
Chemical Composition, Phytotoxic and Antibiofilm Activity of Seven Eucalyptus Species from Tunisia


 , , ,
, , ,  ,
,  and
and
Abstract
:1. Introduction
2. Results and Discussion
2.1. Chemical Composition
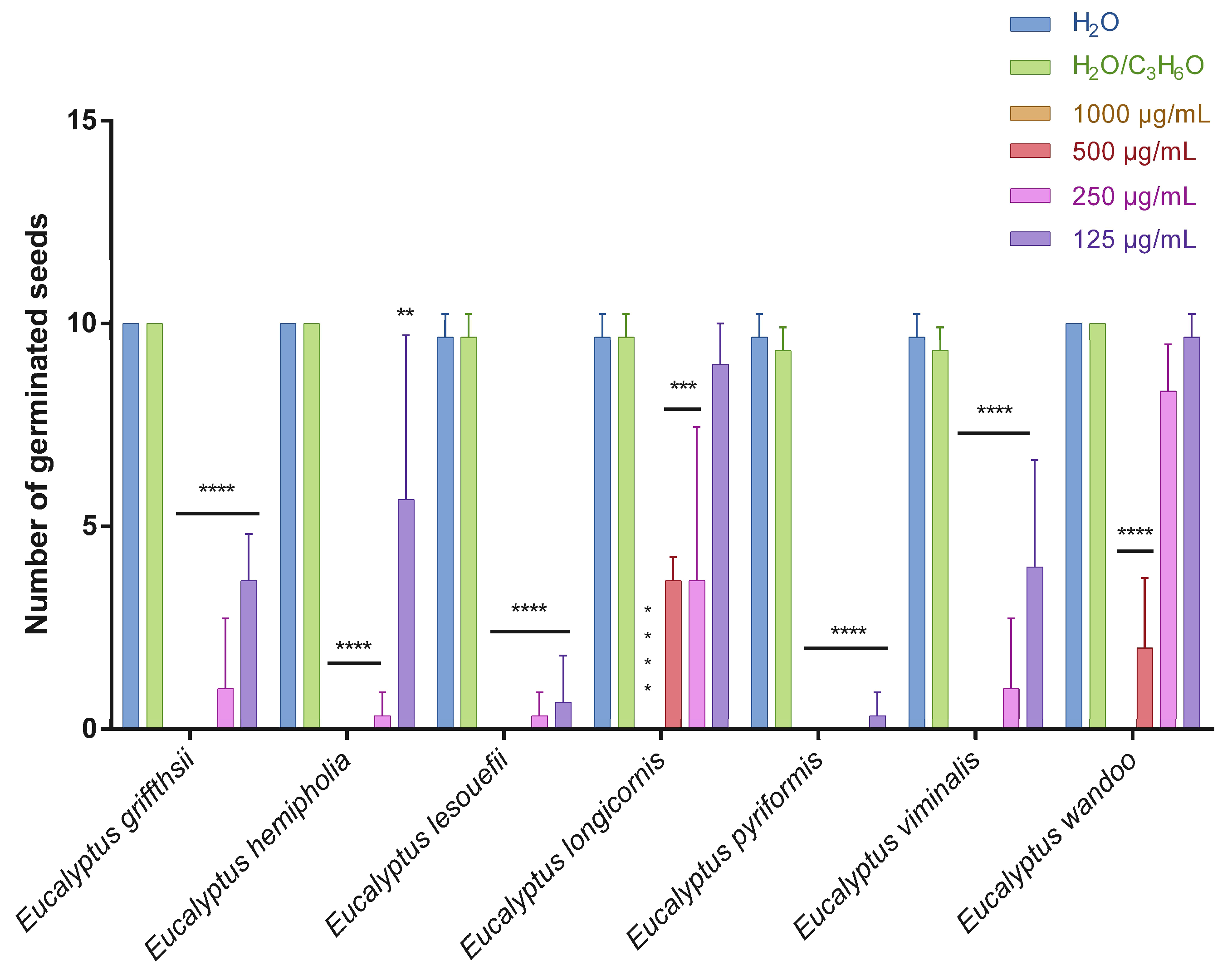
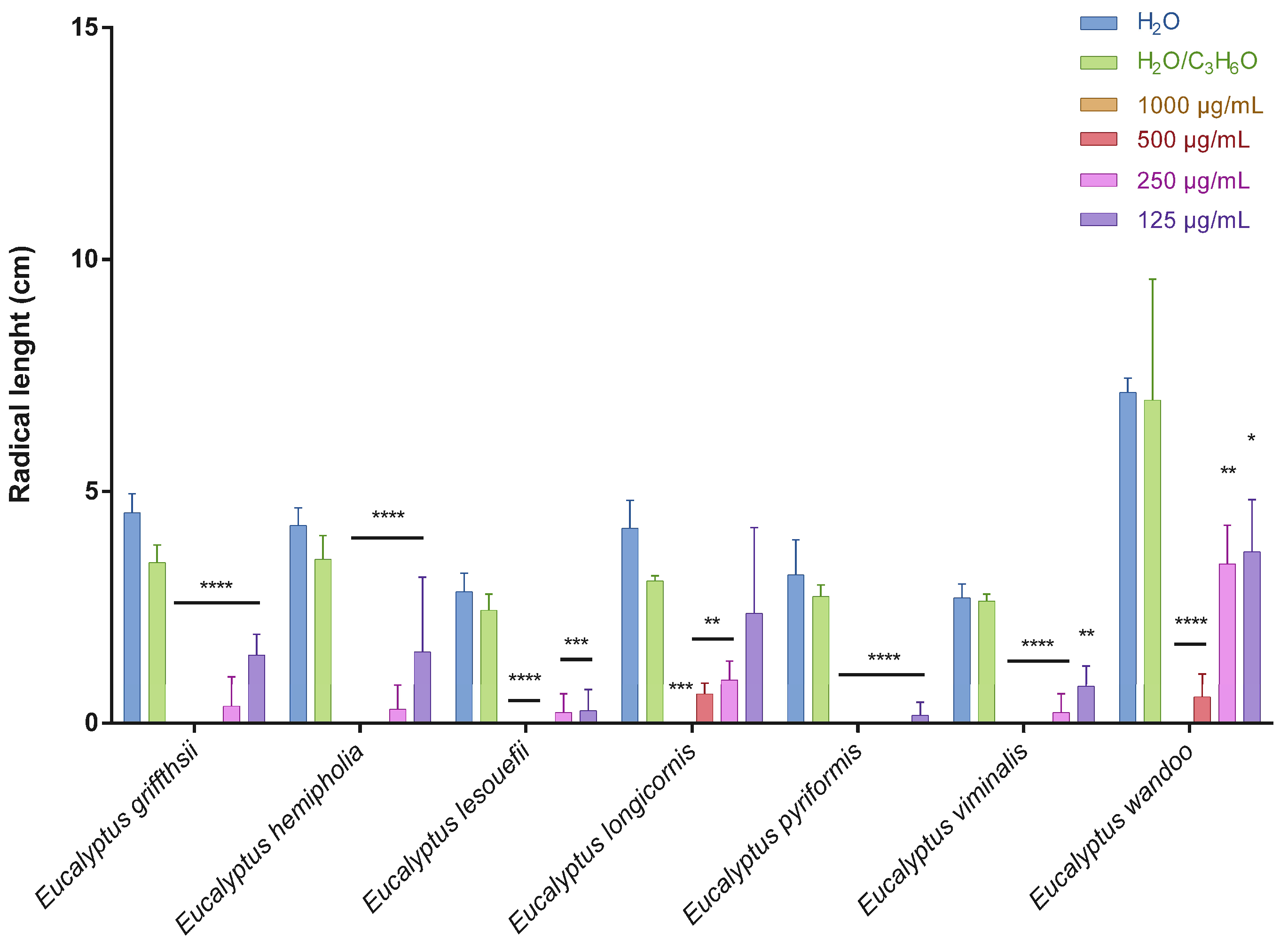
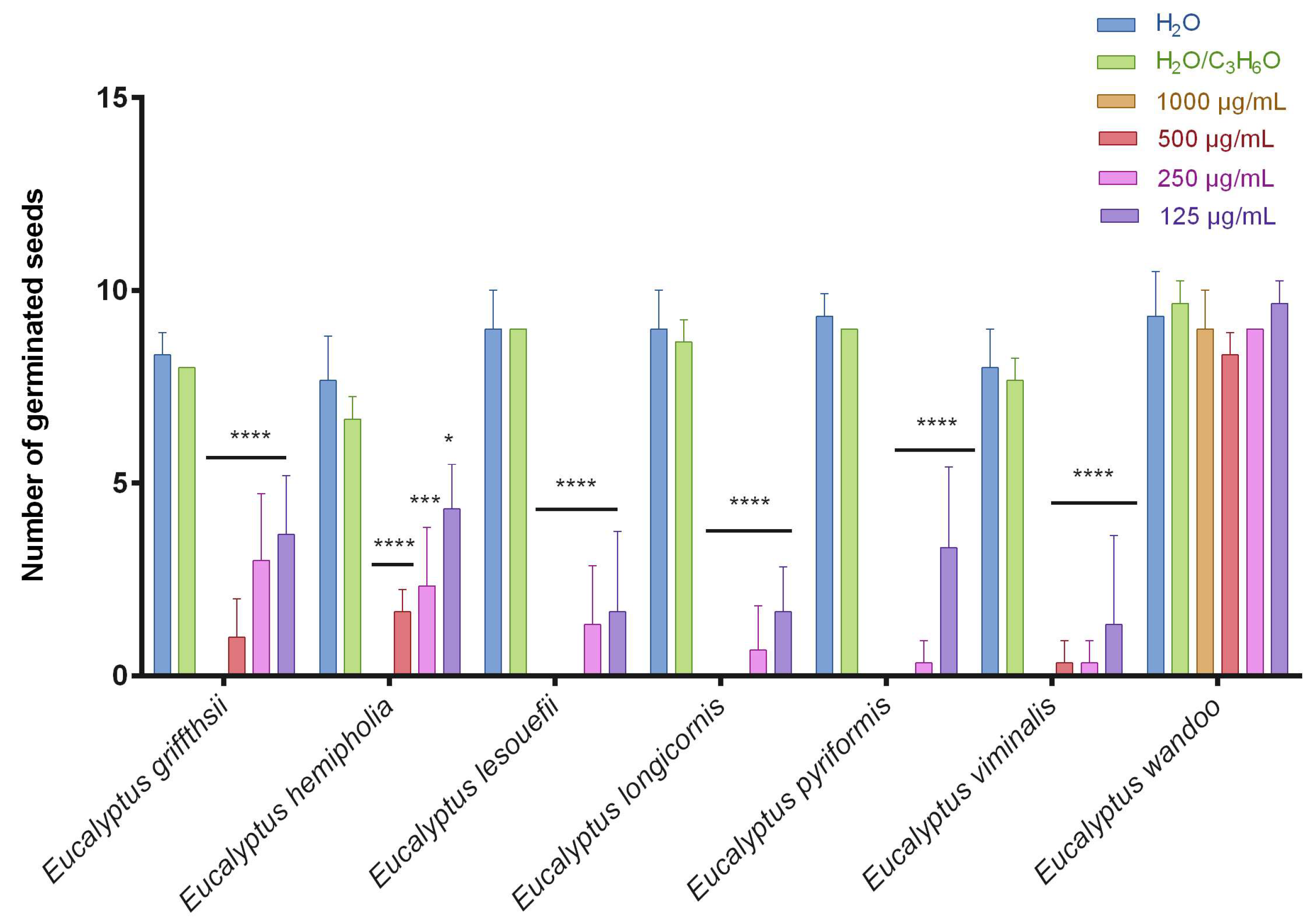
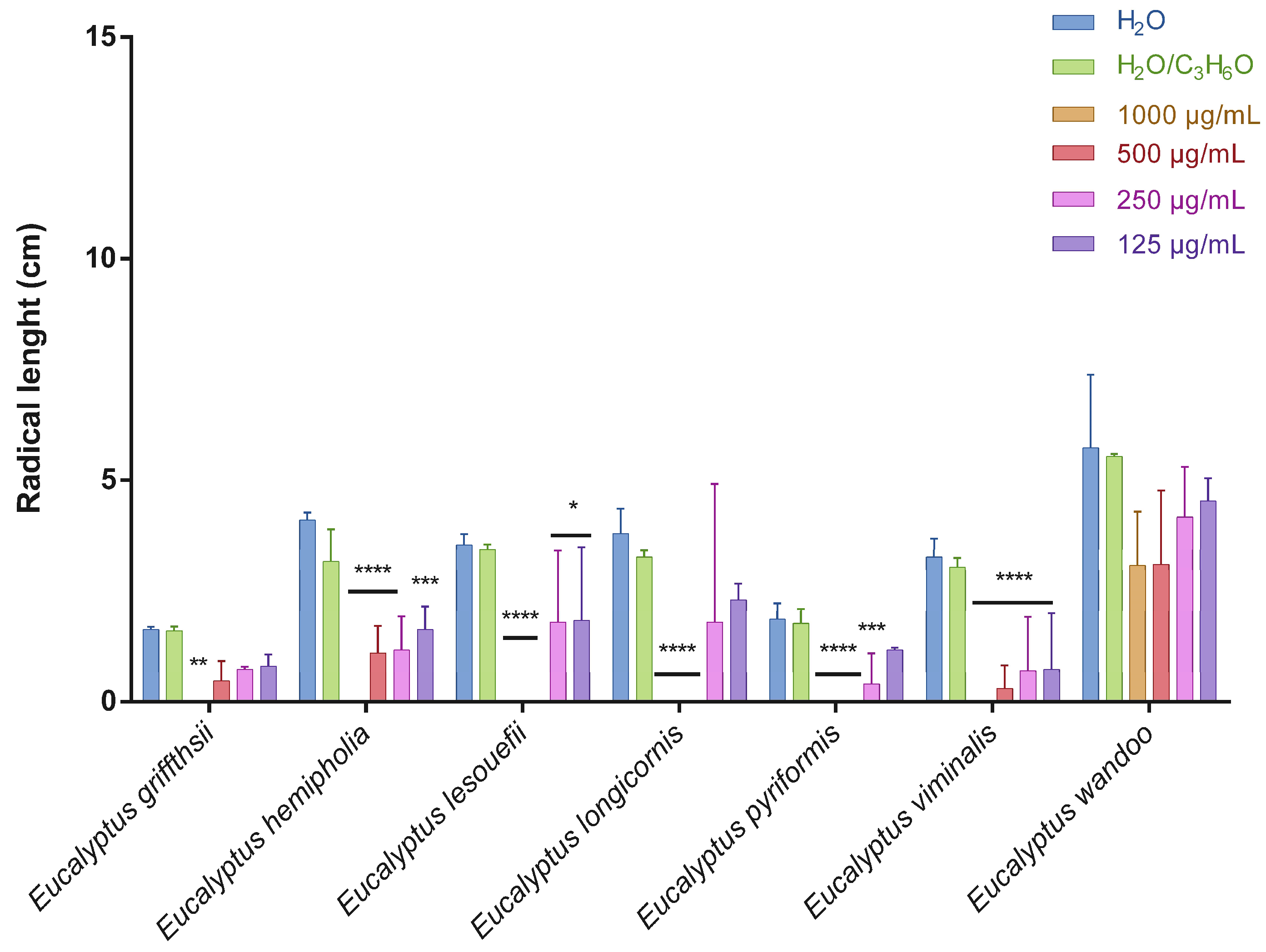
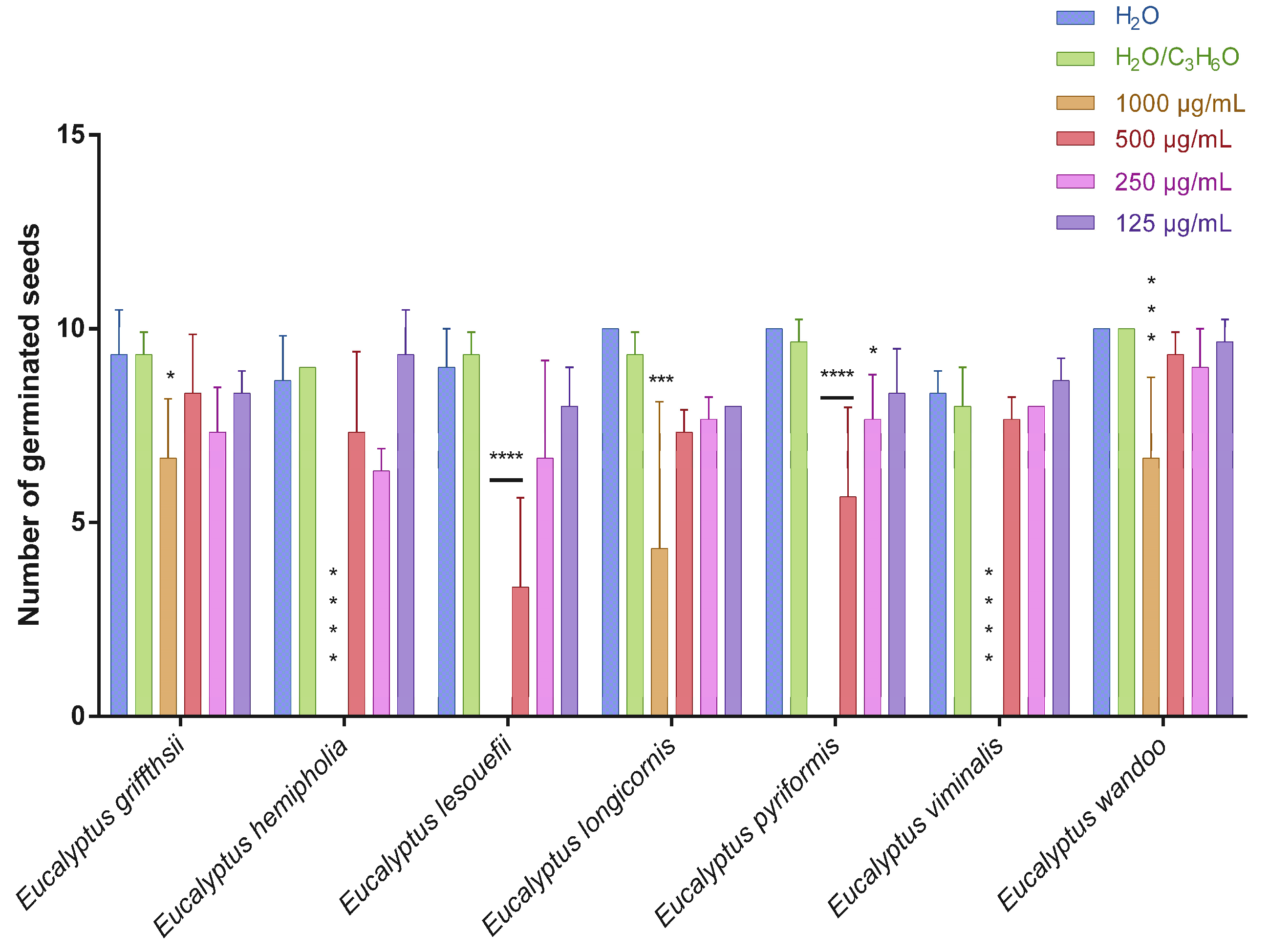
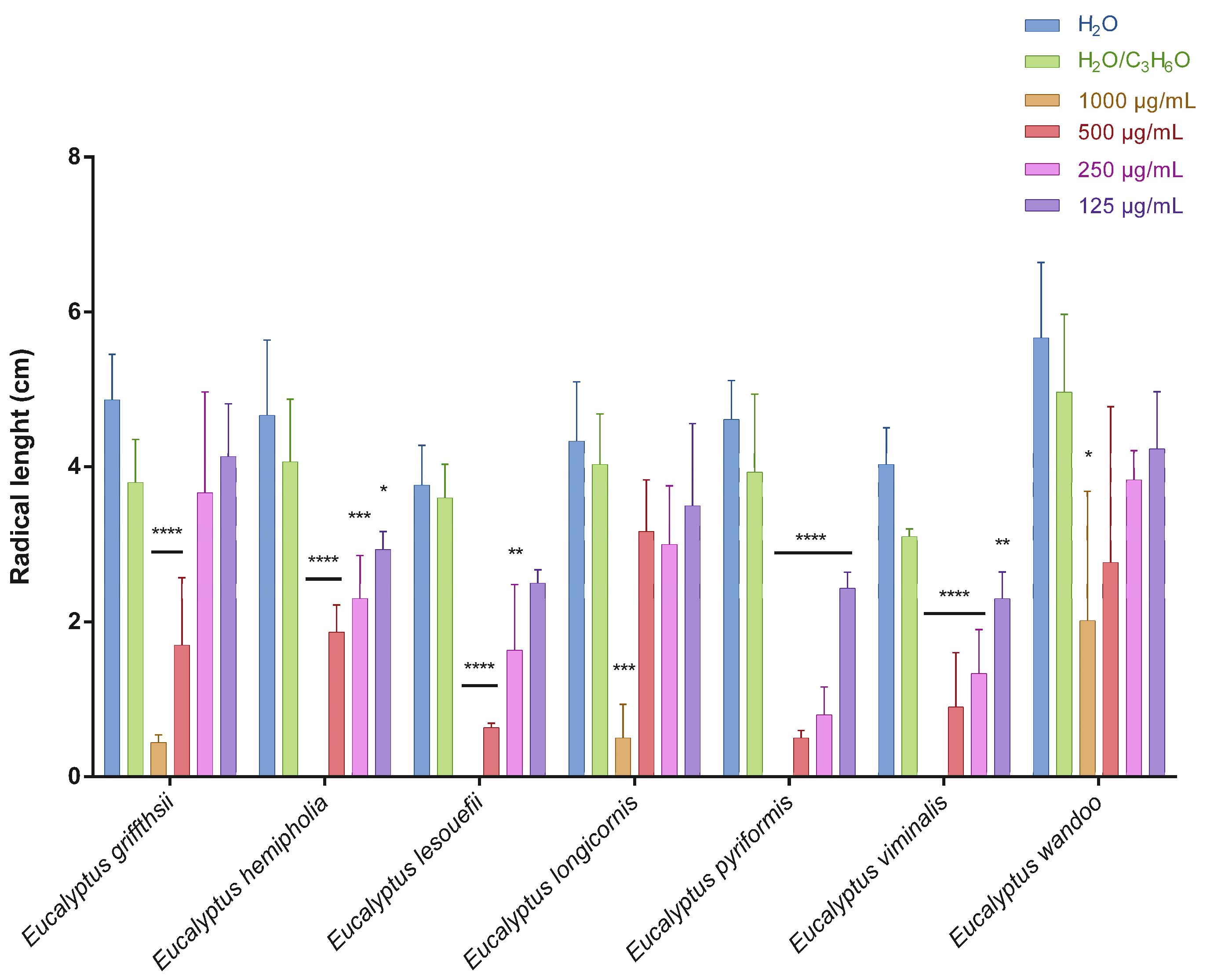
2.2. Phytotoxic Activity
2.3. Antibacterial Activity
3. Material and Methods
3.1. Plant Materials
3.2. Extraction
3.3. Analysis of the Essential Oils
3.4. Phytotoxic Activity
3.5. Antimicrobial Activity
3.5.1. Microorganisms and Culture Conditions
3.5.2. Minimal Inhibitory Concentration (MIC)
3.5.3. Biofilm Inhibitory Activity
3.5.4. Activity on Mature Bacterial Biofilm
3.5.5. Effects of the EOs on Cell Metabolic Activity within the Biofilm
3.6. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Konoshima, T.; Takasaki, M. Chemistry and bioactivity of the non-volatile constituents of eucalyptus. In Eucalyptus: The Genus Eucalyptus; Coppen, J.J.W., Ed.; Taylor & Francis: London, UK, New York, NY, USA, 2002; pp. 269–290. [Google Scholar]
- Kandasamy, O.S.; Mohamed Yassin, M.; Babu, R.C. Biology, ecology, silviculture, and potential uses of Eucalyptus—An overview. J. Med. Aromat. Plant Sci. 2000, 22, 330–335. [Google Scholar]
- Zhang, J.; An, M.; Wu, H.; Stanton, R.; Lemerle, D. Chemistry and bioactivity of Eucalyptus essential oils. Allelopathy J. 2010, 25, 313–330. [Google Scholar]
- Romagni, J.G.; Allen, S.N.; Dayan, F.E. Allelopathic effects of volatile cineoles on two weedy plant species. J. Chem. Ecol. 2000, 26, 303–313. [Google Scholar] [CrossRef]
- Batish, D.R.; Singh, H.P.; Nidhi, S.; Shalinder, K.; Kohli, R.K. Chemical composition and inhibitory activity of essential oil from decaying leaves of Eucalyptus citriodora. Z. Naturforsch. 2006, 61, 52–56. [Google Scholar] [CrossRef]
