Wolz, E.; Davidovich, A.; Bausch, J. Safety Studies on Epigallocatechin Gallate (EGCG) Preparations. Part 2: Dermal, Acute and Short-Term Toxicity Studies. Food Chem. Toxicol. 2006, 44, 636–650. [Google Scholar] [CrossRef] [PubMed]
- Ullmann, U.; Haller, J.; Decourt, J.D.; Girault, J.; Spitzer, V.; Weber, P. Plasma-Kinetic Characteristics of Purified and Isolated Green Tea Catechin Epigallocatechin Gallate (EGCG) after 10 Days Repeated Dosing in Healthy Volunteers. Int. J. Vitam. Nutr. Res. 2004, 74, 269–278. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, D.; Ueda, S.; Yamanoue, M.; Ashida, H.; Shirai, Y. Amelioration of Diabetic Nephropathy by Oral Administration of D-α-Tocopherol and Its Mechanisms. Biosci. Biotechnol. Biochem. 2018, 82, 65–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohan, T.; Velusamy, P.; Chakrapani, L.N.; Srinivasan, A.K.; Singh, A.; Johnson, T.; Periandavan, K. Impact of EGCG Supplementation on the Progression of Diabetic Nephropathy in Rats: An Insight into Fibrosis and Apoptosis. J. Agric. Food Chem. 2017, 65, 8028–8036. [Google Scholar] [CrossRef] [PubMed]
- Mundel, P.; Kriz, W. Structure and Function of Podocytes: An Update. Anat. Embryol. 1995, 192, 385–397. [Google Scholar] [CrossRef] [PubMed]
- Mundel, P. Podocyte Biology and Response to Injury. J. Am. Soc. Nephrol. 2004, 13, 3005–3015. [Google Scholar] [CrossRef] [Green Version]
- Li, J.J.; Kwak, S.J.; Jung, D.S.; Kim, J.J.; Yoo, T.H.; Ryu, D.R.; Han, S.H.; Choi, H.Y.; Lee, J.E.; Moon, S.J.; et al. Podocyte Biology in Diabetic Nephropathy. Kidney Int. 2007, 72, S36–S42. [Google Scholar] [CrossRef] [Green Version]
- Quack, I.; Woznowski, M.; Potthoff, S.A.; Palmer, R.; Königshausen, E.; Sivritas, S.; Schiffer, M.; Stegbauer, J.; Vonend, O.; Rump, L.C.; et al. PKCα Mediates β-Arrestin2-Dependent Nephrin Endocytosis in Hyperglycemia. J. Biol. Chem. 2011, 286, 13959–13970. [Google Scholar] [CrossRef] [Green Version]
- Kerjaschki, D.; Ojha, P.P.; Susani, M.; Horvat, R.; Binder, S.; Hovorka, A.; Hillemanns, P.; Pytela, R. A Beta 1-Integrin Receptor for Fibronectin in Human Kidney Glomeruli. Am. J. Pathol. 1989, 134, 481–489. [Google Scholar]
- Adler, S. Characterization of Glomerular Epithelial Cell Matrix Receptors. Am. J. Pathol. 1992, 141, 571. [Google Scholar] [PubMed]
