


Effect of Structured Phenolic Lipids with EPA/DHA and Gallic Acid against Metabolic-Associated Fatty Liver Disease (MAFLD) in Mice

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Results
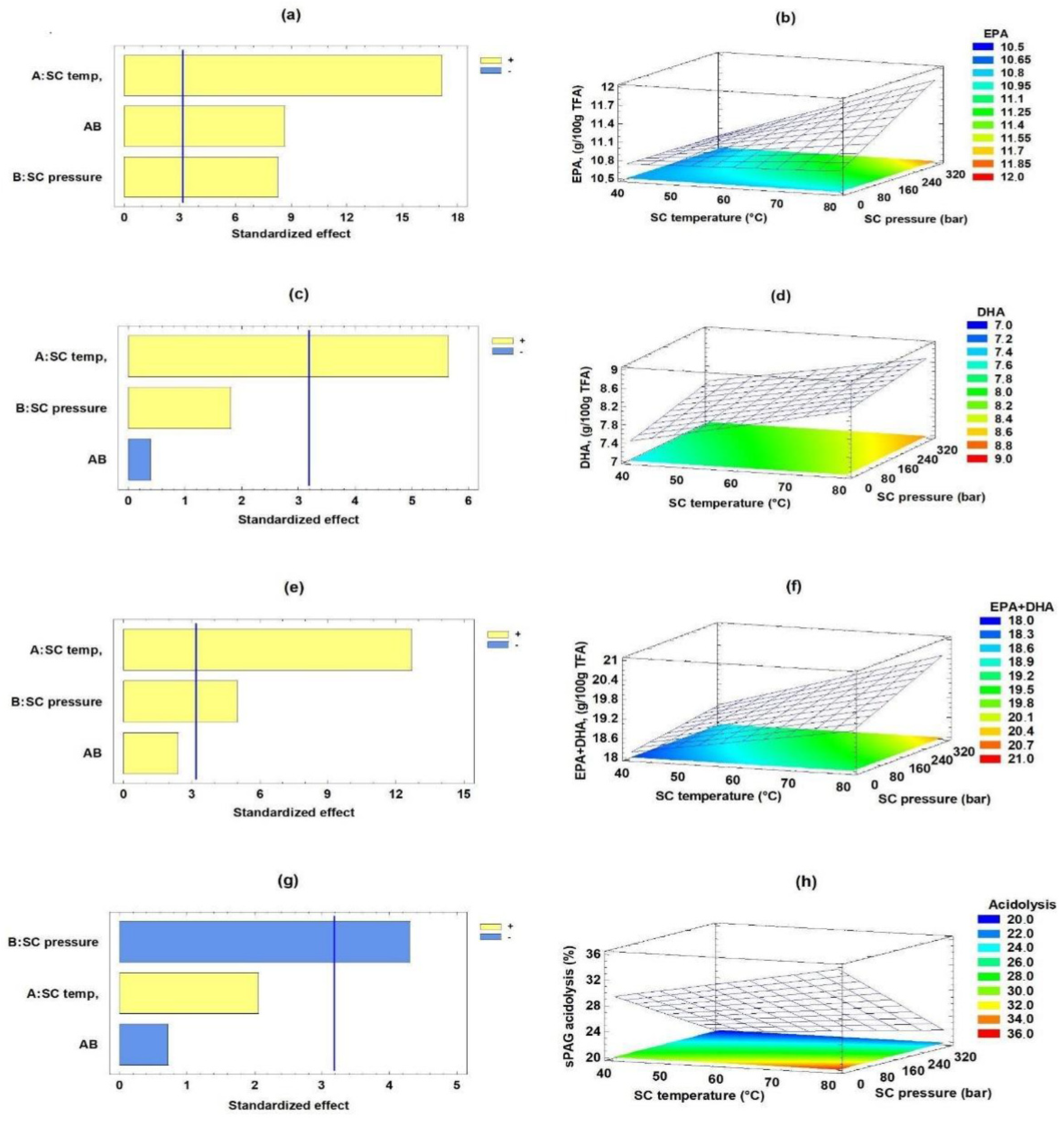
2.1. Acidolysis Process for Obtaining sPAG from sAG by 2²-Screening Factorial Design
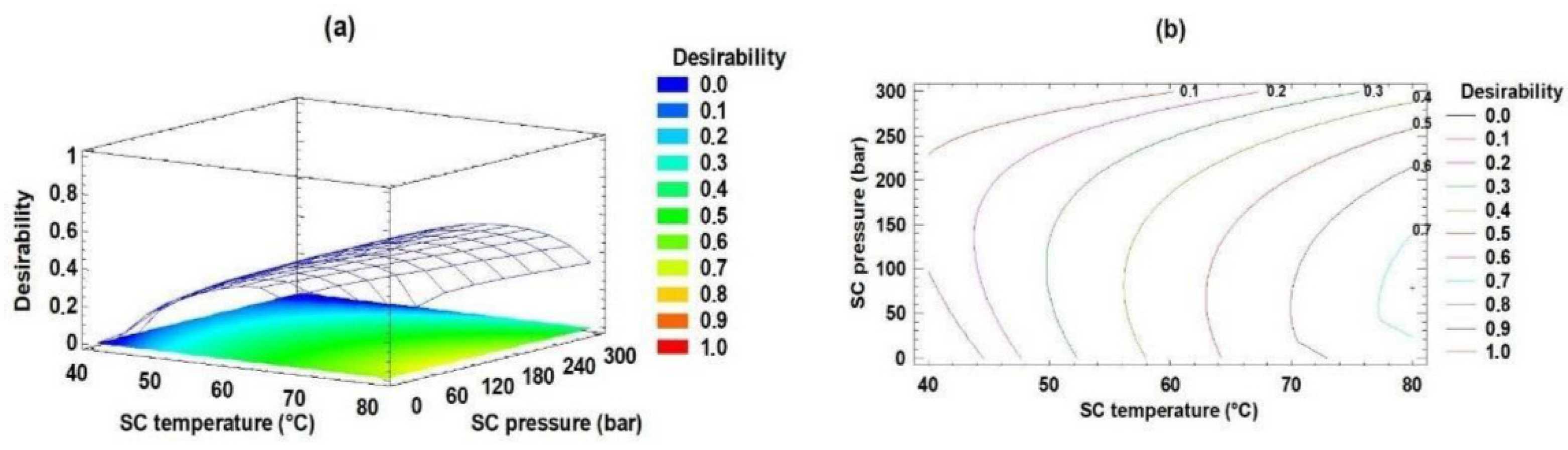
2.2. Optimization of the Acidolysis Process for Obtaining sPAG by RSM
2.3. Validation of the Acidolysis Process for Obtaining sPAG by RSM
2.4. Structural Analysis of Optimal Validated sPAG by HPLC-UV
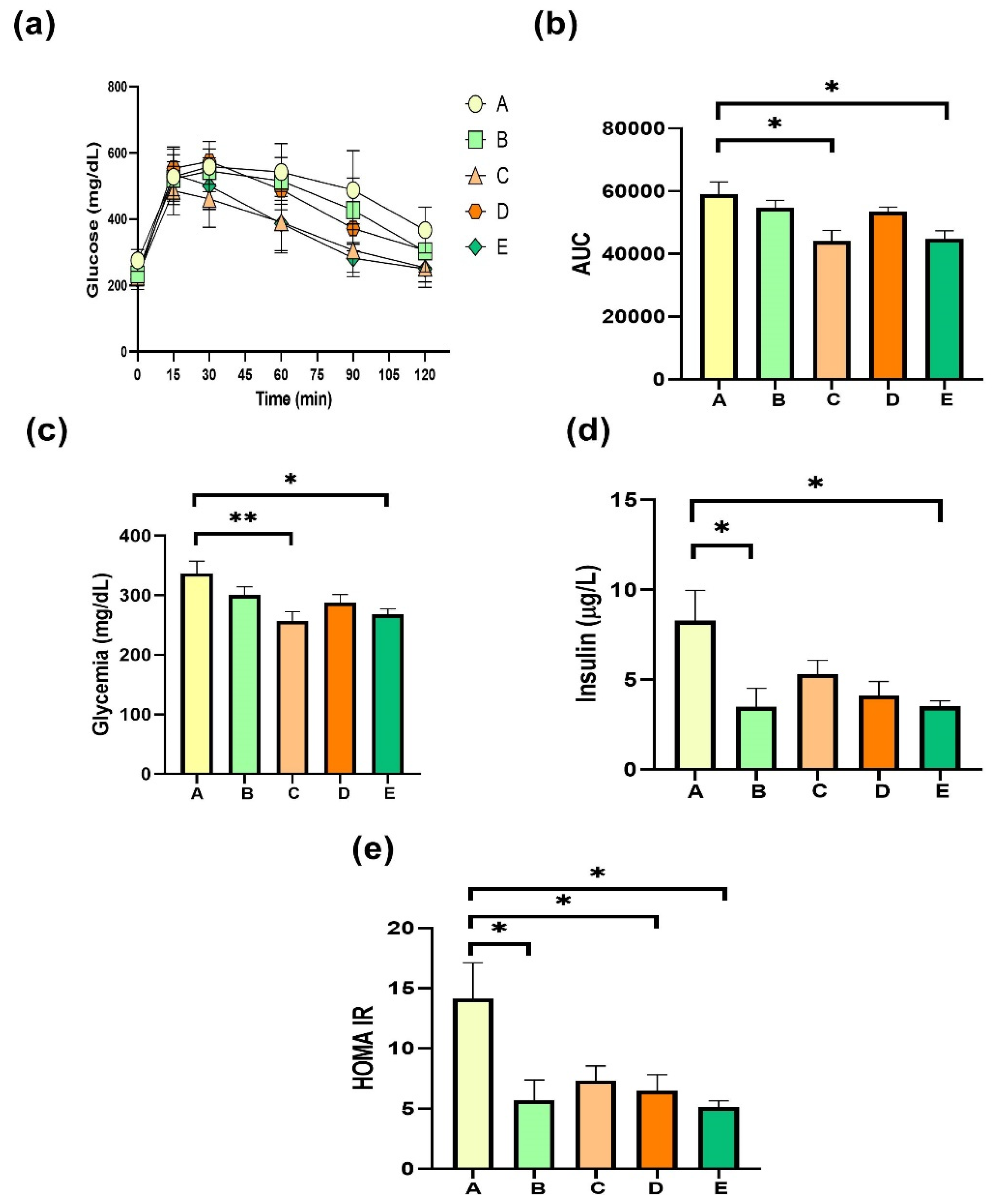
2.5. Glucose Homeostasis after the Treatment with sPAG
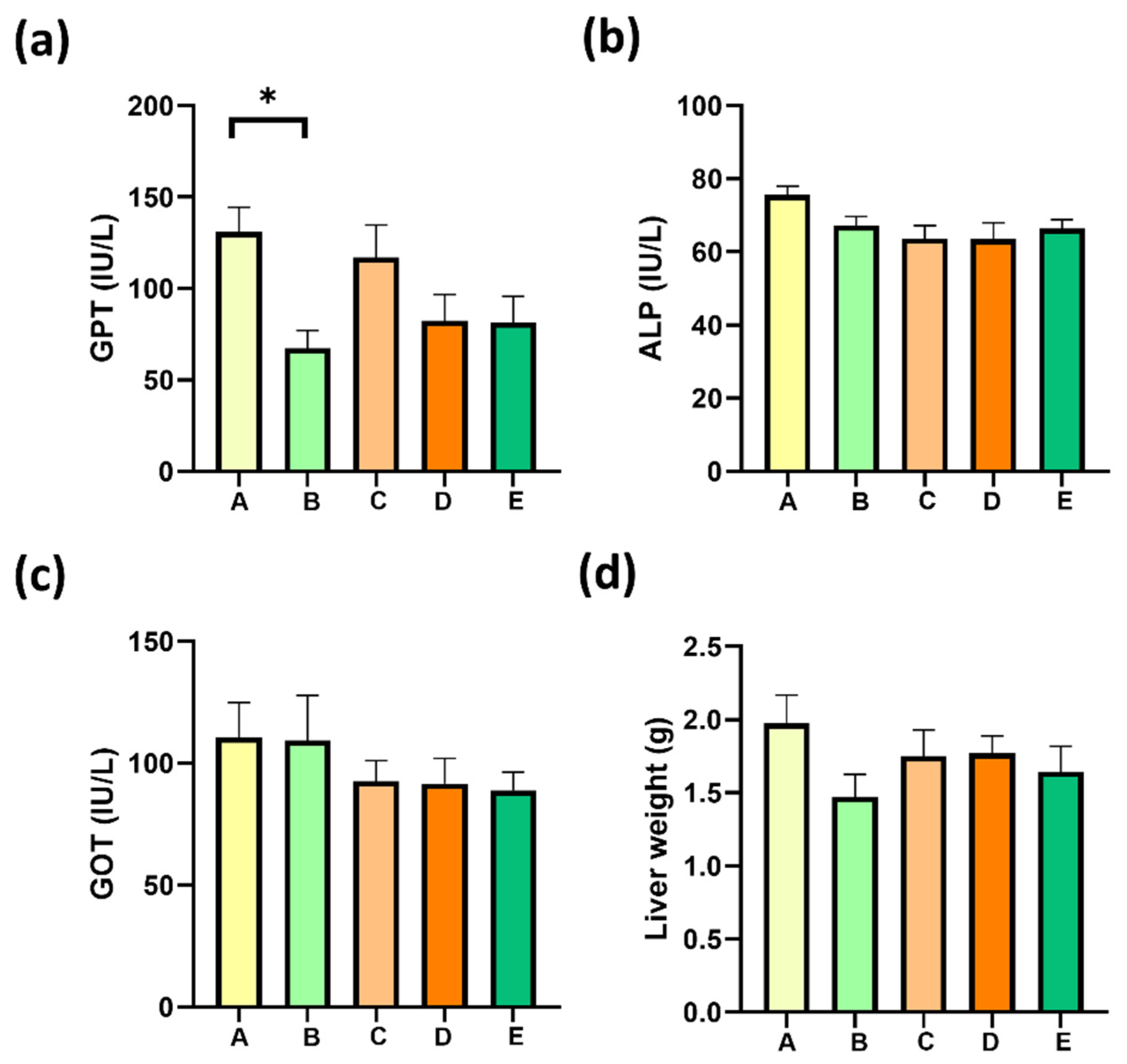
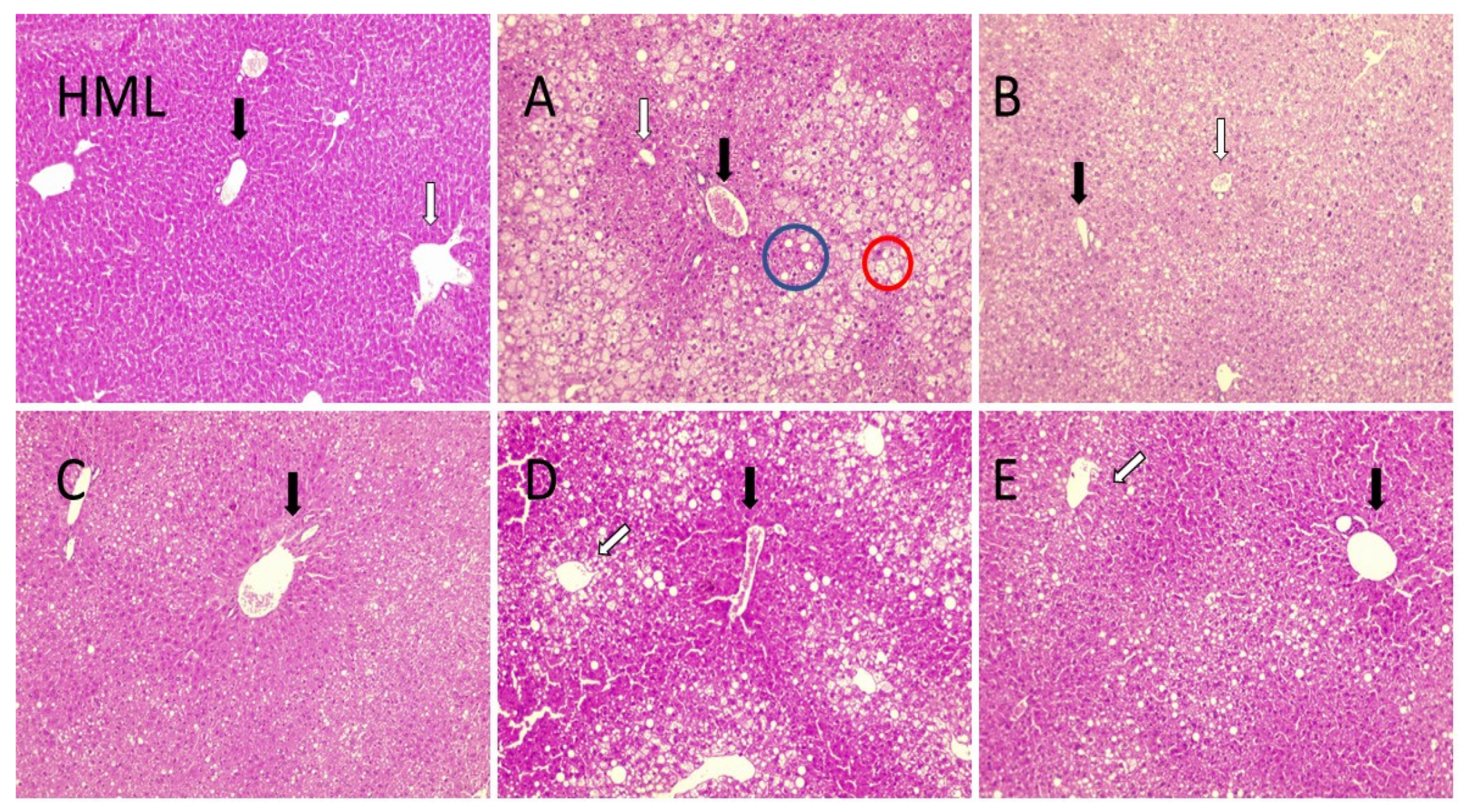
2.6. Gallic Acid Supplementation Decrease MAFLD-Associated Liver Damage
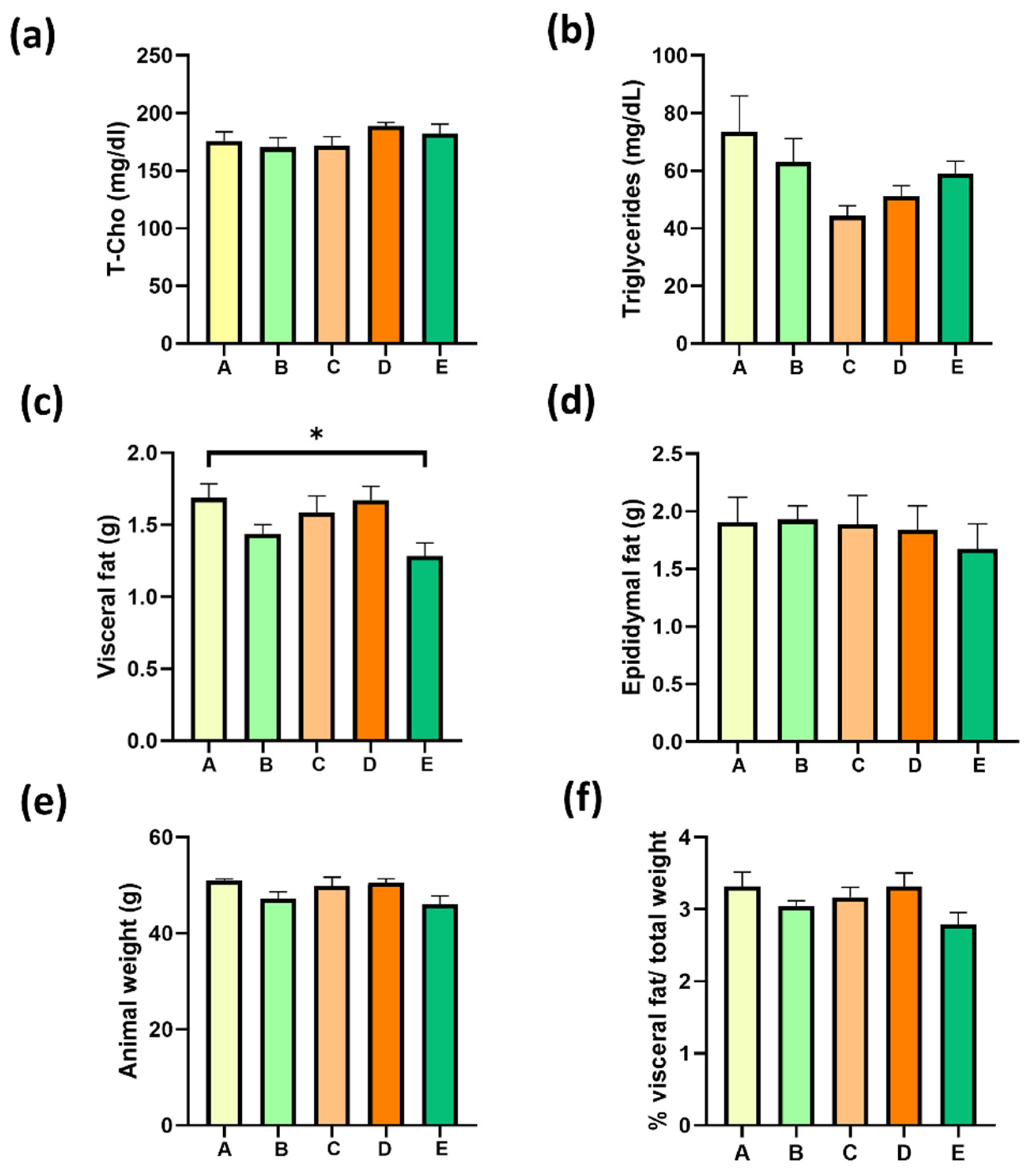
2.7. Four Weeks of Treatment with Structured Lipids Supplementation do Not Decrease MAFLD-Associated Fatty Tissue Weight nor Dyslipidemia
2.8. Four Weeks of Treatment with sPAG Supplementation Decrease Visceral Fat in Obese Mice
3. Discussion
4. Materials and Methods
4.1. Fatty Acid Composition and Quantification by GLC
4.2. Preparation of sPAG by Enzymatic Acidolysis Process under CO2SC Using RSM
4.3. Purification of sPAG by Neutralization with NaOH
4.4. Analysis of the Formation of sPAG with Gallic Acid by HPLC-UV
4.5. Optimization of sPAG Synthesis by RSM
4.6. Animals
4.7. Biological Samples
4.8. Measurements of Serum Parameters
4.9. Glucose Tolerance Test
4.10. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Eslam, M.; Newsome, P.N.; Sarin, S.K.; Anstee, Q.M.; Targher, G.; Romero-Gomez, M.; Zelber-Sagi, S.; Wong, V.W.-S.; Dufour, J.-F.; Schattenberg, J.M.; et al. A new definition for metabolic dysfunction-associated fatty liver disease: An international expert consensus statement. J. Hepatol. 2020, 73, 202–209. [Google Scholar] [CrossRef]
- Yuan, Q.; Wang, H.; Gao, P.; Chen, W.; Lv, M.; Bai, S.; Wu, J. Prevalence and risk factors of metabolic-associated fatty liver disease among 73,566 individuals in Beijing, China. Int. J. Environ. Res. Public Health 2022, 19, 2096. [Google Scholar] [CrossRef]
- Sakurai, Y.; Kubota, N.; Yamauchi, T.; Kadowaki, T. Role of insulin resistance in MAFLD. Int. J. Mol. Sci. 2021, 22, 4156. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, V.H.; Le, M.H.; Cheung, R.C.; Nguyen, M.H. Differential clinical characteristics and mortality outcomes in persons with NAFLD and/or MAFLD. Clin. Gastroenterol. Hepatol. 2021, 19, 2172–2181.e6. [Google Scholar] [CrossRef] [PubMed]
- Younossi, Z.; Anstee, Q.M.; Marietti, M.; Hardy, T.; Henry, L.; Eslam, M.; George, J.; Bugianesi, E. Global burden of NAFLD and NASH: Trends, predictions, risk factors and prevention. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 11–20. [Google Scholar] [CrossRef]
