
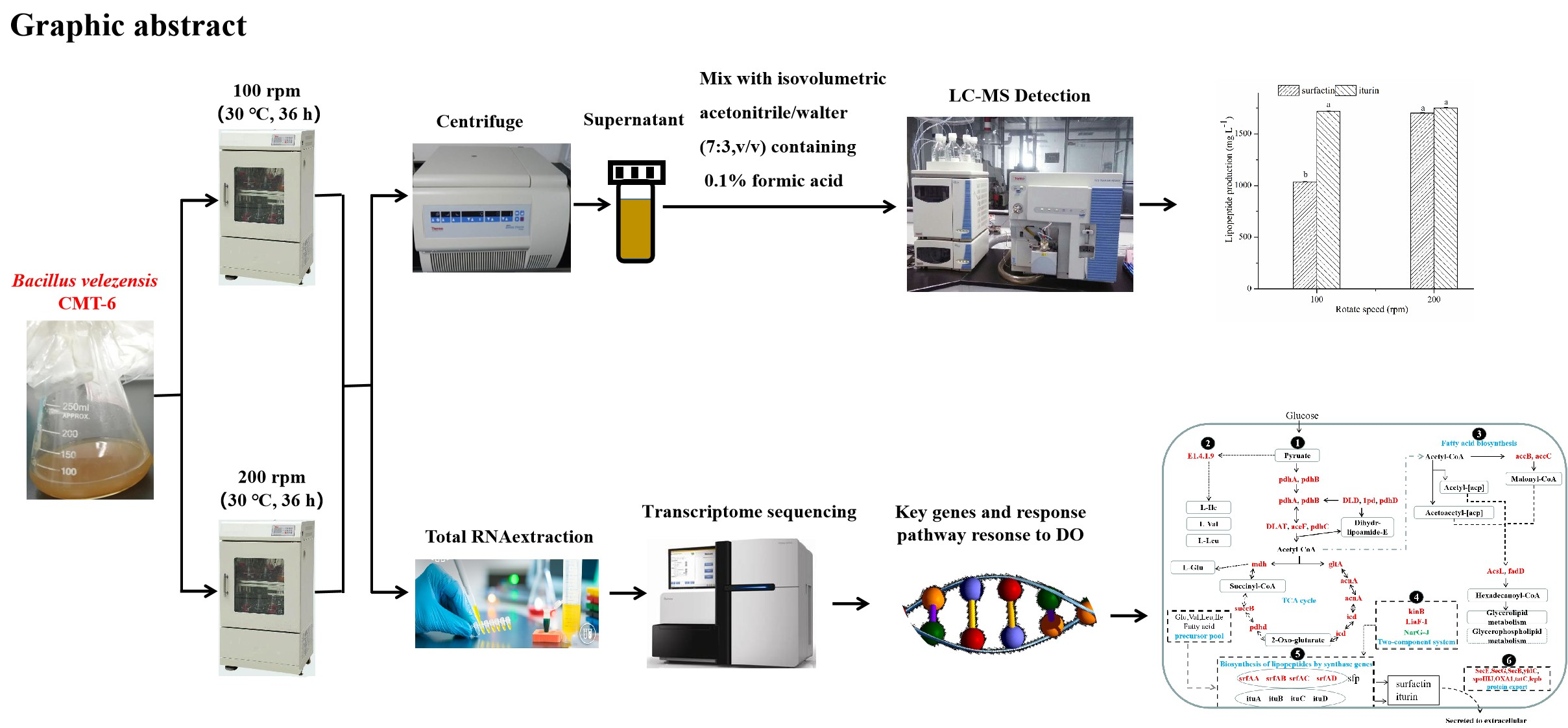
LC-MS and Transcriptome Analysis of Lipopeptide Biosynthesis by Bacillus velezensis CMT-6 Responding to Dissolved Oxygen

 , , ,
, , , 
Abstract
:
1. Introduction
2. Results and Discussion
2.1. Phenotypic Assays for Biomass and Bacillus velezensis CMT-6 Lipopeptide Production
2.2. Global Transcriptome Analysis
2.3. The Effect of Dissolved Oxygen on Synthetase Genes and the Key Genes Associated with Lipopeptides
3. Materials and Methods
3.1. Experimental Strain and Medium
3.2. Cell Growth and Lipopeptide Production Assay
3.3. RNA Extraction and Analysis of RNA Sequencing Data
3.3.1. RNA Extraction
3.3.2. Construction of the cDNA Library and Transcriptomic Data Analysis
3.4. Data Analysis
3.5. Enrichment Analysis
3.6. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Sun, D.F.; Liao, J.M.; Sun, L.J.; Wang, Y.L.; Liu, Y.; Deng, Q.; Zhang, N.; Xu, D.F.; Fang, Z.J.; Wang, W.J.; et al. Effect of Media and Fermentation Conditions on surfactin and iturin Homologues Produced by Bacillus natto NT-6: LC–MS analysis. AMB Express 2019, 9, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Deng, Q.; Wang, R.D.; Sun, L.D.; Wang, Y.L.; Pu, Y.H.; Fang, Z.J.; Xu, D.F.; Liu, Y.; Ye, R.Y.; Yin, S.J.; et al. Complete Genome of Bacillus velezensis CMT-6 and Comparative Genome Analysis Reveals Lipopeptide Diversity. Biochem. Genet. 2020, 58, 1–15. [Google Scholar] [CrossRef]
- Théatre, A.; Hoste, A.C.R.; Rigolet, A.; Benneceur, I.; Bechet, M.; Ongena, M.; Deleu, M.; Jacques, P. Bacillus sp.: A Remarkable Source of Bioactive Lipopeptides. Adv. Biochem. Eng. Biotechnol. 2022, 181, 123–179. [Google Scholar] [PubMed]
- Rajni, S.; Jagdish, S.; Neelam, V. Statistical Optimization and Comparative Study of Lipopeptides Produced by Bacillus amyloliquefaciens SAS-1 and Bacillus subtilis BR-15. Biocatal. Agric. Biotechnol. 2020, 25, 101575. [Google Scholar]
- Zhang, B.; Xu, L.L.; Ding, J.L.; Wang, M.Z.; Ge, R.; Zhao, H.F.; Zhang, B.L.; Fan, J.F. Natural Antimicrobial Lipopeptides Secreted by Bacillus spp. and Their Application in Food Preservation, A Critical Review. Trends Food Sci. Tech. 2022, 127, 26–37. [Google Scholar] [CrossRef]
- Adu, S.A.; Naughton, P.J.; Marchant, R.; Banat, I.M. Microbial Biosurfactants in Cosmetic and Personal Skincare Pharmaceutical Formulations. Pharmaceutics 2020, 12, 1099. [Google Scholar] [CrossRef]
- Nakamoto, H.; Yokoyama, Y.; Suzuki, T.; Miyamoto, Y.; Fujishiro, T.; Morikawa, M.; Miyata, Y. A Cyclic Lipopeptide Surfactin Is A Species-selective Hsp90 Inhibitor That Suppresses Cyanobacterial Growth. J. Biochem. 2021, 170, 255–264. [Google Scholar] [CrossRef] [PubMed]