- Rainero, E.; Caswell, P.T.; Muller, P.A.J.; Grindlay, J.; Mccaffrey, M.W.; Zhang, Q.; Wakelam, M.J.O.; Vousden, K.H.; Graziani, A.; Norman, J.C. Diacylglycerol Kinase α Controls RCP-Dependent Integrin Trafficking to Promote Invasive Migration. J. Cell Biol. 2012, 196, 277–295. [Google Scholar] [CrossRef]
- Rainero, E.; Cianflone, C.; Porporato, P.E.; Chianale, F.; Malacarne, V.; Bettio, V.; Ruffo, E.; Ferrara, M.; Benecchia, F.; Capello, D.; et al. The Diacylglycerol Kinase α/Atypical PKC/Β1 Integrin Pathway in SDF-1α Mammary Carcinoma Invasiveness. PLoS ONE 2014, 9, 97144. [Google Scholar] [CrossRef] [PubMed]
- Dominguez, C.L.; Floyd, D.H.; Xiao, A.; Mullins, G.R.; Benjamin, A.; Xin, W.; Yacur, M.N.; Abounader, R.; Lee, J.K.; Mustata, G.; et al. Therapeutic Target in Glioblastoma and Other Cancers. Cancer Discov. 2013, 3, 782–797. [Google Scholar] [CrossRef] [PubMed]
- Yanagisawa, K.; Yasuda, S.; Kai, M.; Imai, S.I.; Yamada, K.; Yamashita, T.; Jimbow, K.; Kanoh, H.; Sakane, F. Diacylglycerol Kinase α Suppresses Tumor Necrosis Factor-α-Induced Apoptosis of Human Melanoma Cells through NF-ΚB Activation. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2007, 1771, 462–474. [Google Scholar] [CrossRef]
- Sakane, F.; Hoshino, F.; Ebina, M.; Sakai, H.; Takahashi, D. The Roles of Diacylglycerol Kinase α in Cancer Cell Proliferation and Apoptosis. Cancers 2021, 13, 5190. [Google Scholar] [CrossRef] [PubMed]
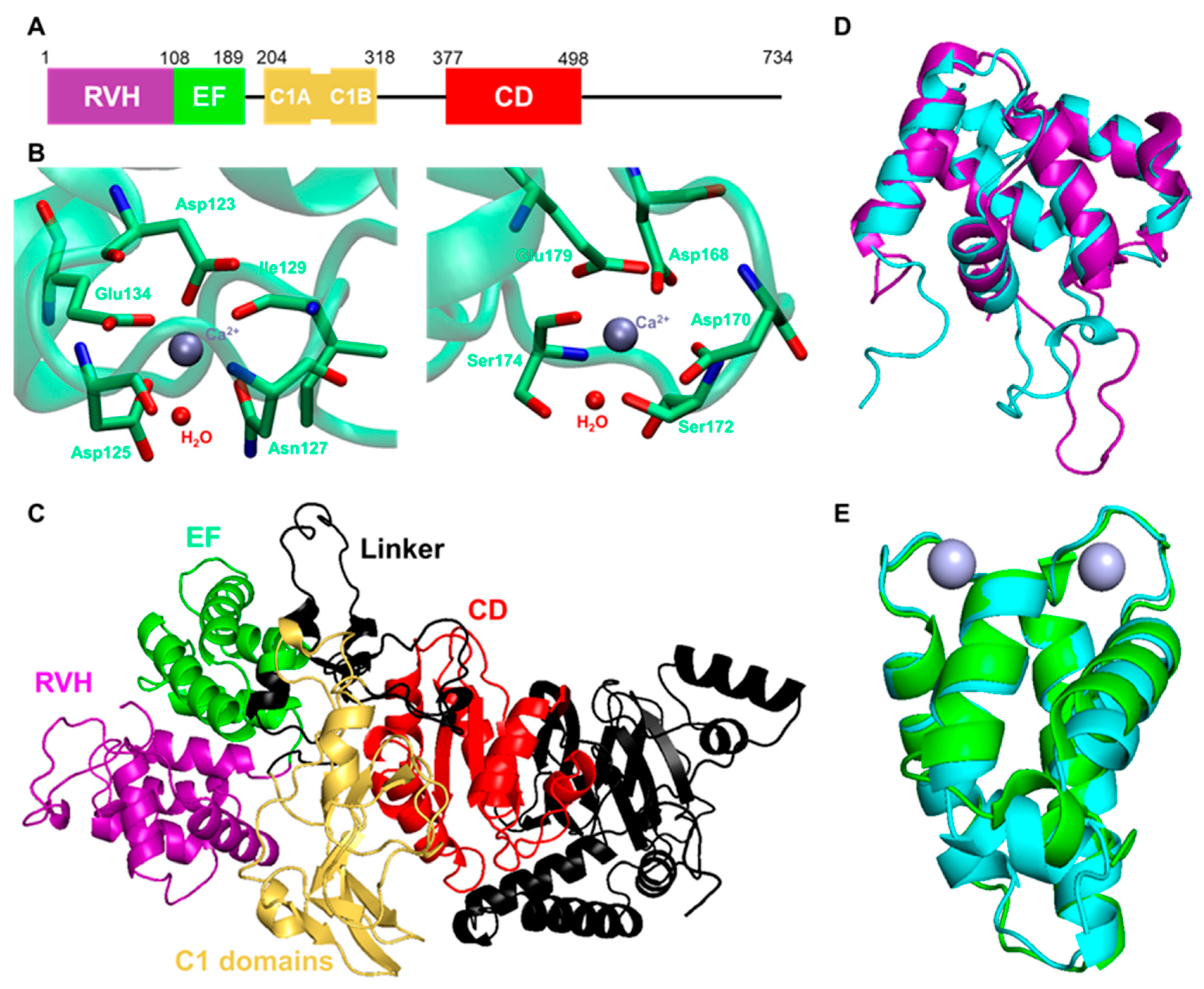
- Yamamoto, T.; Sakai, H.; Sakane, F. EF-Hand Motifs of Diacylglycerol Kinase α Interact Intra-Molecularly with Its C1 Domains. FEBS Open Biol. 2014, 4, 387–392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakane, F.; Yamada, K.; Imai, S.I.; Kanoh, H. Porcine 80-KDa Diacylglycerol Kinase Is a Calcium-Binding and Calcium/Phospholipid-Dependent Enzyme and Undergoes Calcium-Dependent Translocation. J. Biol. Chem. 1991, 266, 7096–7100. [Google Scholar] [CrossRef]
- Abe, T.; Lu, X.; Jiang, Y.; Boccone, C.E.; Qian, S.; Vattem, K.M.; Wek, R.C.; Walsh, J.P. Site-Directed Mutagenesis of the Active Site of Diacylglycerol Kinase α: Calcium and Phosphatidylserine Stimulate Enzyme Activity via Distinct Mechanisms. Biochem. J. 2003, 375, 673–680. [Google Scholar] [CrossRef] [Green Version]
