
Phenolic Profiles, Antioxidant and Anti-Inflammatory Activities of Hydrodistillation Wastewaters from Five Lamiaceae Species

 , , , ,
, , , ,  ,
,  and
and 
Abstract
:1. Introduction
2. Results
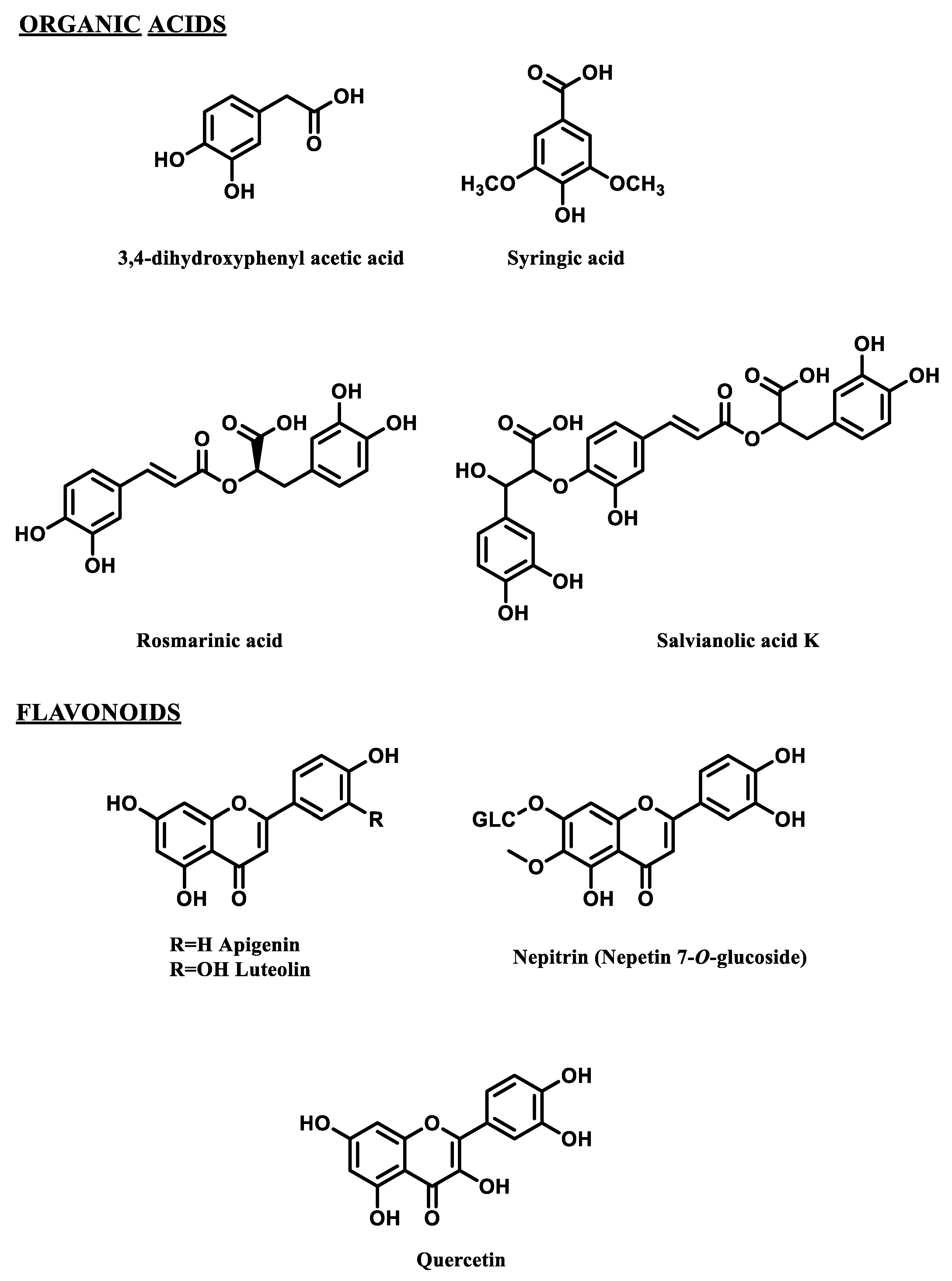
2.1. HPLC Polyphenolic Profiles of Wastewaters
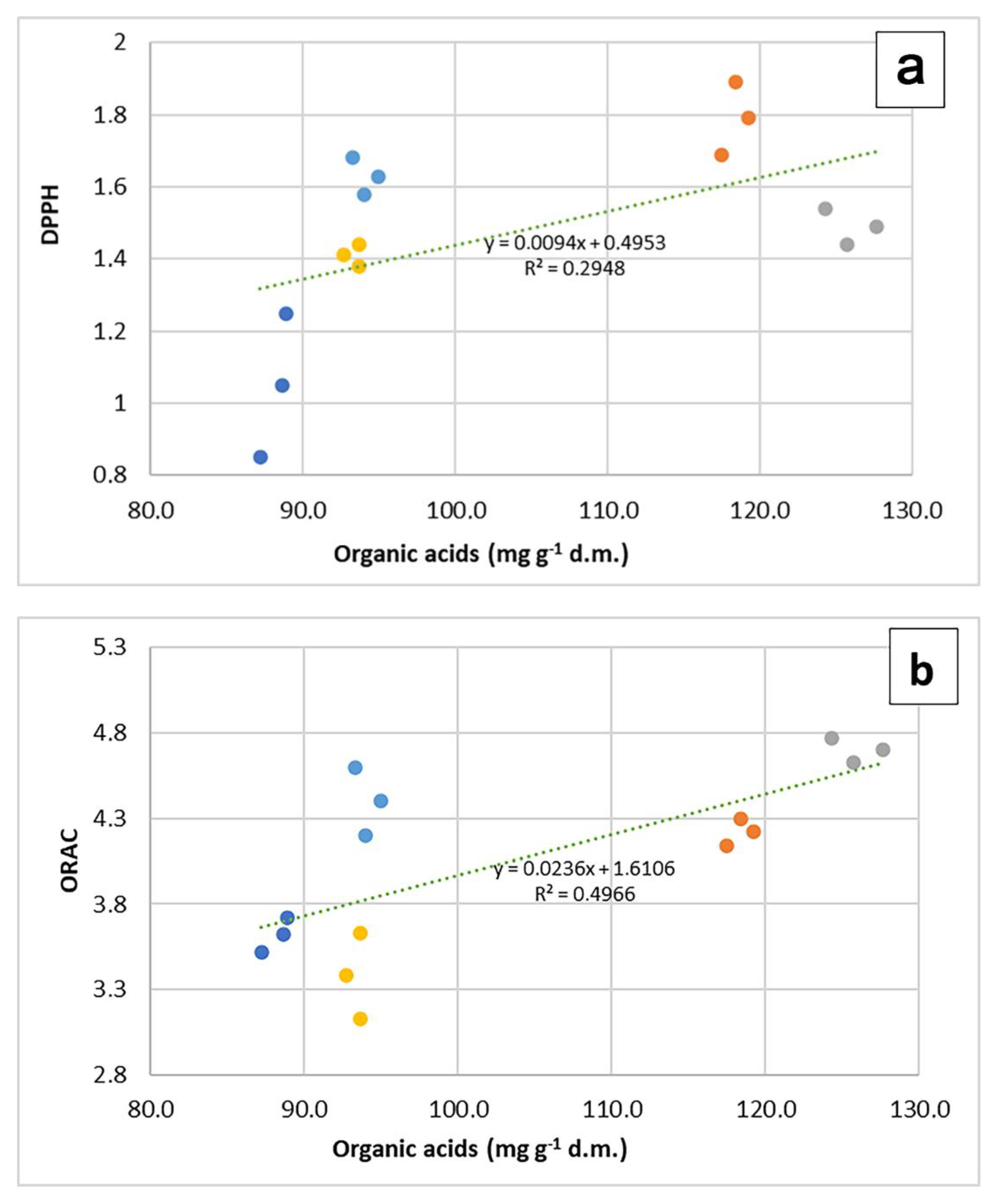
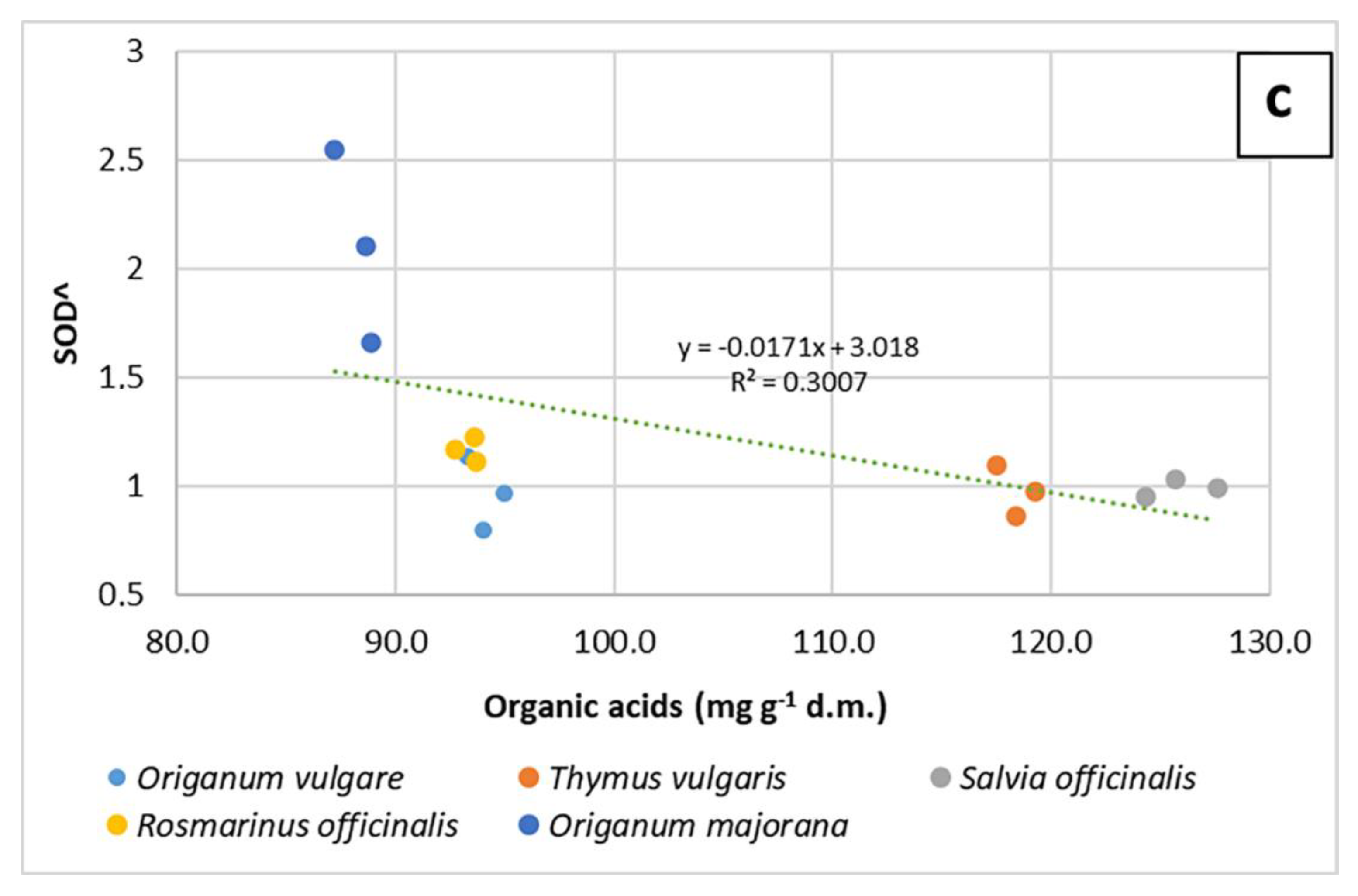
2.2. Evaluation of the Antioxidant Properties
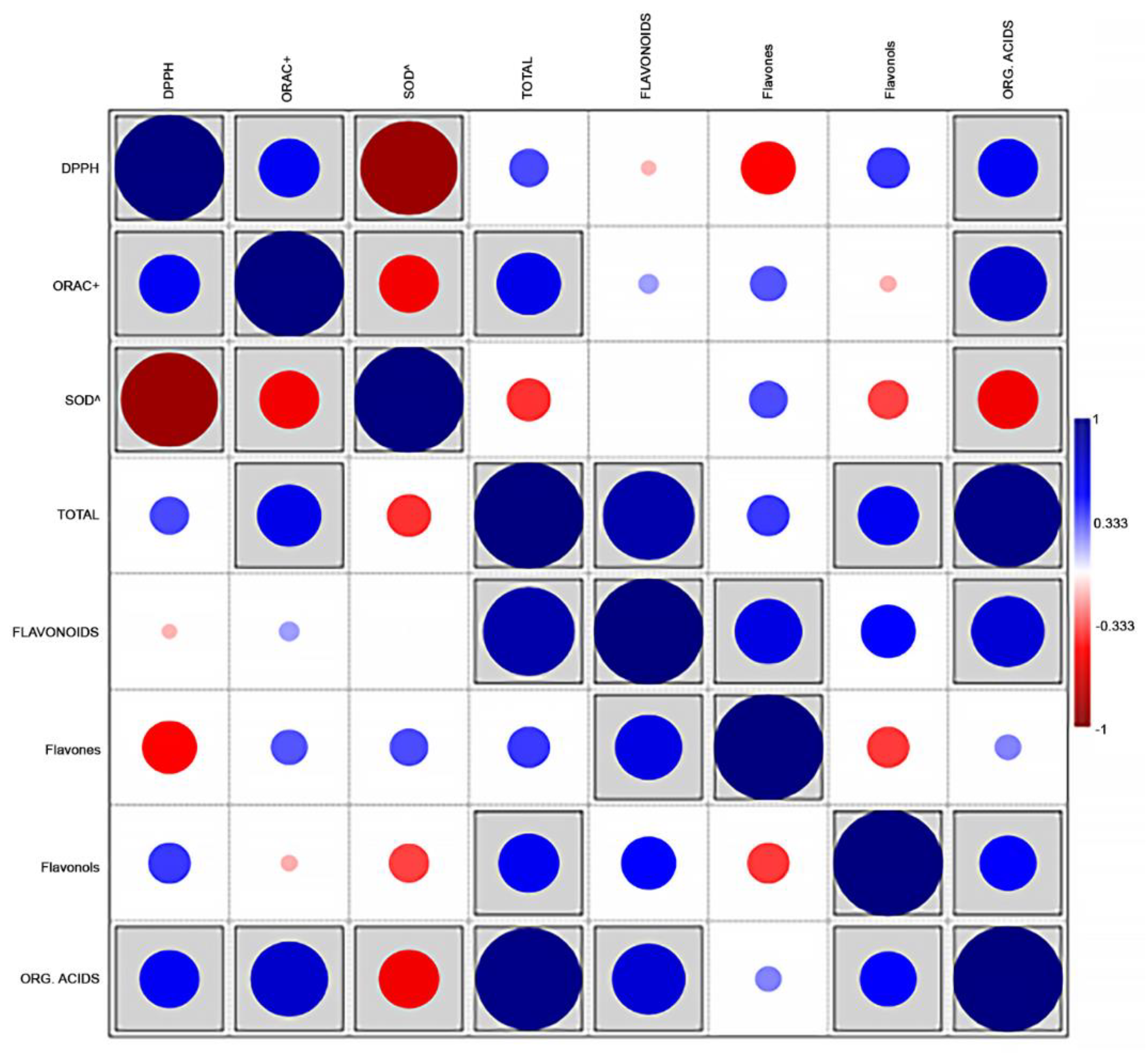
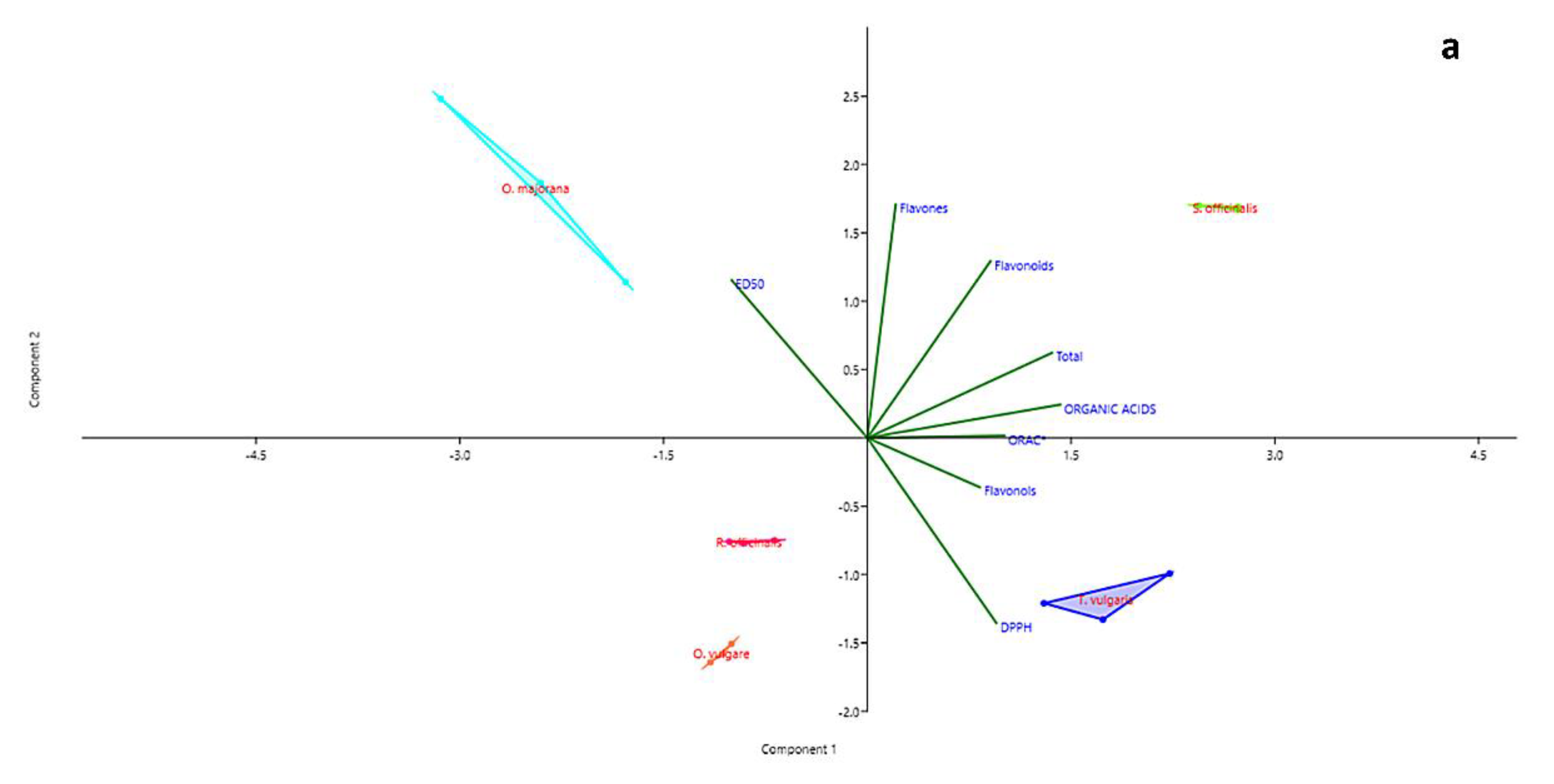
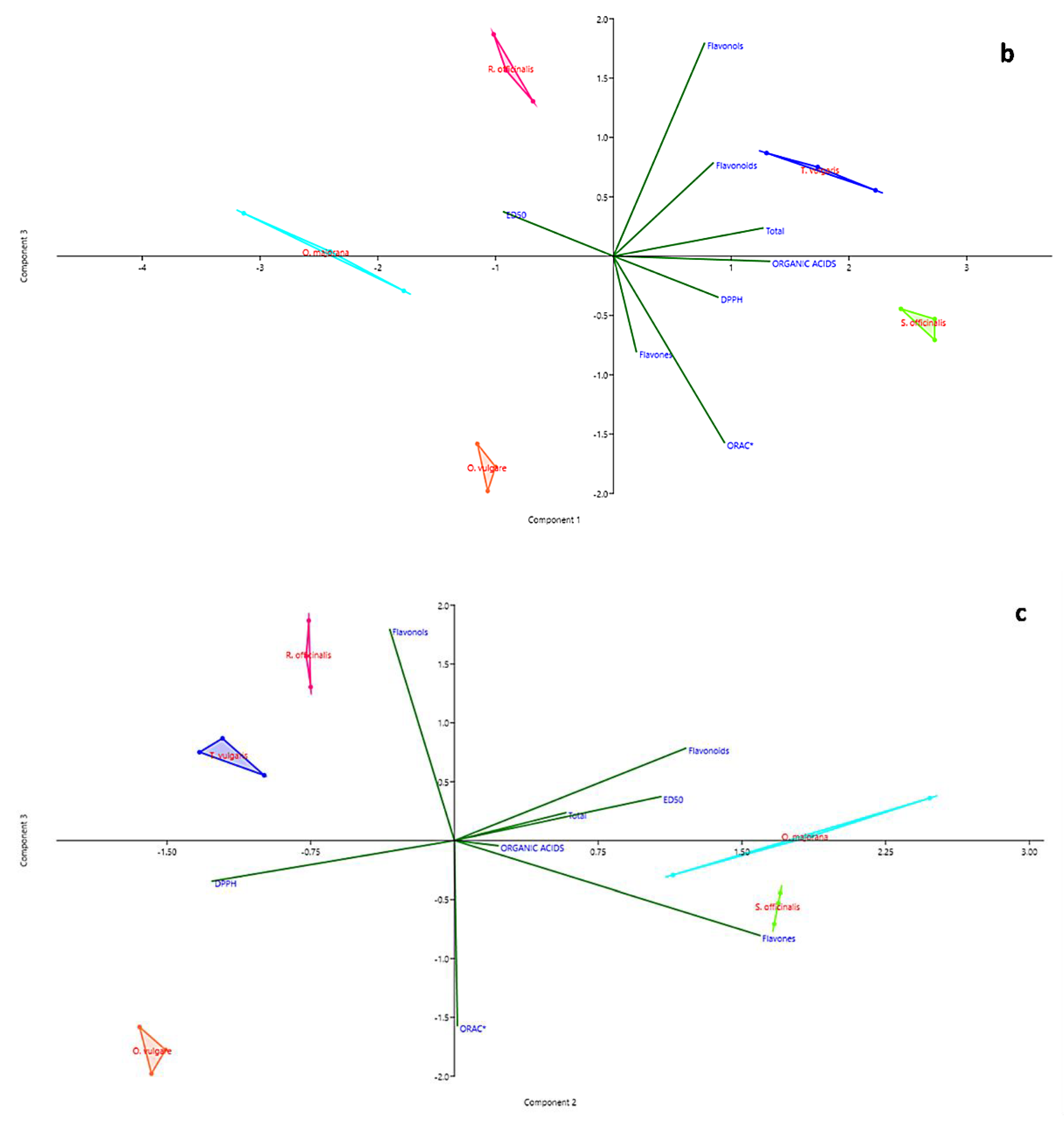
2.3. Statistical Analysis
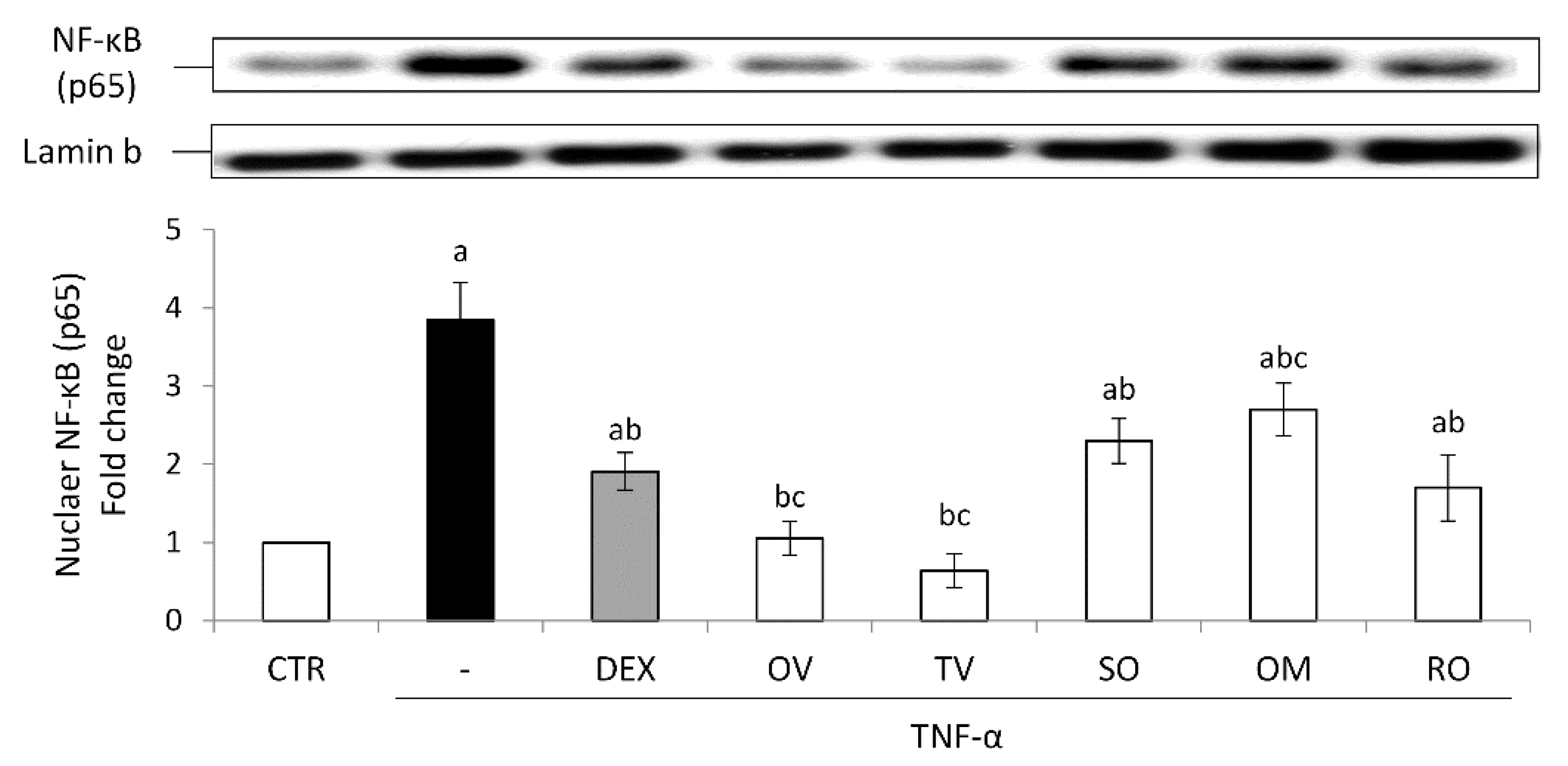
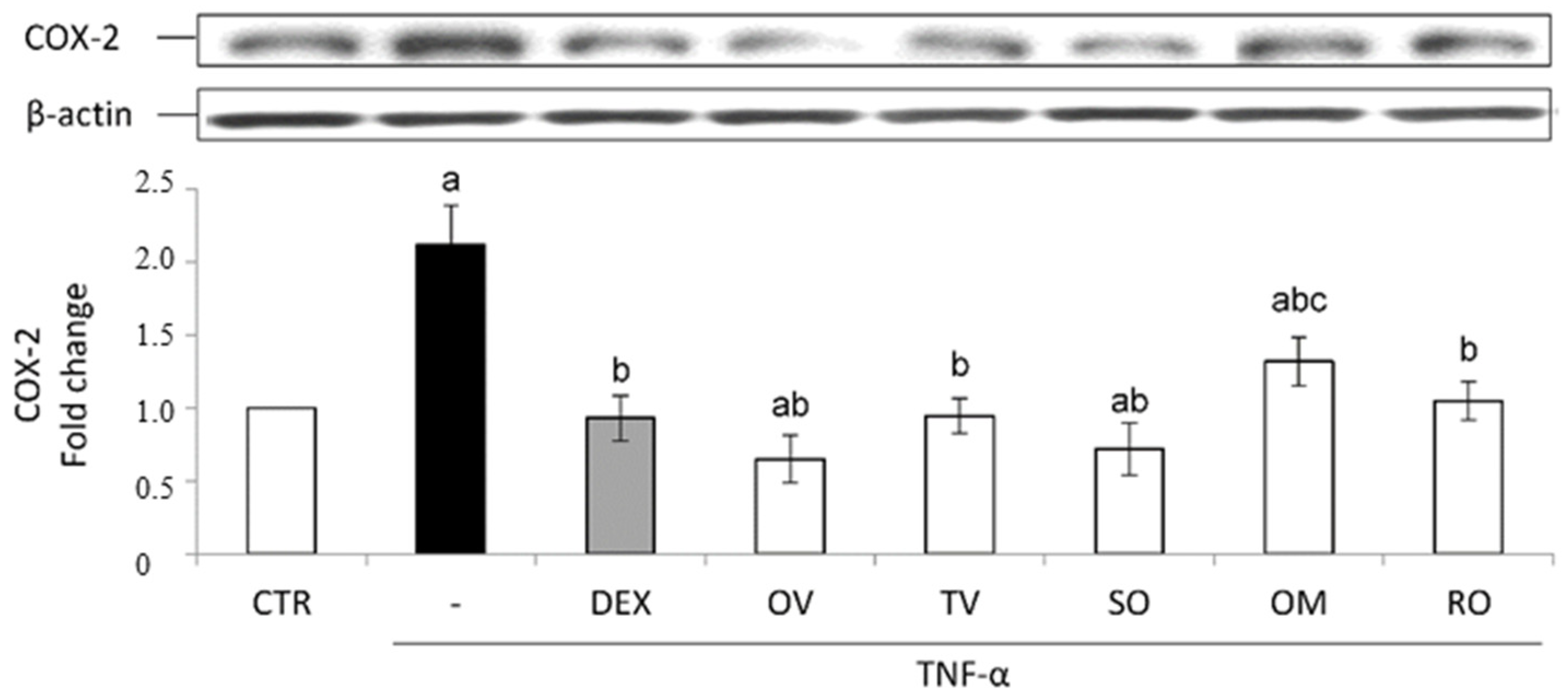
2.4. Anti-Inflammatory Activity in Intestinal Epithelial Cells Exposed to TNF-α
3. Discussion