- Wang, C.; Liu, W.; Yao, L.; Zhang, X.; Zhang, X.; Ye, C.; Jiang, H.; He, J.; Zhu, Y.; Ai, D. Hydroxyeicosapentaenoic acids and epoxyeicosatetraenoic acids attenuate early occurrence of nonalcoholic fatty liver disease. Br. J. Pharmacol. 2017, 174, 2358–2372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antraco, V.J.; Hirata, B.; de Jesus Simão, J.; Cruz, M.M.; da Silva, V.S.; da Cunha de Sá, R.; Abdala, F.M.; Armelin-Correa, L.; Alonso-Vale, M. Omega-3 polyunsaturated fatty acids prevent nonalcoholic steatohepatitis (NASH) and stimulate adipogenesis. Nutrients 2021, 13, 622. [Google Scholar] [CrossRef]
- Soni, N.K.; Nookaew, I.; Sandberg, A.S.; Gabrielsson, B.G. Eicosapentaenoic and docosahexaenoic acid-enriched high fat diet delay the development of fatty liver in mice. Lipids Health Dis. 2015, 14, 74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schunck, W.H.; Konkel, A.; Fischer, R.; Weylandt, K.H. Therapeutic potential of omega-3 fatty acid-derived epoxy eicosanoids in cardiovascular and inflammatory diseases. Pharmacol. Ther. 2018, 183, 177–204. [Google Scholar] [CrossRef] [PubMed]
- Minihane, A.M.; Armah, C.K.; Miles, E.A.; Madden, J.M.; Clark, A.B.; Caslake, M.J.; Packard, C.J.; Kofler, B.M.; Lietz, G.; Curtis, P.J.; et al. Consumption of fish oil providing amounts of eicosapentaenoic acid and docosahexaenoic acid that can be obtained from the diet reduces blood pressure in adults with systolic hypertension: A retrospective analysis. J. Nutr. 2016, 146, 516–523. [Google Scholar] [CrossRef]
- Ofosu, F.K.; Daliri, E.B.M.; Lee, B.H.; Yu, X. Current Trends and Future Perspectives on Omega-3 Fatty Acids. Res. Rev. J. Biol. 2017, 5, 11–20, e-ISSN 2322-0066. [Google Scholar]
- Takahashi, M.; Ando, J.; Shimada, K.; Nishizaki, Y.; Tani, S.; Ogawa, T.; Yamamoto, M.; Nagao, K.; Hirayama, A.; Yoshimura, M.; et al. The ratio of serum n-3 to n-6 polyunsaturated fatty acids is associated with diabetes mellitus in patients with prior myocardial infarction: A multicenter cross-sectional study. BMC Cardiovasc. Disord. 2017, 17, 41. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations (FAO)-WHO-FINUT. Fat and Fatty Acids in Human Nutrition. Report of an Expert Consultation; Post republished; Food and Agriculture Organization of the United Nations (FAO), Ibero-American Nutrition Foundation (FINUT): Rome, Italy, 2010; pp. 1–170. ISSN 0254-4725. [Google Scholar]
- Valenzuela, A.; Sanhueza, J.; Nieto, S. El uso de lípidos estructurados en la nutrición: Una tecnología que abre nuevas perspectivas en el desarrollo de productos innovadores. Rev. Chilena Nutr. 2002, 29, 106–115. [Google Scholar] [CrossRef]