- Kaur, S.; Pal Singh, H.; Batish, D.; Kumar Kholi, R. Role of monoterpenes in Eucalyptus communities. Curr. Bioact. Compd. 2012, 8, 101–107. [Google Scholar] [CrossRef]
- Ibáñez, M.D.; Blázquez, M.A. Phytotoxicity of Essential Oils on Selected Weeds: Potential Hazard on Food Crops. Plants 2018, 7, 79. [Google Scholar] [CrossRef] [Green Version]
- Dayan, F.E.; Duke, S.O. Natural compounds as next-generation herbicides. Plant Physiol. 2014, 166, 1090–1105. [Google Scholar] [CrossRef] [Green Version]
- Flamini, G. Natural herbicides as a safer and more environmentally friendly approach to weed control: A review of the literature since 2000. Stud. Nat. Prod. Chem. 2012, 38, 353–396. [Google Scholar]
- Elaissi, A.; Rouis, Z.; Salem, N.A.B. Chemical composition of 8 eucalyptus species’ essential oils and the evaluation of their antibacterial, antifungal and antiviral activities. BMC Complement. Altern. Med. 2012, 12, 81. [Google Scholar] [CrossRef] [Green Version]
- Luís, Â.; Duarte, A.; Gominho, J.; Domingues, F.; Duarte, A.P. Chemical composition, antioxidant, antibacterial and anti-quorum sensing activities of Eucalyptus globulus and Eucalyptus radiata essential oils. Ind. Crops Prod. 2016, 79, 274–282. [Google Scholar] [CrossRef]
- Sumitra, S.; Sharma, S.K. Antibacterial activity of essential oil and root extract of Eucalyptus teriticornis. Indian J. Nat. Prod. Resour. 2005, 21, 16–17. [Google Scholar]
- Cermelli, C.; Fabio, A.; Fabio, G.; Quaglio, P. Effect of eucalyptus essential oil on respiratory bacteria and viruses. Cur. Microbiol. 2008, 56, 89–92. [Google Scholar] [CrossRef] [PubMed]