4. Materials and Methods
4.1. General
4.2. Plant Material
4.3. Plant Hydrodistillation and Wastewater Recovery
4.4. Hydrodistillation Wastewaters Analysis
4.5. 2,2-Diphenyl-1-Picrylhydrazyl Free Radical (DPPH) Assay
4.6. Oxygen Radical Absorbance Capacity Assay
4.7. Superoxide Anion Scavenging Activity
4.8. TNF-α Induced Inflammation on Intestinal Epithelial Cells
4.8.1. Cell Cultures
4.8.2. Whole Cell and Nuclear Lysates’ Extraction
4.8.3. Western Blot Analysis
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Napoli, E.; Siracusa, L.; Ruberto, G. New tricks for old guys: Recent developments in the chemistry, biochemistry, applications and exploitations of selected species from Lamiaceae family. Chem. Biodivers. 2020, 17, e1900677. [Google Scholar] [CrossRef] [PubMed]
- Napoli, E.; Giovino, A.; Carrubba, A.; Siong, V.H.Y.; Rinoldo, C.; Nina, O.; Ruberto, G. Variations of essential oil constituents in oregano (Origanum vulgare subsp. viridulum (= O. heracleoticum) over cultivation cycles. Plants 2020, 9, 1174. [Google Scholar] [CrossRef] [PubMed]
- Saija, A.; Speciale, A.; Trombetta, D.; Leto, C.; Tuttolomondo, T.; La Bella, S.; Licata, M.; Bonsangue, G.; Gennaro, M.C.; Napoli, E.; et al. Phytochemical, ecological and antioxidant evaluation of wild Sicilian thyme: Thymbra capitata (L.) Cav. Chem. Biodivers. 2016, 13, 1641–1655. [Google Scholar] [CrossRef] [PubMed]