- Fanaei, H.; Mard, S.A.; Sarkaki, A.; Goudarzi, G.; Khorsandi, L. Gallic acid protects the liver against NAFLD induced by dust exposure and high-fat diet through inhibiting oxidative stress and repressing the inflammatory signaling pathways NF-kβ/TNF-α/IL-6 in Wistar rats. Avicenna J. Phytomed. 2021, 11, 527–540. [Google Scholar] [CrossRef]
- Chao, J.; Cheng, H.Y.; Chang, M.L.; Huang, S.S.; Liao, J.W.; Cheng, Y.C.; Peng, W.H.; Pao, L.H. Gallic acid ameliorated impaired lipid homeostasis in a mouse model of high-fat diet-and streptozotocin-induced NAFLD and diabetes through improvement of β-oxidation and ketogenesis. Front. Pharmacol. 2021, 11, 606759. [Google Scholar] [CrossRef]
- Tanaka, M.; Sato, A.; Kishimoto, Y.; Mabashi-Asazuma, H.; Kondo, K.; Iida, K. Gallic acid inhibits lipid accumulation via AMPK pathway and suppresses apoptosis and macrophage-mediated inflammation in hepatocytes. Nutrients 2020, 12, 1479. [Google Scholar] [CrossRef]
- Baladia, E.; Basulto, J.; Manera, M.; Martínez, R.; Calbet, D. Effect of green tea or green tea extract consumption on body weight and body composition; systematic review and meta-analysis. Nutr. Hosp. 2014, 29, 479–490. [Google Scholar] [CrossRef]
- López-Vélez, M.; Martínez-Martínez, F.; Del Valle-Ribes, C. The study of phenolic compounds as natural antioxidants in wine. Crit. Rev. Food Sci. Nutr. 2003, 43, 233–244. [Google Scholar] [CrossRef]
- Sroka, Z.; Cisowski, W. Hydrogen peroxide scavenging, antioxidant and anti-radical activity of some phenolic acids. Food Chem. Toxicol. 2003, 41, 753–758. [Google Scholar] [CrossRef]
- Kroes, B.H.; van den Berg, A.J.; Quarles van Ufford, H.C.; van Dijk, H.; Labadie, R.P. Anti-inflammatory activity of gallic acid. Planta Med. 1992, 58, 499–504. [Google Scholar] [CrossRef] [PubMed]
- Kubo, I.; Xiao, P.; Fujita, K. Antifungal activity of octyl gallate: Structural criteria and mode of action. Bioorg. Med. Chem. Lett. 2001, 11, 347–350. [Google Scholar] [CrossRef]
- Wang, Y.C.; Bachrach, U. The specific anti-cancer activity of green tea (-)-epigallocatechin-3-gallate (EGCG). Amino Acids. 2002, 22, 131–143. [Google Scholar] [CrossRef] [PubMed]
- Lambert, J.D.; Yang, C.S. Cancer chemopreventive activity and bioavailability of tea and tea polyphenols. Mutat. Res. 2003, 523–524, 201–208. [Google Scholar] [CrossRef]
- Roidoung, S.; Dolan, K.D.; Siddiq, M. Gallic acid as a protective antioxidant against anthocyanin degradation and color loss in vitamin-C fortified cranberry juice. Food Chem. 2016, 210, 422–427. [Google Scholar] [CrossRef] [PubMed]
- Cuvelier, M.E.; Richard, H.; Berset, C. Comparison of the antioxidative activity of some acid-phenols: Structure-activity relationship. Biosci. Biotechnol. Biochem. 1992, 56, 324–325. [Google Scholar] [CrossRef] [Green Version]