- Ciprés, A.; Carrasco, S.; Merino, E.; Díaz, E.; Krishna, U.M.; Falck, J.R.; Martínez-A, C.; Mérida, I. Regulation of Diacylglycerol Kinase α by Phosphoinositide 3-Kinase Lipid Products. J. Biol. Chem. 2003, 278, 35629–35635. [Google Scholar] [CrossRef] [Green Version]
- Keiko, Y.; Fumio, S. The Different Effects of Sphingosine on Diacylglycerol Kinase Isozymes in Jurkat Cells, a Human T-Cell Line. Biochim. Biophys. Acta Lipids Lipid Metab. 1993, 1169, 211–216. [Google Scholar] [CrossRef]
- Keiko, Y.; Fumio, S.; Shin-ichi, I.; Haruo, T. Sphingosine Activates Cellular Diacylglycerol Kinase in Intact Jurkat Cells, a Human T-Cell Line. Biochim. Biophys. Acta Lipids Lipid Metab. 1993, 1169, 217–224. [Google Scholar] [CrossRef]
- Liu, K.; Kunii, N.; Sakuma, M.; Yamaki, A.; Mizuno, S.; Sato, M.; Sakai, H.; Kado, S.; Kumagai, K.; Kojima, H.; et al. A Novel Diacylglycerol Kinase α-Selective Inhibitor, CU-3, Induces Cancer Cell Apoptosis and Enhances Immune Response. J. Lipid Res. 2016, 57, 368–379. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, D.; Tsumagari, R.; Liu, K.; Ueda, S.; Yamanoue, M.; Sakane, F.; Shirai, Y. Screening of Subtype-Specific Activators and Inhibitors for Diacylglycerol Kinase. J. Biochem. 2019, 165, 517–522. [Google Scholar] [CrossRef]
- Takahashi, D.; Suzuki, K.; Sakamoto, T.; Iwamoto, T.; Murata, T.; Sakane, F. Crystal Structure and Calcium-Induced Conformational Changes of Diacylglycerol Kinase α EF-Hand Domains. Protein Sci. 2019, 28, 694–706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly Accurate Protein Structure Prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Varadi, M.; Anyango, S.; Deshpande, M.; Nair, S.; Natassia, C.; Yordanova, G.; Yuan, D.; Stroe, O.; Wood, G.; Laydon, A.; et al. AlphaFold Protein Structure Database: Massively Expanding the Structural Coverage of Protein-Sequence Space with High-Accuracy Models. Nucleic Acids Res. 2022, 50, D439–D444. [Google Scholar] [CrossRef]
- Aulakh, S.S.; Bozelli, J.C.; Epand, R.M. Exploring the AlphaFold Predicted Conformational Properties of Human Diacylglycerol Kinases. J. Phys. Chem. B 2022, 126, 7172–7183. [Google Scholar] [CrossRef] [PubMed]
- Merino, E.; Sanjuán, M.A.; Moraga, I.; Ciprés, A.; Mérida, I. Role of the Diacylglycerol Kinase α-Conserved Domains in Membrane Targeting in Intact T Cells. J. Biol. Chem. 2007, 282, 35396–35404. [Google Scholar] [CrossRef] [Green Version]