- Napoli, E.; Siracusa, L.; Ruberto, G.; Carrubba, A.; Lazzara, S.; Speciale, A.; Cimino, F.; Saija, A.; Cristani, M. Phytochemical profiles, phototoxic and antioxidant properties of eleven Hypericum species—A comparative study. Phytochemistry 2018, 152, 162–173. [Google Scholar] [CrossRef]
- Truzzi, E.; Chaouch, M.A.; Rossi, G.; Tagliazucchi, L.; Bertelli, D.; Benvenuti, S. Characterization and Valorization of the Agricultural Waste Obtained from Lavandula Steam Distillation for Its Reuse in the Food and Pharmaceutical Fields. Molecules 2022, 27, 1613. [Google Scholar] [CrossRef]
- Chavdar, C.; Evgeni, S.; Borislava, D.; Velislava, Y.; Swantje, P.; Stefan, H.; Dimitar, P. Valorization of waste lavender residue from the essential oil industry for production of rosmarinico acid—A study on the solid-liquid extraction. J. Chem. Technol. Metall. 2022, 57, 522–532. [Google Scholar]
- Schmidt, E. Production of Essential Oils. In Handbook of Essential Oils, Science, Technology, and Applications; Başer, K.H.C., Buchbauer, G., Eds.; CRC Press: Boca Raton, FL, USA, 2020; Chapter 4; pp. 83–120. [Google Scholar]
- ISO 3218:2014; Essential Oils–Principles of Nomenclature. ISO: Geneva, Switzerland, 2014.
- Kubeczka, K.-H. History and Sources of Essential Oil Research. In Handbook of Essential Oils, Science, Technology, and Applications; Başer, K.H.C., Buchbauer, G., Eds.; CRC Press: Boca Raton, FL, USA, 2020; Chapter 2; pp. 3–38. [Google Scholar]
- European Pharmacopoeia, 10th ed.; European Council: Strasbourg, France, 2019.