- Moghadas, M.; Edalatmanesh, M.A.; Robati, R. Histopathological analysis from gallic acid administration on hippocampal cell density, depression, and anxiety related behaviors in a trimethyltin intoxication model. Cell J. 2016, 17, 659–667. [Google Scholar] [CrossRef]
- Huang, D.W.; Chang, W.C.; Wu, J.S.; Shih, R.W.; Shen, S.C. Gallic acid ameliorates hyperglycemia and improves hepatic carbohydrate metabolism in rats fed a high-fructose diet. Nutr. Res. 2016, 36, 150–160. [Google Scholar] [CrossRef]
- Chao, J.; Huo, T.I.; Cheng, H.Y.; Tsai, J.C.; Liao, J.W.; Lee, M.S.; Qin, X.M.; Hsieh, M.T.; Pao, L.H.; Peng, W.H. Gallic acid ameliorated impaired glucose and lipid homeostasis in high fat diet-induced NAFLD mice. PLoS ONE 2014, 9, e96969. [Google Scholar] [CrossRef] [Green Version]
- Jasińska, K.; Fabiszewska, A.; Białecka-Florjańczyk, E.; Zieniuk, B. Mini-review on the enzymatic lipophilization of phenolics present in plant extracts with the special emphasis on anthocyanins. Antioxidants 2022, 11, 1528. [Google Scholar] [CrossRef]
- Soto-Hernández, M.; Palma-Tenango, M.; García-Mateos, M.d.R. (Eds.) Phenolic Compounds—Natural Sources, Importance and Applications; IntechOpen: London, UK, 2017; p. 456. [Google Scholar] [CrossRef]
- Karboune, S.; St-Louis, R.; Kermasha, S. Enzymatic synthesis of structured phenolic lipids by acidolysis of flaxseed oil with selected phenolic acids. J. Mol. Catal. B Enzym. 2008, 52, 96–105. [Google Scholar] [CrossRef]
- Ciftci, D.; Saldaña, M.D.A. Enzymatic synthesis of phenolic lipids using flaxseed oil and ferulic acid carbon in supercritical dioxide media. J. Supercrit. Fluids. 2012, 72, 255–262. [Google Scholar] [CrossRef]
- Laszlo, J.A.; Compton, D.L. Enzymatic glycerolysis and transesterification of vegetable oil for enhanced production of feruloylated glycerols. J. Am. Oil Chem. Soc. 2006, 83, 765–770. [Google Scholar] [CrossRef]
- Iwasaki, Y.; Jun Han, J.J.; Narita, M.; Rosu, R.; Yamane, T. Enzymatic synthesis of structured lipids from single cell oil of high docosahexaenoic acid content. J. Am. Oil Chem. Soc. 1999, 76, 563–569. [Google Scholar] [CrossRef]
- Pando, M.E.; Rodríguez, A.; Valenzuela, M.A.; Berríos, M.; Rivera, M.; Romero, N. Acylglycerol synthesis including EPA and DHA from rainbow trout (Oncorhynchus mykiss) belly flap oil and caprylic acid catalyzed by Thermomyces lanuginosus lipase under supercritical carbon dioxide. Eur. Food Res. Technol. 2021, 247, 499–511. [Google Scholar] [CrossRef]
- Dovale-Rosabal, G.; Rodríguez, A.; Espinosa, A.; Barriga, A.; Aubourg, S.P. Synthesis of EPA- and DHA-enriched structured acylglycerols at the sn-2 position starting from commercial salmon oil by enzymatic lipase catalysis under supercritical conditions. Molecules 2021, 26, 3094. [Google Scholar] [CrossRef]