- Matsubara, T.; Ikeda, M.; Kiso, Y.; Sakuma, M.; Yoshino, K.I.; Sakane, F.; Merida, I.; Saito, N.; Shirai, Y. C-Abl Tyrosine Kinase Regulates Serum-Induced Nuclear Export of Diacylglycerol Kinase α by Phosphorylation at Tyr-218. J. Biol. Chem. 2012, 287, 5507–5517. [Google Scholar] [CrossRef] [Green Version]
- Azzi, A.; Stocker, A. Vitamin E: Non-Antioxidant Roles. Prog. Lipid Res. 2000, 39, 231–255. [Google Scholar] [CrossRef]
- Rao, N.C.; Barsky, S.H.; Terranova, V.P.; Liotta, L.A. Isolation of a Tumor Cell Laminin Receptor. Biochem. Biophys. Res. Commun. 1983, 111, 804–808. [Google Scholar] [CrossRef]
- Malinoff, H.L.; Wicha, M.S. Isolation of a Cell Surface Receptor Protein for Laminin from Murine Fibrosarcoma Cells. J. Cell Biol. 1983, 96, 1475–1479. [Google Scholar] [CrossRef]
- Lesot, H.; Kühl, U.; von der Mark, K.; Mark, K. Isolation of a Laminin-Binding Protein from Muscle Cell Membranes. EMBO J. 2018, 2, 861–865. [Google Scholar] [CrossRef]
- Gauczynski, S.; Peyrin, J.; Haõ, Â.; Leucht, C.; Hundt, C.; Rieger, R.; Krasemann, S.; Deslys, J.; Dormont, D.; Weiss, S.; et al. The 37-KDa/67-KDa Laminin Receptor Acts as the Cell-Surface Receptor for the Cellular Prion Protein. EMBO J. 2001, 20, 5863–5875. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.S.; Kuhn, R.J.; Strauss, E.G.; Ou, S.; Strauss, J.H. High-Affinity Laminin Receptor Is a Receptor for Sindbis Virus in Mammalian Cells. J. Virol. 1992, 66, 4992–5001. [Google Scholar] [CrossRef] [Green Version]
- Tachibana, H.; Koga, K.; Fujimura, Y.; Yamada, K. A Receptor for Green Tea Polyphenol EGCG. Nat. Struct. Mol. Biol. 2004, 11, 380–381. [Google Scholar] [CrossRef]
- Umeda, D.; Yano, S.; Yamada, K.; Tachibana, H. Green Tea Polyphenol Epigallocatechin-3-Gallate Signaling Pathway through 67-KDa Laminin Receptor. J. Biol. Chem. 2008, 283, 3050–3058. [Google Scholar] [CrossRef] [Green Version]
- Kumazoe, M.; Sugihara, K.; Tsukamoto, S.; Huang, Y.; Tsurudome, Y.; Suzuki, T.; Suemasu, Y.; Ueda, N.; Yamashita, S.; Kim, Y.; et al. 67-KDa Laminin Receptor Increases CGMP to Induce Cancer-Selective Apoptosis. J. Clin. Investig. 2013, 123, 787–799. [Google Scholar] [CrossRef] [PubMed]