- Cid-Perez, T.S.; Avila-Sosa, R.; Ochoa-Velasco, C.E.; Rivera-Chavira, B.E.; Nevarez-Moorillon, G.V. Antioxidant and antimicrobial activity of Mexican oregano (Poliomintha longiflora) essential oil, hydrosol and extracts from waste solid residue. Plants 2019, 8, 22. [Google Scholar] [CrossRef] [Green Version]
- Fascella, G.; D’Angiolillo, F.; Ruberto, G.; Napoli, E. Agronomic performance, essential oils and hydrodistillation wastewaters of Lavandula angustifolia grown on biochar-based substrates. Ind. Crops Prod. 2020, 154, 112733. [Google Scholar] [CrossRef]
- Artasensi, A.; Pedretti, A.; Vistoli, G.; Fumagalli, L. Type 2 Diabetes Mellitus: A review of multi-target drugs. Molecules 2020, 25, 1987. [Google Scholar] [CrossRef]
- Eghbaliferiz, S.; Iranshahi, M. Pro-oxidant activity of polyphenols, flavonoids, anthocyanins and carotenoids: Updated review of mechanisms and catalyzing metals. Phytother. Res. 2016, 30, 1379–1391. [Google Scholar] [CrossRef]
- Chu, K.O.; Chan, S.O.; Pang, C.P.; Wang, C.C. Pro-oxidative and antioxidative controls and signaling modification of polyphenolic phytochemicals: Contribution to health promotion and disease prevention? J. Agric. Food Chem. 2014, 62, 4026–4038. [Google Scholar] [CrossRef] [PubMed]
- Caesar, L.K.; Cech, N.B. Synergy and antagonism in natural product extracts: When 1 + 1 does not equal 2. Nat. Prod. Rep. 2019, 36, 869–888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, J.; Mumper, R.J. Plant phenolics: Extraction, analysis and their antioxidant and anticancer properties. Molecules 2010, 15, 7313–7352. [Google Scholar] [CrossRef] [PubMed]
- Prior, R.L.; Wu, X.; Schaich, K. Standardized methods for the determination of antioxidant capacity and phenolics in foods and dietary supplement. J. Agric. Food Chem. 2005, 53, 4290–4302. [Google Scholar] [CrossRef]
- Dudonné, S.; Vitrac, X.; Couttière, P.; Woillez, M.; Mérillon, J.-M. Comparative study of antioxidant properties and total phenolic content of plant extracts of industrial interest using DPPH, ABTS, FRAP, SOD, and ORAC assays. J. Agric. Food Chem. 2009, 57, 1768–1774. [Google Scholar] [CrossRef]
- Jiang, W.; Hu, M. Mutual interactions between flavonoids and enzymatic and transporter elements responsible for flavonoid disposition via phase II metabolic pathways. RSC Adv. 2012, 2, 7948–7963. [Google Scholar] [CrossRef] [Green Version]
- Xie, L.; Deng, Z.; Zhang, J.; Dong, H.; Wang, W.; Xing, B.; Liu, X. Comparison of flavonoid O-glycoside, C-glycoside and their aglycones on antioxidant capacity and metabolism during in vitro digestion and in vivo. Foods 2022, 11, 882. [Google Scholar] [CrossRef]
- Speisky, H.; Shahidi, F.; de Camargo, A.C.; Fuentes, J. Revisiting the oxidation of flavonoids: Loss, conservation or enhancement of their antioxidant properties. Antioxidants 2022, 11, 133. [Google Scholar] [CrossRef]
- Chen, C.P.; Yokozawa, T.; Chung, H.Y. Inhibitory effect of caffeic acid analogues isolated from Salviae miltiorrhizae radix against 1,1-dipehnyl-2-picrylhydrazyl radical. Exp. Tpxicol. Pathol. 1999, 51, 59–63. [Google Scholar] [CrossRef]
- Tang, Y.; Nakashima, S.; Saiki, S.; Myoi, Y.; Abe, N.; Kuwazuru, S.; Zhu, B.; Ashida, H.; Murata, Y.; Nakamura, Y. 3,4-Dihydroxyphenylacetic acid is a predominant biologically-active catabolite of quercetin glycosides. Food Res. Int. 2016, 89, 716–723. [Google Scholar] [CrossRef]
- Kaulmann, A.; Bohn, T. Bioactivity of polyphenols: Preventive and adjuvant strategies toward reducing inflammatory Bowel disease–promises, perspectives, and pitfalls. Oxidative Med. Cell. Long. 2016, 9346470. [Google Scholar] [CrossRef] [PubMed]
- Arya, V.S.; Kanthal, S.K.; Linda, G. The role of dietary polyphenols in inflammatory bowel disease: A possible clue on the molecular mechanism involved in the prevention of immune and inflammatory response. J. Food Biochem. 2020, 44, e13369. [Google Scholar] [CrossRef]
- Műzes, G.; Molnár, B.; Tulassay, Z.; Sipos, F. Changes of the cytokine profile in inflammatory bowel diseases. World J. Gastroenterol. 2012, 18, 5848–5861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayden, M.S.; Ghosh, S. Signalling to NF-kappaB. Genes Dev. 2004, 18, 2195–2224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsatsanis, C.; Androulidaki, A.; Venihaki, M.; Margioris, A.N. Signalling networks regulating cyclooxygenase-2. Int. J. Biochem. Cell. Biol. 2006, 38, 1654–1661. [Google Scholar] [CrossRef] [PubMed]
- Sheibanie, A.F.; Yen, J.-H.; Khayrullina, T.; Emig, F.; Zhang, M.; Tuma, R.; Ganea, D. The pro-inflammatory effect of prostaglandin E2 in experimental inflammatory bowel disease is mediated through the IL-23—IL-17 axis. J. Immunol. 2007, 178, 8138–8147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Oliveira Formiga, R.; Alves, E.B.; Vasconcelos, R.C.; Guerra, G.C.B.; de Araùjo, A.A.; de Carvalho, T.G.; Garcia, G.C.; de Araùjo, R.F.; Gadelha, F.A.A.F.; Vieira, G.C.; et al. p-Cymene and rosmarinic acid ameliorate TNBS-induced intestinal inflammation upkeeping ZO-1 and MUC-2: Role of antioxidant system and immunomodulation. Int. J. Mol. Sci. 2020, 21, 5870. [Google Scholar] [CrossRef]
- Chung, C.H.; Jung, W.; Keum, H.; Kim, T.W.; Jon, S. Nanoparticles derived from the natural antioxidant rosmarinic acid ameliorate acute inflammatory bowel disease. ACS Nano 2020, 14, 6887–6896. [Google Scholar] [CrossRef]
- Xiang, S.; Xiao, J. Protective effects of syringic acid on inflammation, apoptosis and intestinal barrier function in Caco-2 cells following oxygen-glucose deprivation/re-oxygenation-induced injury. Exp. Ther. Med. 2022, 23, 66. [Google Scholar] [CrossRef]