- Caputo, L.; Smeriglio, A.; Trombetta, D.; Cornara, L.; Trevena, G.; Valussi, M.; Fratianni, F.; De Feo, V.; Nazzaro, F. Chemical Composition and Biological Activities of the Essential Oils of Leptospermum petersonii and Eucalyptus gunnii. Front. Microbiol. 2020, 11, 409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ameur, E.; Sarra, M.; Yosra, D.; Mariem, K.; Nabil, A.; Lynen, F.; Larbi, K.M. Chemical composition of essential oils of eight Tunisian Eucalyptus species and their antibacterial activity against strains responsible for otitis. BMC Complement. Altern. Med. 2021, 21, 209. [Google Scholar]
- Bignell, C.M.; Dunlop, P.J.; Brophy, J.J.; Jackson, J.F. Volatile leaf oils of some south-western and southern Australian species of the genus Eucalyptus. Part I. Subgenus Symphyomyrtus, Section Dumaria, Series Incrassatae. Flavour Fragr. J. 1994, 9, 113–117. [Google Scholar] [CrossRef]
- Zrira, S.S.; Benjilali, B.B.; Fechtal, M.M.; Richard, H.H. Essential oils of twenty-seven Eucalyptus species grown in Morocco. J. Essent. Oil Res. 1992, 4, 259–264. [Google Scholar] [CrossRef]
- Bignell, C.M.; Dunlop, P.J.; Brophy, J.J.; Jackson, J.F. Volatile leaf oils of some south-western and Southern Australian species of the genus Eucalyptus. Part III—Subgenus Symphyomyrtus, Section Bisectaria, Series Macrocarpae. Flavour Fragr. J. 1994, 9, 309–313. [Google Scholar] [CrossRef]
- Faria, J.M.S.; Sanches, J.; Lima, A.S.; Mendes, M.D.; Leiria, R.; Geraldes, D.A.; Figueiredo, A.C.; Trindade, H.; Pedro, L.G.; Barroso, J.G. Eucalyptus from Mata Experimental do Escaroupim (Portugal): Evaluation of the essential oil composition from sixteen species. Acta Hortic. 2011, 925, 61–66. [Google Scholar] [CrossRef]
- Elaissi, A.; Salah, K.H.; Mabrouk, S.; Larbi, K.M.; Chemli, R.; Harzallah-Skhiri, F. Antibacterial activity and chemical composition of 20 Eucalyptus species’ essential oils. Food Chem. 2011, 129, 1427–1434. [Google Scholar] [CrossRef]