- Guo, Y.; Cai, Z.; Xie, Y.; Ma, A.; Zhang, H.; Rao, P.; Wang, Q. Synthesis, physicochemical properties, and health aspects of structured lipids: A review. Compr. Rev. Food Sci. Food Saf. 2020, 19, 759–800. [Google Scholar] [CrossRef]
- Kim, B.H.; Akoh, C.C. Recent research trends on the enzymatic synthesis of structured lipids. J. Food Sci. 2015, 80, C1713–C1724. [Google Scholar] [CrossRef]
- Akoh, C.; Pande, G. Structured lipids and health. In Bailey’s Industrial Oil and Fat Products; John Wiley & Sons: Hoboken, NJ, USA, 2020; pp. 1–17. [Google Scholar]
- Gunnlaugsdottir, H.; Järemo, M.; Sivik, B. Process parameters influencing ethanolysis of cod liver oil in supercritical carbon dioxide. J. Supercrit. Fluids 1998, 12, 85–93. [Google Scholar] [CrossRef]
- Gunstone, F.D. Enzymes as biocatalysts in the modification of natural lipids. J. Sci. Food Agric. 1999, 79, 1535–1549. [Google Scholar] [CrossRef]
- Budisa, N.; Schulze-Makuch, D. Supercritical carbon dioxide and its potential as a life-sustaining solvent in a planetary environment. Life 2014, 4, 331–340. [Google Scholar] [CrossRef]
- Sabally, K.; Karboune, S.; St-Louis, R.; Kermasha, S. Lipase-catalyzed transesterification of dihydrocaffeic acid with flaxseed oil for the synthesis of phenolic lipids. J. Biotechnol. 2006, 127, 167–176. [Google Scholar] [CrossRef]
- Haq, M.; Park, S.-K.; Kim, M.-J.; Cho, Y.-J.; Chun, B.-S. Modifications of Atlantic Salmon By-Product Oil for Obtaining Different ω-3 Polyunsaturated Fatty Acids Concentrates: An Approach to Comparative Analysis. J. Food Drug Anal. 2018, 26, 545–556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sabally, K.; Karboune, S.; Yeboah, F.K.; Kermasha, S. Lipase-Catalyzed Esterification of Selected Phenolic Acids with Linolenyl Alcohols in Organic Solvent Media. Appl. Biochem. Biotechnol. 2005, 127, 017–028. [Google Scholar] [CrossRef]
- Sabally, K.; Karboune, S.; St-Louis, R.; Kermasha, S. Lipase-Catalyzed Transesterification of Trilinolein or Trilinolenin with Selected Phenolic Acids. J. Am. Oil Chem. Soc. 2006, 83, 101–107. [Google Scholar] [CrossRef]
- Nocetti, D.; Espinosa, A.; Pino-De la Fuente, F.; Sacristán, C.; Bucarey, J.L.; Ruiz, P.; Valenzuela, R.; Chouinard-Watkins, R.; Pepper, I.; Troncoso, R.; et al. Lipid Droplets Are Both Highly Oxidized and Plin2-Covered in Hepatocytes of Diet-Induced Obese Mice. Appl. Physiol. Nutr. Metab. 2020, 45, 1368–1376. [Google Scholar] [CrossRef]
