



Protective Effects of Hippophae rhamnoides L. Phenylpropanoids on Doxorubicin-Induced Cardiotoxicity in Zebrafish

 ,
, 
Abstract
:1. Introduction
2. Results
2.1. Effects of H. rhamnoides L. Phenylpropanoids on Dox-Induced Cardiac Injury in Zebrafish
2.2. Effects of H. rhamnoides L. Phenylpropanoids on Locomotor Activity in Zebrafish
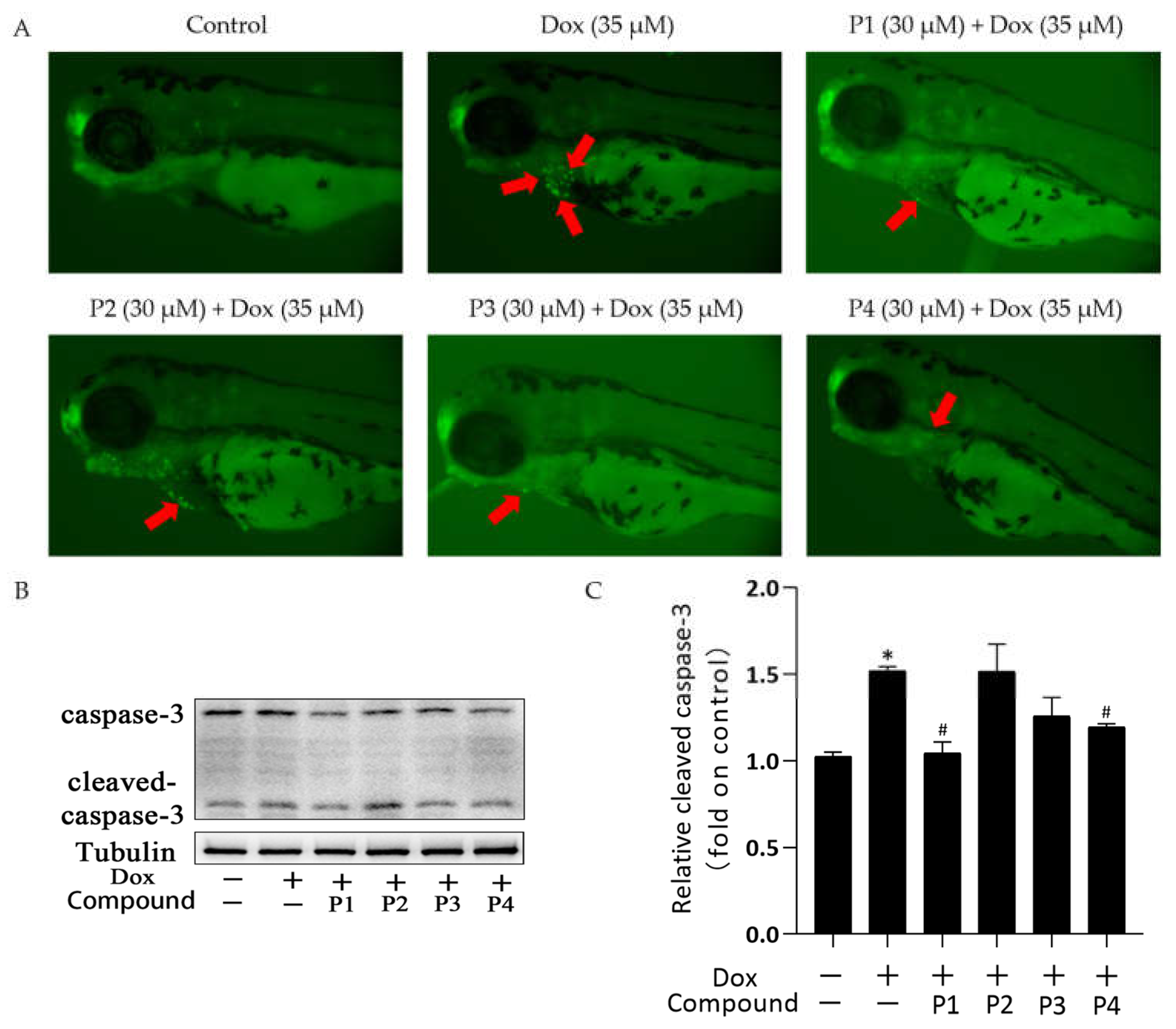