- Maghsoodlou, M.T.; Kazemipoor, N.; Valizadeh, J.; Falak Nezhad Seifi, M.; Rahneshan, N. Essential oil composition of Eucalyptus microtheca and Eucalyptus viminalis. Avicenna J. Phytomed. 2015, 5, 540–552. [Google Scholar]
- Foudil-Cherif, Y.; Meklati, B.Y.; Verzera, A.; Mondello, L.; Dugo, G. Chemical Examination of Essential Oils from the Leaves of Nine Eucalyptus Species Growing in Algeria. J. Essent. Oil Res. 2000, 12, 186–191. [Google Scholar] [CrossRef]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography/Mass Spectroscopy, 4th ed.; Allured Publishing Corporation: Carol Stream, IL, USA, 2007. [Google Scholar]
- Ramezani, S.; Saharkhiz, M.J.; Ramezani, F.; Fotokian, M.H. Use of essential oils as bioherbicides. J. Essent. Oil-Bear Plants 2008, 11, 319–327. [Google Scholar] [CrossRef]
- Rasaeifar, M.; Hosseini, N.; Asl, N.; Zandi, P.; Aghdam, A. Allelopathic effect of Eucalyptus globulus’ essential oil on seed germination and seedling establishment of Amaranthus blitoides and Cyndon dactylon. Trakia J. Sci. 2013, 11, 73–81. [Google Scholar]
- Macias, F.A.; Molinillo, J.M.; Varela, R.M.; Galindo, J.C. Allelopathy—A natural alternative for weed control. Pest Manag. Sci. 2007, 63, 327–348. [Google Scholar] [CrossRef] [PubMed]
- Mennan, H. Economic thresholds of Sinapis arvensis (wild mustard) in winter wheat fields. Pak. J. Agron. 2003, 2, 34–39. [Google Scholar] [CrossRef]
- Khammassi, M.; Polito, F.; Amri, I.; Khedhri, S.; Hamrouni, L.; Nazzaro, F.; Fratianni, F.; De Feo, V. Chemical Composition and Phytotoxic, Antibacterial and Antibiofilm Activity of the Essential Oils of Eucalyptus occidentalis, E. striaticalyx and E. stricklandii. Molecules 2022, 27, 5820. [Google Scholar] [CrossRef]
- Heap, I. The International Survey of Herbicide Resistant Weeds 2016. Available online: http://www.weedscience.org (accessed on 12 September 2022).