- Li, J.Y.; Wang, L.L.; Yi, F.; Zhou, L.; Gang, H.Z.; Liu, J.F.; Yang, S.Z.; Mu, B.Z. Microbial Lipopeptide-Producing Strains and Their Metabolic Roles under Anaerobic Conditions. Microorganisms 2021, 9, 2030. [Google Scholar] [CrossRef]
- Jiang, M.L.; Qing, X.; Hu, S.Y.; He, P.H.; Chen, Y.G.; Cai, D.B.; Wu, Y.T.; Chen, S.W. Enhanced Aerobic Denitrification Performance with Bacillus licheniformis via Secreting Lipopeptide Biosurfactant Lichenysin. Chem. Eng. J. 2022, 434, 134686. [Google Scholar] [CrossRef]
- Sameh, F.; Krasimir, D.; Frederique, G.; Peggy, V.; Philippe, J.; Iordan, N. Impact of Energy Supply and Oxygen Transfer on Selective Lipopeptide Production by Bacillus subtilis BBG21. Bioresour. Technol. 2012, 126, 1–6. [Google Scholar]
- Sameh, F.; Krasimir, D.; Peggy, V.; Frédérique, G.; Guillaume, D.; Philippe, J.; Nikov, I. Oxygen Transfer in Three Phase Inverse Fluidized Bed Bioreactor During Biosurfactant Production by Bacillus subtilis. Biochem. Eng. J. 2013, 76, 70–76. [Google Scholar]
- Arlt, P.; Hashizume, H.; Igarashi, M.; Gross, H. Genome Sequence of Lysobacter sp. Strain BMK333-48F3, the Producer Strain of Potent Lipopeptide Antibiotics of the Tripropeptin Family. Microbiol. Resour Ann. 2021, 10, e0096921. [Google Scholar] [CrossRef] [PubMed]
- Narendra Kumar, P.; Swapna, T.H.; Sathi, R.K.; Archana, K.; Nageshwar, L.; Nalini, S.; Khan, M.Y.; Hameeda, B. Draft Genome Sequence of Bacillus amyloliquefaciens Strain RHNK22, Isolated from Rhizosphere with Biosurfactant (Surfactin, Iturin, and Fengycin) and Antifungal Activity. Genome Announc. 2016, 4, e01682-15. [Google Scholar] [CrossRef]
- Zhi, Y.; Wu, Q.; Xu, Y. Genome and Transcriptome Analysis of Surfactin Biosynthesis in Bacillus amyloliquefaciens MT45. Sci. Rep. 2017, 7, 40976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, P.P.; Lei, S.Z.; Xu, X.G.; Wang, R.; Jiang, C.M.; Shi, J.L. Effect of Medium Compositions on the Production of Antifungal Components by Bacillus velezensis. Food Ferment. Ind. 2020, 46, 147–151. [Google Scholar]
- Wu, Q.; Zhi, Y.; Xu, Y. Systematically Engineering The Biosynthesis of A Green Biosurfactant Surfactin by Bacillus subtilis 168. Metab. Eng. 2019, 52, 87–97. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Yang, H.; Zhang, D.L.; Li, X.; Yu, H.M.; Shen, Z.Y. Overexpression of Specific Proton Motive Force-Dependent Transporters Facilitate The Export of Surfactin in Bacillus Subtilis. J. Ind. Microbiol. Biotechnol. 2015, 42, 93–103. [Google Scholar] [CrossRef] [PubMed]