- Fang, W.; Zhu, S.; Niu, Z.; Yin, Y. The protective effect of syringic acid on dextran sulfate sodium-induced experimental colitis in BALB/c mice. Drug Dev. Res. 2019, 80, 731–740. [Google Scholar] [CrossRef]
- Miene, C.; Weise, A.; Glei, M. Impact of polyphenol metabolites produced by colonic microbiota on expression of COX-2 and GSTT2 in human colon cells (LT97). Nutr. Cancer 2011, 63, 653–662. [Google Scholar] [CrossRef] [PubMed]
- Lofft, Z.; Taibi, A.; Massara, P.; Tokar, T.; Paetau-Robinson, I.; Khoo, C.; Comelli, E.M. Cranberry proanthocyanidin and its microbial metabolite 3,4-dihydroxyphylacetic acid, but not 3-(4-hydroxyphenyl)-proprionic acid, partially reverse pro-inflammatory microRNA responses I human intestinal epithelial cells. Mol. Nutr. Food Res. 2022, 66, 2100853. [Google Scholar] [CrossRef] [PubMed]
- Kure, A.; Nakagawa, K.; Kondo, M.; Kato, F.; Kimura, F.; Watanabe, A.; Shoji, N.; Hatanaka, S.; Tsushida, T.; Miyazawa, T. Metabolic fate of luteolin in rats: Its relationship to anti-inflammatory effect. J. Agric. Food Chem. 2016, 64, 4246–4254. [Google Scholar] [CrossRef]
- Ben-Othman, S.; Jõudu, I.; Bhat, R. Bioactives from agri-food wastes: Present insights and future challenges. Molecules 2020, 25, 510. [Google Scholar] [CrossRef] [Green Version]
- Luo, C.; Zou, L.; Sun, H.; Peng, J.; Gao, C.; Bao, L.; Ji, R.; Jin, Y.; Sun, S. A review of the anti-inflammatory effects of rosmarinic acid on inflammatory diseases. Front. Pharmacol. 2020, 11, 153. [Google Scholar] [CrossRef]
- Petersen, M.; Simmonds, M.S.J. Molecules of interest. Rosmarinic acid. Phytochemistry 2003, 62, 121–125. [Google Scholar] [CrossRef]
- Osakabe, N.; Yasuda, A.; Natsume, M.; Sanbongi, C.; Kato, Y.; Osawa, T.; Yoshikawa, T. Rosmarinic acid, a major polyphenolic component of Perilla frutescens, reduces lipopolysaccharide (LPS)-induced liver injury in d-galactosamine (d-GalN)-sensitized mice. Free Radic. Biol. Med. 2002, 33, 798–806. [Google Scholar] [CrossRef]
- Alagawany, M.; El-Hack, M.E.A.; Farag, M.R.; Gopi, M.; Karthik, K.; Malik, Y.S.; Dhama, K. Rosmarinic acid: Modes of action, medicinal values and health benefits. Anim. Health Res. Rev. 2017, 18, 167–176. [Google Scholar] [CrossRef]
- Nadeem, M.; Imran, M.; Gondal, T.A.; Imran, A.; Shahbaz, M.; Amir, R.M.; Sajid, M.W.; Qaisrani, T.B.; Atir, M.; Hussain, G.; et al. Therapeutic potential of rosmarinic acid: A comprehensive review. Appl. Sci. 2019, 9, 3139. [Google Scholar] [CrossRef] [Green Version]
- Alam, M.; Ahmed, S.; Elasbali, A.M.; Adnan, M.; Alam, S.; Hassan, M.I.; Pasupuleti, V.R. Therapeutic implications of caffeic acid in cancer and neurological diseases. Front. Oncol. 2022, 12, 860508. [Google Scholar] [CrossRef]
- Mirzaei, S.; Gholami, M.H.; Zabolian, G.; Saleki, H.; Farahani, M.V.; Hamzehlou, S.; Far, F.B.; Sharifzadeh, S.O.; Samarghandian, S.; Khan, H.; et al. Caffeic acid and its derivatives as potential modulators of oncogenic molecular pathways: New hope in the fight against cancer. Pharmacol. Res. 2021, 171, 105759. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Tang, L.; Yi, Q. Salvianolic acids: Potential source of natural drugs for the treatment of fibrosis disease and cancer. Front. Pharmacol. 2019, 10, 97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farooqi, A.A.; Butt, G.; El-Zahaby, S.; Attar, R.; Sabitaliyevich, U.Y.; Jovic, J.J.; Tang, K.-F.; Naureen, H.; Xu, B. Luteolin mediated targeting of protein network and microRNAs in different cancers: Focus on JAK-STAT, NOTCH, mTOR and TRAIL-mediated signaling pathways. Pharmacol. Res. 2020, 160, 105188. [Google Scholar] [CrossRef] [PubMed]
- Petersen, M. Rosmarinic acid: New aspects. Phytochem. Rev. 2013, 12, 207–227. [Google Scholar] [CrossRef]
- Bulgakov, V.P.; Inyushkina, Y.V.; Fedoreyev, S.A. Rosmarinic acid and its derivatives: Biotechnology and applications. Crit. Rev. Biotechnol. 2012, 32, 203–217. [Google Scholar] [CrossRef]
- Shetty, K. Biosynthesis and medical applications of rosmarinic acid. J. Herbs Spices Med. Plants 2001, 8, 161–181. [Google Scholar] [CrossRef]
- Chelly, M.; Chelly, S.; Occhiuto, C.; Cimino, F.; Cristani, M.; Saija, A.; Muscarà, C.; Ruberto, G.; Speciale, A.; Bouaziz-Ketata, H.; et al. Comparison of phytochemical profile and bioproperties of methanolic extracts from different parts of Tunisian Rumex roseus. Chem. Biodivers. 2021, 18, e2100185. [Google Scholar] [CrossRef]
- Ouerghemmi, S.; Sebei, H.; Siracusa, L.; Ruberto, G.; Saija, A.; Allouche, F.; Dhaouadi, K.; Cimino, F.; Cristani, M. LC-DAD-ESI-MS and HPLC-DAD phytochemical investigation and in vitro antioxidant assessment of Rosa sp. stem pruning products from different northern areas in Tunisia. Phytochem. Anal. 2020, 31, 98–111. [Google Scholar] [CrossRef]
- Abidi, J.; Occhiuto, C.; Cimino, F.; Speciale, A.; Ruberto, G.; Siracusa, L.; Bouaziz, M.; Boumendjel, B.; Muscarà, C.; Saija, A.; et al. Phytochemical and biological characterization of methanolic extracts from Rumex algeriensis and Rumex tunetanus. Chem. Biodivers. 2020, 17, e2000345. [Google Scholar] [CrossRef]
- Ferrari, D.; Speciale, A.; Cristani, M.; Fratantonio, D.; Molonia, M.S.; Ranaldi, G.; Saija, A.; Cimino, F. Cyanidin-3-O-glucoside inhibits NF-kB signalling in intestinal epithelial cells to TNF-α and exerts protective effects via Nrf2 pathway activation. Toxicol. Lett. 2016, 264, 51–58. [Google Scholar] [CrossRef]