- Nazzaro, F.; Fratianni, F.; De Martino, L.; Coppola, R.; De Feo, V. Effect of essential oils on pathogenic bacteria. Pharmaceuticals 2013, 6, 1451–1474. [Google Scholar] [CrossRef]
- Nazzaro, F.; Fratianni, F.; Coppola, R. Quorum sensing and phytochemicals. Int. J. Mol. Sci. 2013, 14, 12607–12619. [Google Scholar] [CrossRef] [Green Version]
- Dohare, S.; Dubey, S.D.; Kalia, M.; Verma, P.; Pandey, H.; Singh, N.K.; Agarwal, V. Anti-biofilm activity of Eucalyptus globulus oil encapsulated silica nanoparticles against E. coli biofilm. Int. J. Pharm. Sci. 2014, 5, 5011. [Google Scholar]
- Sambyal, S.S.; Sharma, P.; Shrivastava, D. Anti-biofilm activity of selected plant essential oils against Pseudomonas aeruginosa and Staphylococcus aureus. Int. J. Curr. Microbiol. Appl. Sci. 2017, 6, 444–450. [Google Scholar] [CrossRef] [Green Version]
- de Oliveira Carvalho, I.; Purgato, G.A.; Píccolo, M.S.; Pizziolo, V.R.; Coelho, R.R.; Diaz-Muñoz, G.; Diaz, M.A.N. In vitro anticariogenic and antibiofilm activities of toothpastes formulated with essential oils. Arch. Oral Biol. 2020, 117, 104834. [Google Scholar] [CrossRef] [PubMed]
- Jennings, W.; Shibamoto, T. Qualitative Analysis of Flavour and Fragrance Volatiles by Glass Capillary Gas Chromatography; Academic Press: New York, NY, USA, 1980. [Google Scholar]
- Davies, N.W. Gas chromatographic retention indices of monoterpenes and sesquiterpenes on methyl silicone and Carbowax 20M phases. J. Chromatogr. 1990, 503, 1–24. [Google Scholar] [CrossRef]
- Goodner, K.L. Practical retention index models of OV-101, DB-1, DB-5, and DB-Wax for flavor and fragrance compounds. LWT Food Sci. Technol. 2008, 41, 951–958. [Google Scholar] [CrossRef]
- McLafferty, F.W. Wiley Registry of Mass Spectral Data, with NIST Spectral Data CDRom, 7th ed.; John Wiley & Sons: New York, NY, USA, 1998. [Google Scholar]
- Bewley, J.D. Seed germination and dormancy. Plant Cell 1997, 9, 1055–1066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarker, S.D.; Nahar, L.; Kumarasamy, Y. Microtitre plate-based antibacterial assay incorporating resazurin as an indicator of cell growth, and its application in the in vitro antibacterial screening of phytochemicals. Methods 2007, 42, 321–324. [Google Scholar] [CrossRef] [PubMed]
- Nazzaro, F.; Polito, F.; Amato, G.; Caputo, L.; Francolino, R.; d’Acierno, A.; Fratianni, F.; Candido, V.; Coppola, R.; De Feo, V. Chemical composition of essential oils and bulbs and aerial parts of two cultivars of Allium sativum and their biofilm activity against food and nosocomial pathogens. Antibiotics 2022, 11, 724. [Google Scholar] [CrossRef]
- Fratianni, F.; d’Acierno, A.; Ombra, M.N.; Amato, G.; De Feo, V.; Ayala-Zaval, J.F.; Coppola, R.; Nazzaro, F. Fatty acid composition and in vitro anti-inflammatory activity of five cold-pressed Prunus seed oils, and their anti-biofilm effect against pathogenic bacteria. Front. Nutr. 2021, 8, 775751. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Yield (%) | |
|---|---|
| Eucalyptus griffithsii | 1.8 |
| Eucalyptus hemiphloia | 3.6 |
| Eucalyptus lesouefii | 3.2 |
| Eucalyptus longicornis | 1.7 |
| Eucalyptus pyriformis | 2.8 |
| Eucalyptus viminalis | 1.8 |
| Eucalyptus wandoo | 1.3 |
| N | Compound Name | EG | EH | ELE | ELO | EP | EV | EW | KI a | KI b | Identification c |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | α-Thujene | 0.1 | - | - | - | - | - | - | 859 | 930 | 1, 2, 3 |
| 2 | α-Pinene | 9.8 | 0.6 | 14.6 | 3.5 | 1.9 | 7.2 | 4.3 | 864 | 939 | 1, 2, 3 |
| 3 | Camphene | 0.2 | - | 0.2 | 0.2 | 0.1 | 0.1 | 0.1 | 876 | 954 | 1, 2, 3 |
| 4 | Thuja-2,4(10)-diene | - | - | - | - | t | - | - | 882 | 960 | 1, 2 |
| 5 | β-Pinene | 3.8 | 0.1 | 4.4 | 0.1 | 3.5 | 0.1 | - | 902 | 979 | 1, 2, 3 |
| 6 | Myrcene | - | 0.1 | - | - | - | - | - | 923 | 990 | 1, 2, 3 |
| 7 | δ-3-Carene | 0.1 | - | - | - | - | - | - | 930 | 1002 | 1, 2, 3 |
| 8 | α-Phellandrene | - | 0.5 | 0.2 | - | 0.3 | 0.1 | - | 931 | 1011 | 1, 2, 3 |
| 9 | α-Terpinene | 0.1 | 0.3 | - | - | 0.4 | - | - | 944 | 1017 | 1, 2, 3 |
| 10 | o-Cymene | 4.2 | - | - | - | - | - | - | 951 | 1024 | 1, 2, 3 |
| 11 | p-Cymene | - | 5.5 | 14.3 | - | 28.8 | - | 0.6 | 952 | 1026 | 1, 2, 3 |
| 12 | Eucalyptol | 48.9 | 63.2 | 40.8 | 84.2 | 15.1 | 68.1 | 73.6 | 958 | 1031 | 1, 2, 3 |
| 13 | (Z)-β-Ocimene | - | - | - | - | 0.4 | - | - | 983 | 1037 | 1, 2, 3 |
| 14 | γ-Terpinene | 0.6 | 1.3 | 0.5 | - | - | 1.1 | 984 | 1059 | 1, 2, 3 | |
| 15 | cis-Sabinene hydrate | - | - | - | - | 0.1 | - | - | 994 | 1070 | 1, 2 |
| 16 | cis-Linalool oxide | - | 0.3 | - | - | 0.2 | - | - | 997 | 1072 | 1, 2 |
| 17 | Terpinolene | 0.1 | - | - | - | - | - | - | 1007 | 1088 | 1, 2, 3 |
| 18 | p-Mentha-3,8-diene | - | - | - | - | 0.3 | - | - | 1008 | 1072 | 1, 2 |
| 19 | trans-Linalool oxide | - | 0.3 | - | - | - | - | - | 1009 | 1086 | 1, 2 |
| 20 | m-Cymenene | - | - | - | - | 0.5 | - | - | 1010 | 1085 | 1, 2 |
| 21 | p-Cymenene | 0.2 | 0.3 | - | - | - | - | - | 1010 | 1091 | 1, 2, 3 |
| 22 | α-Pinene oxide | - | - | 0.1 | - | - | - | - | 1010 | 1099 | 1, 2 |
| 23 | 6-Camphenol | - | - | - | - | - | 0.2 | - | 1010 | 1113 | 1, 2 |
| 24 | 6-Camphenone | - | - | - | - | 0.1 | - | 0.1 | 1018 | 1096 | 1, 2 |