- Ortiz, M.; Soto-Alarcón, S.A.; Orellana, P.; Espinosa, A.; Campos, C.; López-Arana, S.; Rincón, M.A.; Illesca, P.; Valenzuela, R.; Videla, L.A. Suppression of High-Fat Diet-Induced Obesity-Associated Liver Mitochondrial Dysfunction by Docosahexaenoic Acid and Hydroxytyrosol Co-Administration. Dig. Liver Dis. 2020, 52, 895–904. [Google Scholar] [CrossRef]
- Sousa, J.N.; Paraíso, A.F.; Andrade, J.M.O.; Lelis, D.F.; Santos, E.M.; Lima, J.P.; Monteiro-Junior, R.S.; D’Angelo, M.F.S.V.; de Paula, A.M.B.; Guimarães, A.L.S.; et al. Oral Gallic Acid Improve Liver Steatosis and Metabolism Modulating Hepatic Lipogenic Markers in Obese Mice. Exp. Gerontol. 2020, 134, 110881. [Google Scholar] [CrossRef] [PubMed]
- Espinosa, A.; Ross, A.; Dovale-Rosabal, G.; Pino-de la Fuente, F.; Uribe-Oporto, E.; Sacristán, C.; Ruiz, P.; Valenzuela, R.; Romero, N.; Aubourg, S.P.; et al. EPA/DHA Concentrate by Urea Complexation Decreases Hyperinsulinemia and Increases Plin5 in the Liver of Mice Fed a High-Fat Diet. Molecules 2020, 25, 3289. [Google Scholar] [CrossRef] [PubMed]
- Illesca, P.; Valenzuela, R.; Espinosa, A.; Echeverría, F.; Soto-Alarcon, S.; Campos, C.; Rodriguez, A.; Vargas, R.; Magrone, T.; Videla, L.A. Protective Effects of Eicosapentaenoic Acid Plus Hydroxytyrosol Supplementation Against White Adipose Tissue Abnormalities in Mice Fed a High-Fat Diet. Molecules 2020, 25, 4433. [Google Scholar] [CrossRef]
- Echeverría, F.; Valenzuela, R.; Bustamante, A.; Álvarez, D.; Ortiz, M.; Espinosa, A.; Illesca, P.; Gonzalez-Mañan, D.; Videla, L.A. High-Fat Diet Induces Mouse Liver Steatosis with a Concomitant Decline in Energy Metabolism: Attenuation by Eicosapentaenoic Acid (EPA) or Hydroxytyrosol (HT) Supplementation and the Additive Effects upon EPA and HT Co-Administration. Food Funct. 2019, 10, 6170–6183. [Google Scholar] [CrossRef]
- Rahmadi, M.; Nurhan, A.D.; Pratiwi, E.D.; Prameswari, D.A.; Panggono, S.M.; Nisak, K.; Khotib, J. The Effect of Various High-Fat Diet on Liver Histology in the Development of NAFLD Models in Mice. J. Basic Clin. Physiol. Pharmacol. 2021, 32, 547–553. [Google Scholar] [CrossRef] [PubMed]
- Reckziegel, P.; Dias, V.T.; Benvegnú, D.M.; Boufleur, N.; Barcelos, R.C.S.; Segat, H.J.; Pase, C.S.; dos Santos, C.M.M.; Flores, É.M.M.; Bürger, M.E. Antioxidant Protection of Gallic Acid against Toxicity Induced by Pb in Blood, Liver and Kidney of Rats. Toxicol. Rep. 2016, 3, 351–356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cerdán, L.E.; Medina, A.R.; Giménez, A.G.; González, M.J.I.; Grima, E.M. Synthesis of Polyunsaturated Fatty Acid-Enriched Triglycerides by Lipase-Catalyzed Esterification. J. Am. Oil Chem. Soc. 1998, 75, 1329–1337. [Google Scholar] [CrossRef]
- Nunes, P.A.; Pires-Cabral, P.; Guillén, M.; Valero, F.; Luna, D.; Ferreira-Dias, S. Production of MLM-Type Structured Lipids Catalyzed by Immobilized Heterologous Rhizopus oryzae Lipase. J. Am. Oil Chem. Soc. 2011, 88, 473–480. [Google Scholar] [CrossRef]