- Cimino, F.; Speciale, A.; Anwar, S.; Canali, R.; Ricciardi, E.; Virgili, F.; Trombetta, D.; Saija, A. Anthocyanins protect human endothelial cells from mild hyperoxia damage through modulation of Nrf2 pathway. Genes Nutr. 2013, 8, 391–399. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantification of micrograms quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Box, G.E.P.; Cox, D.R. An analysis of transformations. J. R. Stat. Soc. Ser. B 1964, 26, 211–234. [Google Scholar] [CrossRef]
- Osborne, J.W. Improving your data transformations: Applying the Box-Cox transformation. Pract. Assess. Res. Eval. 2010, 15, 1–9. [Google Scholar]
- Gomez, K.A.; Gomez, A.A. Statistical Procedures for Agricultural Research; John Wiley & Sons: New York, NY, USA, 1984. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | Significance of F (1) | O. vulgare | T. vulgaris | S. officinalis | R. officinalis | O. majorana |
|---|---|---|---|---|---|---|
| Organic Acids Derivatives | *** | 94.08 ± 0.84 C | 118.37± 0.87 B | 123.38 ± 1.65 A | 92.35 ± 0.55 C | 88.22 ± 0.90 D |
| Dihydroxyphenylacetic acid | 20.39 | 2.79 | 6.28 | 17.31 | 4.46 | |
| Syringic acid | 5.47 | 1.94 | - | - | - | |
| Benzoic acid derivative | 1.36 | 1.45 | - | - | 2.49 | |
| Cinnamic acid derivative A | - | - | 1.42 | - | - | |
| Feroulic acid derivative | 3.68 | 6.04 | 3.88 | 4.63 | 2.23 | |
| Caffeic acid | - | 0.36 | - | - | 0.52 | |
| Caffeic acid trimer A | - | - | 1.60 | - | - | |
| Cinnamic acid derivative B | - | - | - | - | 7.57 | |
| Syringoyl dihexoside | 9.30 | - | - | - | - | |
| Salvianolic acid isomer A | 4.55 | - | - | - | - | |
| Rosmarinic acid glucoside | - | 3.05 | - | - | 1.51 | |
| Salvianolic acid isomer B | - | - | 3.43 | - | - | |
| Caffeic acid tetramer A | - | - | 13.85 | - | - | |
| Rosmarinic acid | 49.35 | 66.37 | 54.78 | 70.41 | 53.98 | |
| Salvianolic acid K | - | 18.43 | 35.92 | - | - | |
| Salvianolic acid isomer C | - | 2.83 | - | - | - | |
| Caffeic acid trimer B | - | 13.24 | 2.22 | - | - | |
| Caffeic acid tetramer B | - | - | - | - | 15.46 | |
| Cinnamic acid derivative C | - | 1.86 | - | - | - | |
| Flavonoids Derivatives | *** | 30.77 ± 0.33 C | 42.91 ± 2.51 B | 51.88 ± 0.64 A | 41.90 ± 0.07 B | 41.53 ± 0.24 B |
| Flavones (2) | *** | 30.77 ± 0.33 C | 27.63 ± 2.50 C | 42.74 ± 0.77 A | 28.16 ± 0.02 C | 39.93 ± 0.22 B |
| Apigenin 6-8-di-C-glucoside | - | 1.11 | 1.31 | - | 3.35 | |
| Luteolin derivative | 6.55 | - | - | - | - | |
| Nepitrin | - | - | 10.12 | - | - | |
| Apigenin derivative A | 2.84 | - | - | - | - | |
| Luteolin 7-O-rutinoside | - | - | 2.07 | - | - | |
| Luteolin 7-O-glucuronide | 8.97 | 26.24 | 28.49 | 3.28 | 26.07 | |
| Apigenin X-O-glucuronide | - | - | - | - | 10.51 | |
| Apigenin derivative B | 5.50 | - | - | - | - | |
| Apigenin 7-O-glucoside | 5.83 | - | - | - | - | |
| Luteolin 3-O-glucuronide | - | - | 0.75 | 15.64 | - | |
| Luteolin X-O-acetylglucuronide A | - | - | - | 3.35 | - | |
| Luteolin X-O-acetylglucuronide B | - | - | - | 3.41 | - | |
| Luteolin X-O-acetylglucuronide C | - | - | - | 2.47 | - | |
| Methyl apigenin X-O-glucuronide | 1.08 | - | - | - | - | |
| Flavonols | *** | 0.00 E | 15.28 ± 0.15 A | 9.14 ± 0.18 C | 13.74 ± 0.07 B | 1.60 ± 0.02 D |
| Quercetin 3-O-glucoside | - | 7.16 | - | - | - | |
| Quercetin derivative | - | 3.65 | - | - | - | |
| Quercetin 3-O-rhamnoside | - | 4.48 | 6.35 | - | 1.60 | |
| Quercetin 3-O-glucuronide | - | - | 2.79 | 13.74 | - | |
| Total | *** | 124.86 ± 1.17 E | 161.28 ± 2.81 B | 175.26 ± 1.46 A | 134.24 ± 0.60 C | 129.75 ± 1.12 D |
| DPPH Assay μmol TE/mg dm | ORAC Assay μmol TE/mg dm | SOD-Like Activity Assay SC50 mg dm/mL (CL 95%) | |
|---|---|---|---|
| Origanum vulgare | 1.63 ± 0.13 | 4.40 ± 0.17 b | 0.967 (0.726–1.287) |
| Thymus vulgaris | 1.79 ± 0.22 | 4.22 ± 0.06 b | 0.978 (0.738–1.295) |
| Salvia officinalis | 1.49 ± 0.08 | 4.70 ± 0.11 b | 0.991 (0.747–1.315) |
| Rosmarinus officinalis | 1.41 ± 0.15 | 3.35 ± 0.63 | 1.168 (0.856–1.592) |
| Origanum majorana | 1.05 ± 0.22 c | 3.63 ± 0.26 | 2.102 a (1.658–3.029) |
| PC | Eigenvalue | % Variance |
|---|---|---|
| 1 | 3.93527 | 49.191 |
| 2 | 2.34933 | 29.367 |
| 3 | 1.43137 | 17.892 |
| 4 | 0.20556 | 2.5695 |
| 5 | 0.05069 | 0.6326 |
| 6 | 0.02786 | 0.3482 |
| 7 | 8.75278 × 10−8 | 1.0941 × 10−6 |
| 8 | 6.86664 × 10−9 | 8.5833 × 10−8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Napoli, E.; Ruberto, G.; Carrubba, A.; Sarno, M.; Muscarà, C.; Speciale, A.; Cristani, M.; Cimino, F.; Saija, A. Phenolic Profiles, Antioxidant and Anti-Inflammatory Activities of Hydrodistillation Wastewaters from Five Lamiaceae Species. Molecules 2022, 27, 7427. https://doi.org/10.3390/molecules27217427
Napoli E, Ruberto G, Carrubba A, Sarno M, Muscarà C, Speciale A, Cristani M, Cimino F, Saija A. Phenolic Profiles, Antioxidant and Anti-Inflammatory Activities of Hydrodistillation Wastewaters from Five Lamiaceae Species. Molecules. 2022; 27(21):7427. https://doi.org/10.3390/molecules27217427
Chicago/Turabian StyleNapoli, Edoardo, Giuseppe Ruberto, Alessandra Carrubba, Mauro Sarno, Claudia Muscarà, Antonio Speciale, Mariateresa Cristani, Francesco Cimino, and Antonella Saija. 2022. "Phenolic Profiles, Antioxidant and Anti-Inflammatory Activities of Hydrodistillation Wastewaters from Five Lamiaceae Species" Molecules 27, no. 21: 7427. https://doi.org/10.3390/molecules27217427
APA StyleNapoli, E., Ruberto, G., Carrubba, A., Sarno, M., Muscarà, C., Speciale, A., Cristani, M., Cimino, F., & Saija, A. (2022). Phenolic Profiles, Antioxidant and Anti-Inflammatory Activities of Hydrodistillation Wastewaters from Five Lamiaceae Species. Molecules, 27(21), 7427. https://doi.org/10.3390/molecules27217427