| 25 | Linalool | - | 0.4 | - | - | - | - | - | 1024 | 1096 | 1, 2, 3 |
| 26 | cis-Thujone | 0.4 | 0.2 | - | - | 0.4 | - | - | 1022 | 1102 | 1, 2, 3 |
| 27 | Isoamyl isovalerate | 0.6 | 0.5 | 0.4 | 0.4 | - | - | - | 1029 | 1, 2 | |
| 28 | Isopentyl isovalerate | - | - | - | - | 0.1 | - | - | 1029 | 1, 2 | |
| 29 | exo-Fenchol | - | 0.1 | 0.6 | 0.4 | - | 0.4 | 0.2 | 1031 | 1121 | 1, 2, 3 |
| 30 | trans-Thujol | 0.1 | - | - | - | 0.4 | - | - | 1033 | 1, 2, 3 | |
| 31 | trans-p-Mentha-2,8-dien-1-ol | - | 3.2 | - | - | - | - | - | 1040 | 1122 | 1, 2 |
| 32 | cis-p -Menth-2-en-1-ol | 0.1 | - | - | - | 0.3 | - | - | 1041 | 1121 | 1, 2 |
| 33 | α-Campholenal | 0.4 | 0.4 | 0.3 | 0.1 | 0.8 | 0.2 | 0,1 | 1043 | 1126 | 1, 2 |
| 34 | cis- p-Mentha-2,8-dien-1-ol | - | - | - | - | 0.1 | - | - | 1047 | 1137 | 1, 2 |
| 35 | allo-Ocimene | 0.3 | - | - | - | - | 0.1 | - | 1051 | 1132 | 1, 2, 3 |
| 36 | Terpineol | - | - | 0.4 | - | - | - | - | 1051 | 1133 | 1, 2, 3 |
| 37 | iso-3-Thujanol | - | - | - | - | 0.7 | - | - | 1052 | 1138 | 1, 2 |
| 38 | trans-Pinocarveol | 4.2 | 0.3 | 4.9 | 4.2 | 4.4 | 3.9 | 6.1 | 1055 | 1139 | 1, 2 |
| 39 | Camphor | 0.2 | - | - | 0.4 | - | - | - | 1058 | 1146 | 1, 2, 3 |
| 40 | trans-p-Menth-2-en-1-ol | - | 0.3 | - | - | 0.3 | - | - | 1059 | 1140 | 1, 2 |
| 41 | Sabina ketone | 2.0 | - | 1.4 | - | 3.9 | 1.2 | - | 1067 | 1159 | 1, 2, 3 |
| 43 | cis-Verbenol | 0.3 | - | - | - | 0.2 | - | - | 1064 | 1141 | 1, 2, 3 |
| 44 | Pinocarvone | 1.3 | 0.3 | 1.3 | 0.9 | 1.3 | 1.2 | 1.3 | 1077 | 1164 | 1, 2, 3 |
| 45 | Borneol | 0.6 | 0.4 | 0.9 | 0.5 | 0.3 | 0.5 | 0.4 | 1084 | 1169 | 1, 2, 3 |
| 46 | Menthol | - | - | - | - | - | 0.2 | - | 1084 | 1171 | 1, 2, 3 |
| 47 | p-Mentha-1,5-dien-8-ol | - | - | - | 0.2 | 0.6 | - | - | 1084 | 1170 | 1, 2 |
| 48 | Umbellunone | 0.3 | - | - | - | - | - | - | 1087 | 1171 | 1, 2 |
| 49 | trans-2-hydroxy-Pinocamphone | - | - | - | - | - | - | 0.7 | 1093 | 1250 | 1, 2 |
| 50 | Terpinen-4-ol | 1.8 | 1.7 | 1.5 | 0.1 | 3 | 1.1 | - | 1094 | 1177 | 1, 2 |
| 51 | neo-Verbenol | 2.9 | - | 1.6 | - | - | - | - | 1093 | 1, 2 | |
| 52 | Cryptone | - | - | - | - | - | 1.4 | - | 1094 | 1185 | 1, 2 |
| 53 | m-Cymen-8-ol | - | - | - | - | 8.8 | - | - | 1096 | 1179 | 1, 2 |
| 54 | iso-Menthol | - | 4.6 | - | - | - | - | - | 1096 | 1182 | 1, 2 |
| 55 | p-Cymen-8-ol | 0.6 | 1.0 | - | - | - | - | - | 1099 | 1182 | 1, 2, 3 |
| 56 | cis-Pinocarveol | - | - | - | 0.4 | - | - | - | 1098 | 1184 | 1, 2 |
| 58 | α-Terpinyl acetate | - | - | - | - | - | - | 1.9 | 1101 | 1349 | 1, 2 |
| 59 | Dihydro carveol | 1.6 | - | 3.9 | - | - | 0.2 | 0.1 | 1102 | 1193 | 1, 2 |
| 60 | α-Terpineol | 1.9 | 1.2 | 1.7 | 0.8 | - | - | 0.1 | 1102 | 1188 | 1, 2, 3 |
| 61 | γ-Terpineol | - | - | - | - | 1.9 | 1.0 | - | 1104 | 1199 | 1, 2, 3 |
| 62 | trans-Carveol | - | - | - | - | 1.7 | - | - | 1109 | 1216 | 1, 2 |
| 63 | Myrtenol | 0.2 | - | - | - | - | - | - | 1112 | 1195 | 1, 2, 3 |
| 64 | trans-Pulegol | - | - | - | - | - | 0.3 | - | 1112 | 1214 | 1, 2, 3 |
| 65 | Verbenone | 0.3 | 0.8 | - | - | 1.7 | - | - | 1113 | 1205 | 1, 2, 3 |
| 66 | 4-Methyleneisophorone | - | - | - | - | 0.7 | - | - | 1118 | 1, 2 | |
| 67 | cis-Carveol | 0.2 | - | - | - | - | - | - | 1132 | 1229 | 1, 2, 3 |
| 68 | cis-p-Mentha-1(7),8-dien-2-ol | - | - | - | - | 0.3 | - | - | 1133 | 1230 | 1, 2 |
| 69 | (E)-Ocimenone | 0.3 | - | - | - | - | - | - | 1141 | 1238 | 1, 2 |
| 70 | (Z)-Ocimenone | - | 0.3 | - | - | - | - | - | 1142 | 1229 | 1, 2 |
| 71 | Pulegone | - | - | - | - | 0.1 | - | - | 1142 | 1237 | 1, 2, 3 |
| 72 | Cumin aldehyde | 1.9 | 4 | 1.0 | - | 5.2 | 1.0 | - | 1150 | 1241 | 1, 2, 3 |
| 73 | Carvone | 0.9 | - | - | - | - | - | - | 1155 | 1243 | 1, 2, 3 |
| 74 | trans-p-Mentha-1(7),8-dien-2-ol | - | - | - | 0.5 | - | 0.7 | - | 1143 | 1189 | 1, 2 |
| 75 | trans-Piperitol acetate | - | - | - | - | 2.2 | - | - | 1154 | 1346 | 1, 2 |
| 76 | Piperitone | 0.4 | 0.6 | - | - | - | - | - | 1163 | 1252 | 1, 2 |
| 77 | cis-p-Mentha-8-thiol-3-one | - | - | - | - | 2.4 | - | - | 1182 | 1360 | 1, 2 |
| 78 | Perilla aldehyde | - | 2.2 | - | - | - | - | - | 1182 | 1271 | 1, 2, 3 |
| 79 | cis-Carvone oxide | - | - | - | - | - | 0.3 | - | 1183 | 1263 | 1, 2 |
| 80 | p-Menth-1-en-7-al | 1.2 | - | 0.5 | - | - | 0.2 | - | 1184 | 1275 | 1, 2 |
| 81 | α-Terpinen-7-al | 0.2 | 0.3 | - | - | - | - | - | 1198 | 1285 | 1, 2 |
| 82 | p-Cymen-7-ol | 0.6 | - | - | - | - | - | - | 1204 | 1290 | 1, 2 |
| 83 | Carvacrol acetate | - | - | - | - | 1.9 | - | 0.1 | 1208 | 1372 | 1, 2 |
| 84 | Carvacrol | 1.0 | 1.8 | - | - | - | 0.3 | - | 1230 | 1299 | 1, 2, 3 |
| 85 | δ-Elemene | - | - | 0.1 | - | - | - | - | 1230 | 1338 | 1, 2, 3 |
| 86 | Piperitenone | 0.3 | - | - | - | - | - | - | 1244 | 1343 | 1, 2 |
| 87 | trans-Carvyl acetate | - | - | - | 0.1 | - | - | 0.6 | 1244 | 1342 | 1, 2 |
| 88 | (E)-Jasmonyl acetate | - | - | - | - | 0.5 | - | - | 1252 | 1398 | 1, 2 |
| 89 | allo-Aromadendrene- | - | - | - | - | - | 0.6 | - | 1327 | 1460 | 1, 2, 3 |
| 90 | dehydro-Aromadendrane | - | - | - | - | - | 0.4 | - | 1348 | 1462 | 1, 2 |
| 91 | α-Vetispirene | - | - | - | - | - | 0.1 | - | 1376 | 1490 | 1, 2 |
| 92 | cis-β-Guaiene | - | - | - | - | 0.2 | - | - | 1384 | 1493 | 1, 2 |