- Barale, S.S.; Ghane, S.G.; Sonawane, K.D. Purification and Characterization of Antibacterial Surfactin Isoforms Produced by Bacillus velezensis SK. AMB Express 2022, 12, 7–17. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.Y.; Zhao, H.Y.; Lu, Z.X. Research Progress in Biosynthesis and Application of Surfactin. J. Nanjing Agric. Univ. 2021, 44, 1024–1034. [Google Scholar]
- Valdés-Velasco, L.M.; Favela-Torres, E.; Théatre, A.; Arguelles-Arias, A.; Saucedo-Castañeda, J.G.; Jacques, P. Relationship Between Lipopeptide Biosurfactant and Primary Metabolite Production by Bacillus Strains in Solid-state and Submerged Fermentation. Bioresour. Technol. 2022, 345, 126566. [Google Scholar] [CrossRef]
- Gupta, V.; Jain, K.K.; Garg, R.; Malik, A.; Gulati, P.; Bhatnagar, R. Characterization of a Two Component System, Bas1213-1214, Important for Oxidative Stress in Bacillus anthracis. J. Cell Biochem. 2018, 119, 5761–5774. [Google Scholar] [CrossRef]
- Guo, Q.G.; Dong, L.H.; Wang, P.P.; Li, S.Z.; Zhao, W.S.; Lu, X.Y.; Zhang, X.Y.; Ma, P. The PhoR/PhoP Two-Component System Regulates Fengycin Production in Bacillus subtilis NCD-2 Under Low-Phosphate Conditions. J. Integr. Agr. 2018, 17, 149–157. [Google Scholar] [CrossRef] [Green Version]
- Somya, A.; Vikas, K.S.; Vatika, G.; Jaspreet, K.; Damini, S.; Abhinav, G.; Rakesh, B. Functional Characterization of PhoPR Two Component System and Its Implication in Regulating Phosphate Homeostasis in Bacillus anthracis. BBA-Gen. Subj. 2017, 1861, 2956–2970. [Google Scholar]
- Fan, M.; Huang, S.Q.; Li, Y.L.; Zhao, Y.J.; Fan, B. Flagella and Motility-related Properties of Bacillus: A Review. Microbiol. China 2022, 49, 1832–1845. [Google Scholar]
- Roggiani, M.; Dubnau, D. ComA, a Phosphorylated Response Regulator Protein of Bacillus subtilis, Binds to The Promoter Region of srfA. J. Bacteriol. 1993, 175, 3182–3187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neiditch, M.B.; Capodagli, G.C.; Prehna, G.; Federle, M.J. Genetic and Structural Analyses of RRNPP Intercellular Peptide Signaling of Gram- Positive Bacteria. Annu. Rev. Genet. 2017, 51, 311–333. [Google Scholar] [CrossRef]
- Zhang, Z.; Ding, Z.T.; Zhong, J.Y.; Shu, D.; Luo, D.; Yang, J.; Tan, H. Improvement of Iturin A Production in Bacillus subtilis ZK0 by Overexpression of the ComA and SigA Genes. Lett. Appl. Microbiol. 2017, 64, 452–458. [Google Scholar] [CrossRef]
- Rasiya, K.T.; Denoj, S. Iturin and Surfactin From the Endophyte Bacillus amyloliquefaciens Strain RKEA3 Exhibits Antagonism Against Staphylococcus aureus. Biocatal. Agric. Biotechnol. 2021, 36, 102125. [Google Scholar]