- Hong Byun, E.; Fujimura, Y.; Yamada, K.; Tachibana, H. TLR4 Signaling Inhibitory Pathway Induced by Green Tea Polyphenol Epigallocatechin-3-Gallate through 67-KDa Laminin Receptor. J. Immunol. 2010, 185, 33–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsukamoto, S.; Huang, Y.; Umeda, D.; Yamada, S.; Yamashita, S.; Kumazoe, M.; Kim, Y.; Murata, M.; Yamada, K.; Tachibana, H. 67-KDa Laminin Receptor-Dependent Protein Phosphatase 2A (PP2A) Activation Elicits Melanoma-Specific Antitumor Activity Overcoming Drug Resistance. J. Biol. Chem. 2014, 289, 32671–32681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumazoe, M.; Kadomatsu, M.; Bae, J.; Otsuka, Y.; Fujimura, Y. Src Mediates Epigallocatechin-3-O-Gallate-Elicited Acid Sphingomyelinase Activation. Molecules 2020, 25, 5481. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, D.; Mouchlis, V.D.; Okamoto, S.; Namba, T.; Wang, L.; Li, S.; Ueda, S.; Yamanoue, M.; Tachibana, H.; Arai, H.; et al. Vitamin E Functions by Association with a Novel Binding Site on the 67 KDa Laminin Receptor Activating Diacylglycerol Kinase. J. Nutr. Biochem. 2022, 110, 109129. [Google Scholar] [CrossRef] [PubMed]
- Bhat, K.P.L.; Aldape, K. Not Expecting the Unexpected: Diacylglycerol Kinase Alpha as a Cancer Target. Cancer Discov. 2013, 3, 726–727. [Google Scholar] [CrossRef]
- Krishna, S.; Zhong, X.P. Regulation of Lipid Signaling by Diacylglycerol Kinases during T Cell Development and Function. Front. Immunol. 2013, 4, 178. [Google Scholar] [CrossRef] [Green Version]
- Velnati, S.; Ruffo, E.; Massarotti, A.; Talmon, M.; Varma, K.S.S.; Gesu, A.; Fresu, L.G.; Snow, A.L.; Bertoni, A.; Capello, D.; et al. Identification of a Novel DGKα Inhibitor for XLP-1 Therapy by Virtual Screening. Eur. J. Med. Chem. 2019, 164, 378–390. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hayashi, D.; Shirai, Y. The Role of Diacylglycerol Kinase in the Amelioration of Diabetic Nephropathy. Molecules 2022, 27, 6784. https://doi.org/10.3390/molecules27206784
Hayashi D, Shirai Y. The Role of Diacylglycerol Kinase in the Amelioration of Diabetic Nephropathy. Molecules. 2022; 27(20):6784. https://doi.org/10.3390/molecules27206784
Chicago/Turabian StyleHayashi, Daiki, and Yasuhito Shirai. 2022. "The Role of Diacylglycerol Kinase in the Amelioration of Diabetic Nephropathy" Molecules 27, no. 20: 6784. https://doi.org/10.3390/molecules27206784
APA StyleHayashi, D., & Shirai, Y. (2022). The Role of Diacylglycerol Kinase in the Amelioration of Diabetic Nephropathy. Molecules, 27(20), 6784. https://doi.org/10.3390/molecules27206784