| 93 | Viridiflorene | - | - | - | - | - | 0.1 | 0.6 | 1383 | 1496 | 1, 2, 3 |
| 94 | 2-Propyl-heptanol | - | - | - | 0.1 | - | - | - | 1389 | 1, 2 | |
| 95 | Phenetyl pivalate | - | - | - | 0.3 | - | - | - | 1393 | 1, 2 | |
| 96 | Germacrene B | - | - | - | - | - | - | 0.3 | 1441 | 1561 | 1, 2, 3 |
| 97 | Spathulenol | 1.3 | 0.2 | 1.2 | - | 0.2 | 5.1 | 2.1 | 1459 | 1578 | 1, 2, 3 |
| 98 | Caryophyllene oxide | - | 0.2 | - | - | 0.4 | - | - | 1462 | 1583 | 1, 2, 3 |
| 99 | Guaiol | 0.2 | - | - | - | - | - | - | 1464 | 1600 | 1, 2, 3 |
| 100 | Viridiflorol | - | - | - | - | 0.3 | - | 0.2 | 1465 | 1592 | 1, 2, 3 |
| 101 | Globulol | - | - | - | 0.1 | - | 0.2 | - | 1466 | 1590 | 1, 2, 3 |
| 102 | Cubeban-11-ol | - | - | - | - | - | - | 0.1 | 1474 | 1595 | 1, 2, 3 |
| 103 | Rosifoliol | - | - | - | - | - | 0.1 | 1.0 | 1496 | 1600 | 1, 2, 3 |
| 104 | 1-epi-Cubenol | - | - | - | - | 0.1 | - | - | 1502 | 1628 | 1, 2, 3 |
| 105 | α-Eudesmol | - | - | - | - | 0.2 | - | 0.9 | 1524 | 1653 | 1, 2, 3 |
| 106 | β-Eudesmol | - | - | - | 0.1 | - | - | 1.5 | 1526 | 1650 | 1, 2, 3 |
| Total | 96.7 | 97.5 | 96.8 | 97.6 | 97.3 | 96.6 | 98.0 | ||||
| Monoterpene hydrocarbons | 19.5 | 9.1 | 34.2 | 3.8 | 36.2 | 7.6 | 6.1 | ||||
| Oxygenated monoterpenes | 73.2 | 81.3 | 59.9 | 92.8 | 53.9 | 81.4 | 85.2 | ||||
| Sesquiterpene hydrocarbons | - | - | 0.1 | - | 0.7 | 1.2 | 0.9 | ||||
| Oxygenated sesquiterpenes | 1.5 | 0.4 | 1.2 | 0.2 | 1.2 | 5.4 | 5.8 | ||||
| Other | 2.5 | 6.7 | 1.4 | 0.8 | 5.3 | 1 | - |
| MIC (µL/mL) | |||||
|---|---|---|---|---|---|
| A. baumannii | E. coli | L. monocytogenes | P. aeruginosa | S. aureus | |
| E. griffithsii | 25 ± 1 ** | 25 ± 1 | 26 ± 1 | 26 ± 1 | 25 ± 1 |
| E. hemiphloia | 26 ± 2 | 26 ± 1 * | 28 ± 1 ** | 30 ± 2 ** | 28 ± 1 *** |
| E. lesoufii | 28 ± 1 ** | 28 ± 1 *** | 26 ± 1 | 27 ± 2 | 27 ± 1 ** |
| E. longicornis | 30 ± 1 *** | 30 ± 1 **** | 38 ± 2 **** | 27 ± 2 | 35 ± 1 **** |
| E. pyriformis | 25 ± 1 | 25 ± 1 | 26 ± 2 | 26 ± 1 | 26 ± 2 * |
| E. viminalis | 30 ± 1 *** | 28 ± 1 *** | 25 ± 1 | 25 ± 1 | 30 ± 1 **** |
| E. wandoo | 26 ± 1 | 29 ± 1 **** | 35 ± 1 **** | 26 ± 1 | 29 ± 1 **** |
| Tetracycline | 24 ± 2 | 23 ± 1 | 23 ± 1 | 24 ± 2 | 23 ± 1 |
| Time 0 | A. baumannii | E. coli | L. monocytogenes | P. aeruginosa | S. aureus |
|---|---|---|---|---|---|
| EG 10 µL/mL | 82.48 ± 0.2 **** | 85.44 ± 0.2 **** | 85.03 ± 0.1 **** | 89.62 ± 0.2 **** | 87.17 ± 0.1 **** |
| EG 20 µL/mL | 92.53 ± 0.1 **** | 86.39 ± 0.2 **** | 87.56 ± 0.2 **** | 91.40 ± 0.1 **** | 93.73 ± 0.1 **** |
| EH 10 µL/mL | 37.53 ± 0.2 **** | 47.00 ± 0.3 **** | 77.59 ± 0.3 **** | 78.81 ± 0.2 **** | 84.23 ± 0.1 **** |
| EH 20 µL/mL | 74.98 ± 0.3 **** | 78.44 ± 0.4 **** | 81.06 ± 0.3 **** | 79.71 ± 0.3 **** | 84.99 ± 0.2 **** |
| EP 10 µL/mL | 92.08 ± 0.2 **** | 89.71 ± 0.2 **** | 79.13 ± 0.2 **** | 84.81 ± 0.1 **** | 88.51 ± 0.1 **** |
| EP 20 µL/mL | 92.37 ± 0.1 **** | 94.39 ± 0.1 **** | 86.65 ± 0.2 **** | 90.64 ± 0.1 **** | 95.04 ± 0.2 **** |
| ELE 10 µL/mL | 73.89 ± 0.3 **** | 60.58 ± 0.3 **** | 0.00 ± 0.00 | 64.08 ± 0.3 **** | 77.32 ± 0.2 **** |
| ELE 20 µL/mL | 79.61 ± 0.2 **** | 89.92 ± 0.2 **** | 92.14 ± 0.1 **** | 89.48 ± 0.1 **** | 86.03 ± 0.2 **** |
| ELO 10 µL/mL | 84.82 ± 0.1 **** | 85.64 ± 0.2 **** | 82.19 ± 0.3 **** | 80.91 ± 0.2 **** | 62.11 ± 0.3 **** |
| ELO 20 µL/mL | 86.09 ± 0.1 **** | 88.16 ± 0.1 **** | 88.14 ± 0.3 **** | 87.38 ± 0.2 **** | 73.98 ± 0.2 **** |
| EV 10 µL/mL | 59.39 ± 0.1 **** | 54.29 ± 0.2 **** | 0.00 ± 0.00 | 83.31 ± 0.3 **** | 4.36 ± 0.1 **** |
| EV 20 µL/mL | 63.16 ± 0.2 **** | 77.07 ± 0.2 **** | 98.67 ± 0.1 **** | 93.70 ± 0.1 **** | 58.84 ± 0.2 **** |
| EW 10 µL/mL | 89.15 ± 0.2 **** | 77.64 ± 0.1 **** | 0.00 ± 0.00 | 89.24 ± 0.2 **** | 59.16 ± 0.2 **** |
| EW 20 µL/mL | 89.43 ± 0.2 **** | 91.00 ± 0.1 **** | 88.78 ± 0.2 **** | 90.09 ± 0.1 **** | 75.90 ± 0.2 **** |
| 24 h | A. baumannii | E. coli | L. monocytogenes | P. aeruginosa | S. aureus |
| EG 10 µL/mL | 67.88 ± 0.3 **** | 33.66 ± 0.2 **** | 24.31 ± 0.2 **** | 78.64 ± 0.2 **** | 71.21 ± 0.4 **** |
| EG 20 µL/mL | 68.39 ± 0.2 **** | 33.31 ± 0.2 **** | 35.76 ± 0.1 **** | 81.79 ± 0.3 **** | 87.15 ± 0.2 **** |
| EH 10 µL/mL | 9.32 ± 0.1 **** | 52.23 ± 0.2 **** | 6.10 ± 0.1 **** | 53.15 ± 0.2 **** | 73.93 ± 0.2 **** |
| EH 20 µL/mL | 51.64 ± 0.2 **** | 53.28 ± 0.2 **** | 25.63 ± 0.2 **** | 59.50 ± 0.1 **** | 75.65 ± 0.3 **** |
| EP 10 µL/mL | 52.16 ± 0.2 **** | 26.85 ± 0.2 **** | 6.18 ± 0.1 **** | 59.35 ± 0.2 **** | 82.33 ± 0.2 **** |
| EP 20 µL/mL | 71.31 ± 0.2 **** | 36.34 ± 0.2 **** | 67.88 ± 0.2 **** | 87.56 ± 0.3 **** | 88.84 ± 0.3 **** |
| ELE 10 µL/mL | 52.15 ± 0.2 **** | 62.56 ± 0.2 **** | 0.00 ± 0.00 | 34.58 ± 0.2 **** | 52.13 ± 0.4 **** |
| ELE 20 µL/mL | 70.90 ± 0.3 | 65.96 ± 0.2 **** | 61.19 ± 0.2 **** | 57.35 ± 0.2 **** | 58.20 ± 0.4 **** |
| ELO 10 µL/mL | 62.15 ± 0.3 **** | 1.77 ± 0.2 * | 63.33 ± 0.2 **** | 52.63 ± 0.3 **** | 55.23 ± 0.3 **** |
| ELO 20 µL/mL | 68.94 ± 0.2 **** | 50.42 ± 0.2 **** | 65.22 ± 0.1 **** | 52.71 ± 0.2 **** | 60.41 ± 0.3 **** |
| EV 10 µL/mL | 14.70 ± 0.1 **** | 0.30 ± 0.2 | 0.00 ± 0.00 | 5.49 ± 0.1 *** | 50.77 ± 0.2 **** |