- Rahman, F.B.; Sarkar, B.; Moni, R.; Rahman, M.S. Molecular Genetics of Surfactin and Its Effects on Different Sub-populations of Bacillus subtilis. Biotechnol. Rep. 2021, 32, e00686. [Google Scholar] [CrossRef]
- Sun, J.; Liu, Y.; Lin, F.; Lu, Z.; Lu, Y. CodY, ComA, DegU and Spo0A Controlling Lipopeptides Biosynthesis in Bacillus amyloliquefaciens fmb. J. Appl. Microbiol. 2021, 131, 1289–1304. [Google Scholar] [CrossRef]
- Damian, N.; Maciej, J.; Marta, B.; Krzysztof, G.; Wojciech, K.; Derek, J.M. Ultrashort Cationic Lipopeptides–Effect of N-Terminal Amino Acid and Fatty Acid Type on Antimicrobial Activity and Hemolysis. Molecules 2020, 25, 257. [Google Scholar]
- Tan, W.; Yin, Y.; Wen, J.P. Increasing Fengycin Production by Strengthening the Fatty Acid Synthesis Pathway and Optimizing Fermentation Conditions. Biochem. Eng. J. 2022, 177, 108235. [Google Scholar] [CrossRef]
- Lozoya, O.A.; Martinez-Reyes, I.; Wang, T.Y.; Grenet, D.; Bushel, P.; Li, J.Y.; Chandel, N.; Woychik, R.P.; Santos, J.H. Mitochondrial Nicotinamide Adenine Dinucleotide Reduced (NADH) Oxidation Links the Tricarboxylic Acid (TCA) Cycle with Methionine Metabolism and Nuclear DNA Methylation. PLoS Biol. 2018, 16, e2005707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, L.S.; Guo, W.B.; Chen, X.H. Exogenous Addition of Alkanoic Acids Enhanced Production of Antifungal Lipopeptides in Bacillus amyloliquefaciens Pc3. Appl Microbiol. Biotechnol. 2019, 103, 5367–5377. [Google Scholar] [CrossRef] [PubMed]
- Kraas, F.I.; Helmetag, V.; Wittmann, M.; Strieker, M.; Marahiel, M.A. Functional Dissection of Surfactin Synthetase Initiation Module Reveals Insights Into The Mechanism of Lipoinitiation. Chem. Biol. 2010, 17, 872–880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, R.R.; Lei, S.Z.; Xu, X.G.; Jin, H.; Sun, H.; Zhao, X.X.; Pang, B.; Shi, J.L. Key Elements and Regulation Strategies of NRPSs for Biosynthesis of Lipopeptides by Bacillus. Appl. Microbiol. Biotechnol. 2020, 104, 8077. [Google Scholar] [CrossRef] [PubMed]
- Ikhsan, Z.S.; Oksana; Agustien, A.; Syamsuardi; Djamaan, A.; Mustafa, I. The Effect of Nitrogen Sources on Anti-phytopathogenic Activities Fermented Filtrate of Bacillus subtilis AAF2. IOP Conf. Ser. Earth Environ. Sci. 2019, 391, 012005. [Google Scholar] [CrossRef]
- Deng, Q.; Wang, W.J.; Sun, L.J.; Wang, Y.L.; Liao, J.M.; Xu, D.F.; Liu, Y.; Ye, R.Y.; Gooneratne, R. A Sensitive Method for Simultaneous Quantitative Determination of Surfactin and Iturin by LC-MS/MS. Anal. Bioanal. Chem. 2017, 409, 179–191. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Gene Expression | Gene Function | |
|---|---|---|---|
| 100 rpm | 200 rpm | ||
| pckA | 43.66 | 129.17 | phosphoenolpyruvate carboxykinase (ATP) |