- European Pharmacopoeia. Composition of Fatty Acids in Oils Rich in Omega-3-Acid, 5th ed.; 01/2005:20429; EDQM, Council of Europe: Strasbourg, France, 2005. [Google Scholar]
- AOCS. Determination of cis-, trans-, saturated, monounsaturated, and polyunsaturated fatty acids by capillary gas liquid chromatography (GLC). Sampling and analysis of commercial fats and oils. In Official Method Ce 1j-7. Official Methods and Recommended Practices of the American Oil Chemists Society (edited by AOCS); AOCS: Champaign, IL, USA, 2009. [Google Scholar]
- Jiménez, M.J.; Esteban, L.; Robles, A.; Hita, E.; González, P.A.; Muñío, M.M.; Molina, E. Production of Triacylglycerols Rich in Palmitic Acid at Position 2 as Intermediates for the Synthesis of Human Milk Fat Substitutes by Enzymatic Acidolysis. Process Biochem. 2010, 45, 407–414. [Google Scholar] [CrossRef]
- Myers, R.H.; Montgomery, D.C. Response Surface Methodology. A.E. Wiley-Interscience Publication; John Wiley & Sons, Inc.: New York, NY, USA, 1995. [Google Scholar]
- Wanasundara, U.N.; Shahidi, F. Positional distribution of fatty acids in triacylglycerols of seal blubber oil. J. Food Lipids 1997, 4, 51–64. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Independent Variables | Response Variables | |||||
|---|---|---|---|---|---|---|
| Run | Supercritical Temperature (°C) | Supercritical Pressure (Bar) | EPA (g/100 g TFA) | DHA (g/100 g TFA) | EPA + DHA (g/100 g TFA) | Synthesis of sPAG (% Acidolysis) |
| 1 | 40 | 78 | 10.745 | 7.504 | 18.249 | 28.1 |
| 2 | 80 | 78 | 11.089 | 8.421 | 19.510 | 33.3 |
| 3 | 40 | 300 | 10.730 | 7.837 | 18.567 | 21.4 |
| 4 | 80 | 300 | 11.779 | 8.636 | 20.415 | 23.9 |
| 5 | 60 | 189 | 11.043 | 8.337 | 19.381 | 24.2 |
| 6 | 60 | 189 | 11.016 | 8.223 | 19.239 | 23.8 |
| 7 | 60 | 189 | 11.088 | 8.009 | 19.097 | 24.4 |
| Systematic Name | Abbreviated Name | sAG | sPAG |
|---|---|---|---|
| Lauric acid | C12:0 | N/I | N/I |
| Myristic acid | C14:0 | 2.15 ± 0.00 | 1.66 ± 0.00 |
| Palmitic acid | C16:0 | 10.60 ± 0.00 | 7.90 ± 0.01 |
| Palmitoelaidic acid | C16:1 9t | 0.21 ± 0.00 | 0.22 ± 0.00 |
| Palmitoleic acid | C16:1 9c | 3.39 ± 0.00 | 3.21 ± 0.00 |
| Heptadecanoic acid | C17:0 | N/I | N/I |
| Heptadecenoic acid | C 17:1 10c | 0.44 ± 0.00 | 0.57 ± 0.00 |
| Stearic acid | C18:0 | 3.17 ± 0.00 | 2.54 ± 0.00 |
| Trans-vaccenic acid | C 18:1 11t | 0.47 ± 0.01 | 0.61 ± 0.00 |
| Oleic acid | C18:1 9c | 34.66 ± 0.03 | 27.60 ± 0.03 |
| Cis-Vaccenic acid | C18:1 7c | 2.74 ± 0.00 | 2.22 ± 0.00 |
| Linoleaidic acid | C18:2 9t 12t | 0.75 ± 0.01 | 0.94 ± 0.01 |
| Linoleic acid | C18:2 9c 12c | 15.48 ± 0.00 | 18.12 ± 0.00 |
| Gamma linolenic acid | C18:3 6c 9c 12c | 0.37 ± 0.00 | 0.44 ± 0.00 |