| EV 20 µL/mL | 25.29 ± 0.1 **** | 39.64 ± 0.2 **** | 4.39 ± 0.1 **** | 20.44 ± 0.1 **** | 53.49 ± 0.1 **** |
| EW 10 µL/mL | 30.58 ± 0.1 **** | 25.46 ± 0.2 **** | 0.00 ± 0.00 | 56.68 ± 0.2 **** | 94.55 ± 0.1 **** |
| EW 20 µL/mL | 36.92 ± 0.2 **** | 41.65 ± 0.2 **** | 57.27 ± 0.2 **** | 63.53 ± 0.1 **** | 95.83 ± 0.1 **** |
| Time 0 | A. baumannii | E. coli | L. monocytogenes | p. aeruginosa | S. aureus |
|---|---|---|---|---|---|
| EG 10 µL/mL | 83.35 ± 0.3 **** | 75.29 ± 0.3 **** | 80.04 ± 0.2 **** | 79.71 ± 0.3 **** | 79.03 ± 0.2 **** |
| EG 20 µL/mL | 85.12 ± 0.3 **** | 78.23 ± 0.3 **** | 83.22 ± 0.1 **** | 81.93 ± 0.2 **** | 81.77 ± 0.1 **** |
| EH 10 µL/mL | 82.30 ± 0.1 **** | 69.79 ± 0.3 **** | 79.21 ± 0.2 **** | 75.79 ± 0.1 **** | 79.64 ± 0.3 **** |
| EH 20 µL/mL | 83.74 ± 0.1 **** | 79.41 ± 0.3 **** | 79.48 ± 0.3 **** | 78.77 ± 0.2 **** | 80.70 ± 0.2 **** |
| EP 10 µL/mL | 80.37 ± 0.1 **** | 48.89 ± 0.1 **** | 39.63 ± 0.1 **** | 72.40 ± 0.2 **** | 29.93 ± 0.2 **** |
| EP 20 µL/mL | 83.98 ± 0.2 **** | 79.52 ± 0.3 **** | 73.00 ± 0.2 **** | 78.49 ± 0.3 **** | 80.94 ± 0.1 **** |
| ELE 10 µL/mL | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| ELE 20 µL/mL | 71.98 ± 0.3 **** | 90.71 ± 0.1 **** | 82.25 ± 0.1 **** | 74.74 ± 0.2 **** | 88.08 ± 0.3 **** |
| ELO 10 µL/mL | 21.26 ± 0.2 **** | 72.66 ± 0.1 **** | 0.00 ± 0.00 | 43.38 ± 0.4 **** | 0.27 ± 0.00 |
| ELO 20 µL/mL | 30.43 ± 0.2 **** | 74.94 ± 0.2 **** | 15.59 ± 0.1 **** | 53.92 ± 0.3 **** | 41.34 ± 0.3 **** |
| EV 10 µL/mL | 22.36 ± 0.1 **** | 58.87 ± 0.2 **** | 78.17 ± 0.3 **** | 65.27 ± 0.3 **** | 70.23 ± 0.2 **** |
| EV 20 µL/mL | 53.64 ± 0.2 **** | 70.03 ± 0.1 **** | 92.76 ± 0.1 **** | 65.13 ± 0.3 **** | 79.60 ± 0.4 **** |
| EW 10 µL/mL | 52.84 ± 0.2 | 20.14 ± 0.1 | 20.67 ± 0.1 **** | 1.06 ± 0.00 **** | 11.91 ± 0.1 |
| EW 20 µL/mL | 57.60 ± 0.4 **** | 63.66 ± 0.2 | 75.41 ± 0.4 **** | 74.42 ± 0.2 **** | 55.43 ± 0.2 **** |
| 24 h | A. baumannii | E. coli | L. monocytogenes | P. aeruginosa | S. aureus |
| EG 10 µL/mL | 0.00 ± 0.00 | 20.59 ± 0.2 **** | 0.00 ± 0.00 | 0.00 ± 0.00 | 7.95 ± 0.2 **** |
| EG 20 µL/mL | 0.00 ± 0.00 | 58.68 ± 0.2 **** | 0.00 ± 0.00 | 0.00 ± 0.00 | 28.29 ± 0.2 **** |
| EH 10 µL/mL | 0.00 ± 0.00 | 31.89 ± 0.2 **** | 0.00 ± 0.00 | 0.00 ± 0.00 | 16.72 ± 0.2 **** |
| EH 20 µL/mL | 0.00 ± 0.00 | 36.54 ± 0.2 **** | 0.00 ± 0.00 | 0.00 ± 0.00 | 21.38 ± 0.2 **** |
| EP 10 µL/mL | 2.09 ± 0.00 **** | 33.52 ± 0.2 **** | 29.90 ± 0.2 **** | 0 ± 0.2 | 30.88 ± 0.2 **** |
| EP 20 µL/mL | 35.07 ± 0.2 **** | 51.34 ± 0.2 **** | 54.04 ± 0.2 **** | 7.09 ± 0.2 **** | 33.22 ± 0.2 **** |
| ELE 10 µL/mL | 0.00 ± 0.00 | 31.89 ± 0.2 **** | 0.00 ± 0.00 | 33.95 ± 0.2 | 11.46 ± 0.2 **** |
| ELE 20 µL/mL | 86.99 ± 0.1 **** | 34.32 ± 0.2 **** | 0.00 ± 0.00 | 0.00 ± 0.00 | 18.77 ± 0.2 **** |
| ELO 10 µL/mL | 0.00 ± 0.00 | 16.48 ± 0.2 **** | 0.00 ± 0.00 | 4.16 ± 0.00 **** | 8.67 ± 0.2 **** |
| ELO 20 µL/mL | 0.00 ± 0.00 | 38.92 ± 0.2 **** | 0.00 ± 0.00 | 17.08 ± 0.2 **** | 18.97 ± 0.2 **** |
| EV 10 µL/mL | 0.00 ± 0.00 | 0 ± 0.2 **** | 27.16 ± 0.2 **** | 42.96 ± 0.2 **** | 21.21 ± 0.2 **** |
| EV 20 µL/mL | 65.38 ± 0.3 **** | 38.39 ± 0.2 **** | 27.90 ± 0.2 **** | 83.43 ± 0.1 **** | 73.94 ± 0.2 **** |
| EW 10 µL/mL | 0.00 ± 0.00 | 10.32 ± 0.2 **** | 0.00 ± 0.00 | 0.00 ± 0.00 | 18.43 ± 0.2 **** |
| EW 20 µL/mL | 19.87 ± 0.2 **** | 27.08 ± 0.2 **** | 0.00 ± 0.00 | 9.73 ± 0.1 **** | 30.54 ± 0.2 **** |
| Date of Harvest | Srboretum (Region) | |
|---|---|---|
| Eucalyptus griffithsii | April 2021 | Henchir Naam (Siliana) |
| Eucalyptus hemiphloia | May 2021 | Djebel Manasour (Zaghouen) |
| Eucalyptus lesouefii | April 2021 | Henchir Naam (Siliana) |
| Eucalyptus longicornis | April 2021 | Henchir Naam (Siliana) |
| Eucalyptus pyriformis | May 2021 | Henchir Naam (Siliana) |
| Eucalyptus viminalis | May 2021 | Souiniet (Jendouba) |
| Eucalyptus wandoo | May 2021 | Djebel Manasour (Zaghouen) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khedhri, S.; Polito, F.; Caputo, L.; Manna, F.; Khammassi, M.; Hamrouni, L.; Amri, I.; Nazzaro, F.; De Feo, V.; Fratianni, F. Chemical Composition, Phytotoxic and Antibiofilm Activity of Seven Eucalyptus Species from Tunisia. Molecules 2022, 27, 8227. https://doi.org/10.3390/molecules27238227
Khedhri S, Polito F, Caputo L, Manna F, Khammassi M, Hamrouni L, Amri I, Nazzaro F, De Feo V, Fratianni F. Chemical Composition, Phytotoxic and Antibiofilm Activity of Seven Eucalyptus Species from Tunisia. Molecules. 2022; 27(23):8227. https://doi.org/10.3390/molecules27238227
Chicago/Turabian StyleKhedhri, Sana, Flavio Polito, Lucia Caputo, Francesco Manna, Marwa Khammassi, Lamia Hamrouni, Ismail Amri, Filomena Nazzaro, Vincenzo De Feo, and Florinda Fratianni. 2022. "Chemical Composition, Phytotoxic and Antibiofilm Activity of Seven Eucalyptus Species from Tunisia" Molecules 27, no. 23: 8227. https://doi.org/10.3390/molecules27238227
APA StyleKhedhri, S., Polito, F., Caputo, L., Manna, F., Khammassi, M., Hamrouni, L., Amri, I., Nazzaro, F., De Feo, V., & Fratianni, F. (2022). Chemical Composition, Phytotoxic and Antibiofilm Activity of Seven Eucalyptus Species from Tunisia. Molecules, 27(23), 8227. https://doi.org/10.3390/molecules27238227