| aceF, pdhC | 6.10 | 3284.60 | dihydrolipoamide acetyltransferase |
| pdhA | 5.75 | 2596.92 | pyruvate dehydrogenase E1 component beta subunit |
| pdhB | 2.74 | 1931.38 | |
| lpd | 8.72 | 4663.19 | dihydrolipoamide dehydrogenase |
| pdhD | 39.09 | 82.20 | |
| mdh | 725.33 | 1505.83 | malate dehydrogenase |
| gltA | 511.65 | 1315.62 | citrate synthetase |
| acnA | 349.26 | 811.09 | aconitate hydratase |
| sdhB, frdB | 399.27 | 582.47 | succinate dehydrogenase |
| sucB | 438.12 | 783.63 | dihydrolipoamide succinyltransferase |
| icD | 683.05 | 1187.85 | isocitrate dehydrogenase |
| Gene | Gene Expression | Gene Function | |
|---|---|---|---|
| 100 rpm | 200 rpm | ||
| accC | 52.23 | 169.80 | acetyl-CoA carboxylase, biotin carboxylase subunit |
| accB | 4.77 | 14.25 | acetyl-CoA carboxylase biotin carboxyl carrier protein |
| acsL | 77.05 | 151.61 | long-chain acyl-CoA synthetase |
| fadD | 83.39 | 377.19 | long-chain acyl-CoA synthetase |
| Gene | Gene Expression | Gene Function | |
|---|---|---|---|
| 100 rpm | 200 rpm | ||
| kinC | 79.87 | 47.95 | sporulation sensor kinase C |
| kinD | 90.39 | 33.96 | sporulation sensor kinase D |
| narG | 19.12 | 1.83 | nitrate reductase alpha subunit |
| narH | 7.29 | 1.27 | nitrate reductase beta subunit |
| narI | 12.36 | 3.01 | nitrate reductase gamma subunit |
| narJ | 8.20 | 2.72 | nitrate reductase delta subunit |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deng, Q.; Lin, H.; Hua, M.; Sun, L.; Pu, Y.; Liao, J.; Fang, Z.; Zhong, S.; Gooneratne, R. LC-MS and Transcriptome Analysis of Lipopeptide Biosynthesis by Bacillus velezensis CMT-6 Responding to Dissolved Oxygen. Molecules 2022, 27, 6822. https://doi.org/10.3390/molecules27206822
Deng Q, Lin H, Hua M, Sun L, Pu Y, Liao J, Fang Z, Zhong S, Gooneratne R. LC-MS and Transcriptome Analysis of Lipopeptide Biosynthesis by Bacillus velezensis CMT-6 Responding to Dissolved Oxygen. Molecules. 2022; 27(20):6822. https://doi.org/10.3390/molecules27206822
Chicago/Turabian StyleDeng, Qi, Haisheng Lin, Meifang Hua, Lijun Sun, Yuehua Pu, Jianmeng Liao, Zhijia Fang, Saiyi Zhong, and Ravi Gooneratne. 2022. "LC-MS and Transcriptome Analysis of Lipopeptide Biosynthesis by Bacillus velezensis CMT-6 Responding to Dissolved Oxygen" Molecules 27, no. 20: 6822. https://doi.org/10.3390/molecules27206822
APA StyleDeng, Q., Lin, H., Hua, M., Sun, L., Pu, Y., Liao, J., Fang, Z., Zhong, S., & Gooneratne, R. (2022). LC-MS and Transcriptome Analysis of Lipopeptide Biosynthesis by Bacillus velezensis CMT-6 Responding to Dissolved Oxygen. Molecules, 27(20), 6822. https://doi.org/10.3390/molecules27206822