| 5-Eicosanoic acid | C 20:1 5c | N/I | 0.44 ± 0.00 |
| 8-Eicosanoic acid | C 20:1 8c | N/I | N/I |
| 11-Eicosanoic acid | C 20:1 11c | 1.94 ± 0.00 | 1.57 ± 0.00 |
| α-Linolenic acid | C 18:3 9c 12c 15c | 4.78 ± 0.00 | 5.95 ± 0.01 |
| Eicosadienoic acid | C 20:2 11c 14c | 0.90 ± 0.00 | 0.78 ± 0.00 |
| Behenoic acid | C 22:0 | N/I | N/I |
| Eicosatrienoic acid | C 20:3 11c 14c 17c | 1.29 ± 0.00 | 1.08 ± 0.01 |
| Erucic acid | C 22:1 13c | N/I | N/I |
| Arachidonic acid | C 20:4 5c 8c 11c 14c | N/I | N/I |
| Docosadienoic acid | C 22:2 13c 16c | 1.07 ± 0.00 | 1.41 ± 0.00 |
| Eicosapentaenoic acid | C 20:5 5c 8c 11c 14c 17c | 5.92 ± 0.01 | 11.11 ± 0.03 |
| Nervonic acid | C 24:1 9c | N/I | N/I |
| Docosatetraenoic acid | C 22:4 7c 10c 13c 16c | 0.49 ± 0.00 | 0.67 ± 0.00 |
| Docosapentaenoic acid | C 22:5 7c, 10c 13c 16c 19c | 2.00 ± 0.00 | 2.52 ± 0.00 |
| Docosahexaenoic acid | C 22:6 4c 7c 10c 13c 16c 19c | 7.18 ± 0.02 | 8.44 ± 0.00 |
| Total saturated fatty acids (TSFA) | 15.92 | 12.10 | |
| Total monounsaturated fatty acids (TMUFA) | 43.85 | 36.44 | |
| Total polyunsaturated fatty acids (TPUFA) | 40.23 | 51.46 | |
| Total n-3 long-chain PUFA (n-3 TLCPUFA) | 16.39 | 23.15 | |
| Total n-3 fatty acids (n-3 TFA) | 21.17 | 29.10 | |
| EPA + DHA | 13.10 | 19.55 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dovale-Rosabal, G.; Espinosa, A.; Rodríguez, A.; Barriga, A.; Palomino-Calderón, A.; Romero, N.; Troncoso, R.H.; Aubourg, S.P. Effect of Structured Phenolic Lipids with EPA/DHA and Gallic Acid against Metabolic-Associated Fatty Liver Disease (MAFLD) in Mice. Molecules 2022, 27, 7702. https://doi.org/10.3390/molecules27227702
Dovale-Rosabal G, Espinosa A, Rodríguez A, Barriga A, Palomino-Calderón A, Romero N, Troncoso RH, Aubourg SP. Effect of Structured Phenolic Lipids with EPA/DHA and Gallic Acid against Metabolic-Associated Fatty Liver Disease (MAFLD) in Mice. Molecules. 2022; 27(22):7702. https://doi.org/10.3390/molecules27227702
Chicago/Turabian StyleDovale-Rosabal, Gretel, Alejandra Espinosa, Alicia Rodríguez, Andrés Barriga, Alan Palomino-Calderón, Nalda Romero, Rodrigo Hernán Troncoso, and Santiago Pedro Aubourg. 2022. "Effect of Structured Phenolic Lipids with EPA/DHA and Gallic Acid against Metabolic-Associated Fatty Liver Disease (MAFLD) in Mice" Molecules 27, no. 22: 7702. https://doi.org/10.3390/molecules27227702
APA StyleDovale-Rosabal, G., Espinosa, A., Rodríguez, A., Barriga, A., Palomino-Calderón, A., Romero, N., Troncoso, R. H., & Aubourg, S. P. (2022). Effect of Structured Phenolic Lipids with EPA/DHA and Gallic Acid against Metabolic-Associated Fatty Liver Disease (MAFLD) in Mice. Molecules, 27(22), 7702. https://doi.org/10.3390/molecules27227702