2.3. Effects of H. rhamnoides L. Phenylpropanoids on Dox-Induced Apoptosis in Zebrafish
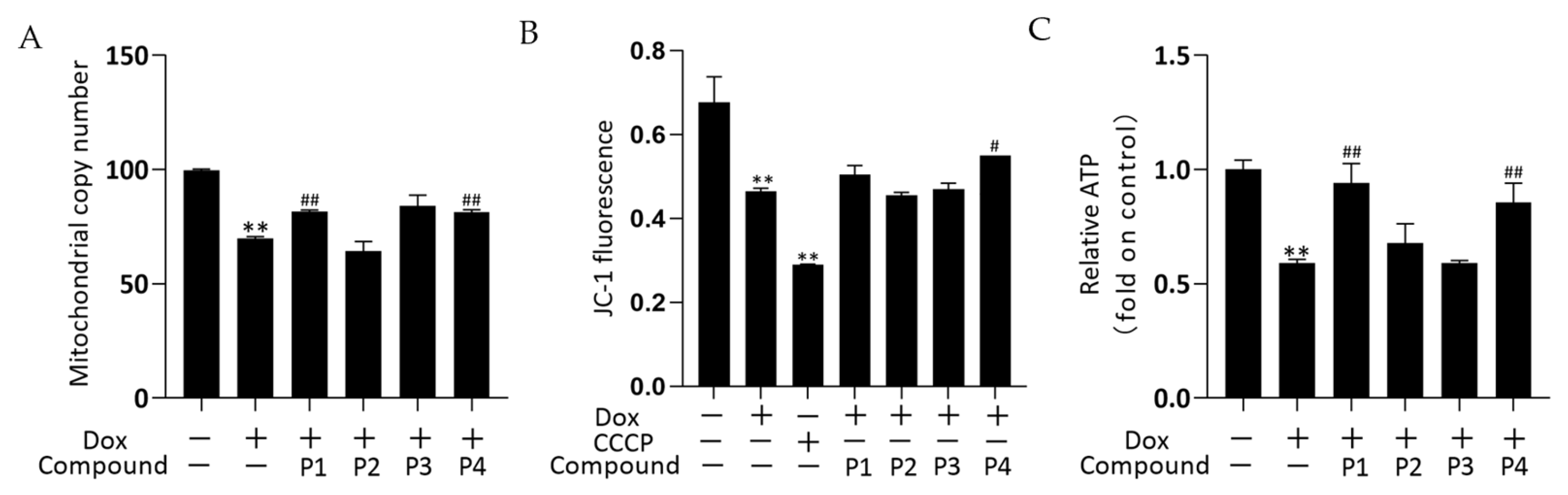
2.4. Effects of H. rhamnoides L. Phenylpropanoids on Mitochondrial-Related Assays in Zebrafish
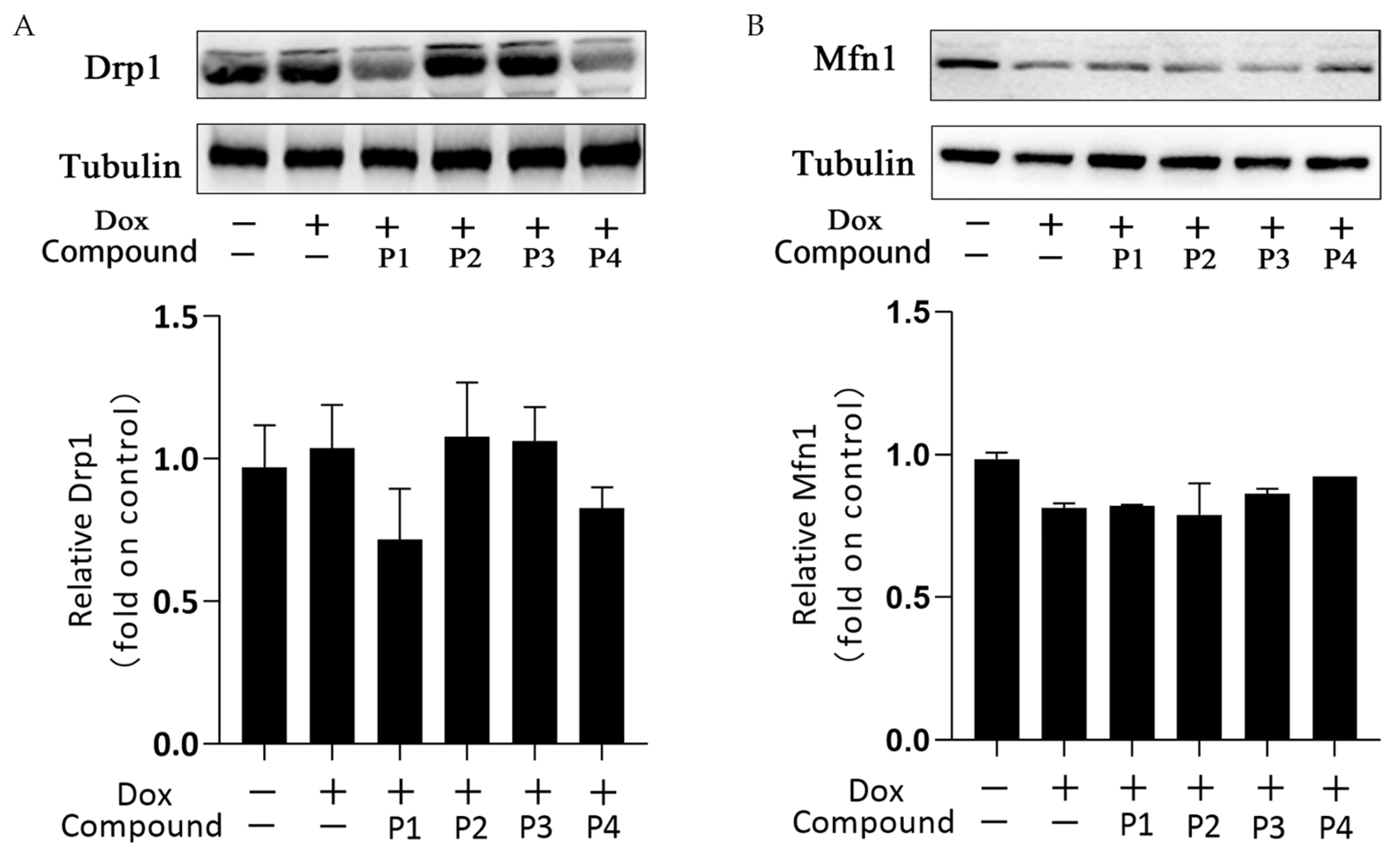
2.5. Effects of H. rhamnoides L. Phenylpropanoids on Mitochondrial Dynamics–Related Proteins
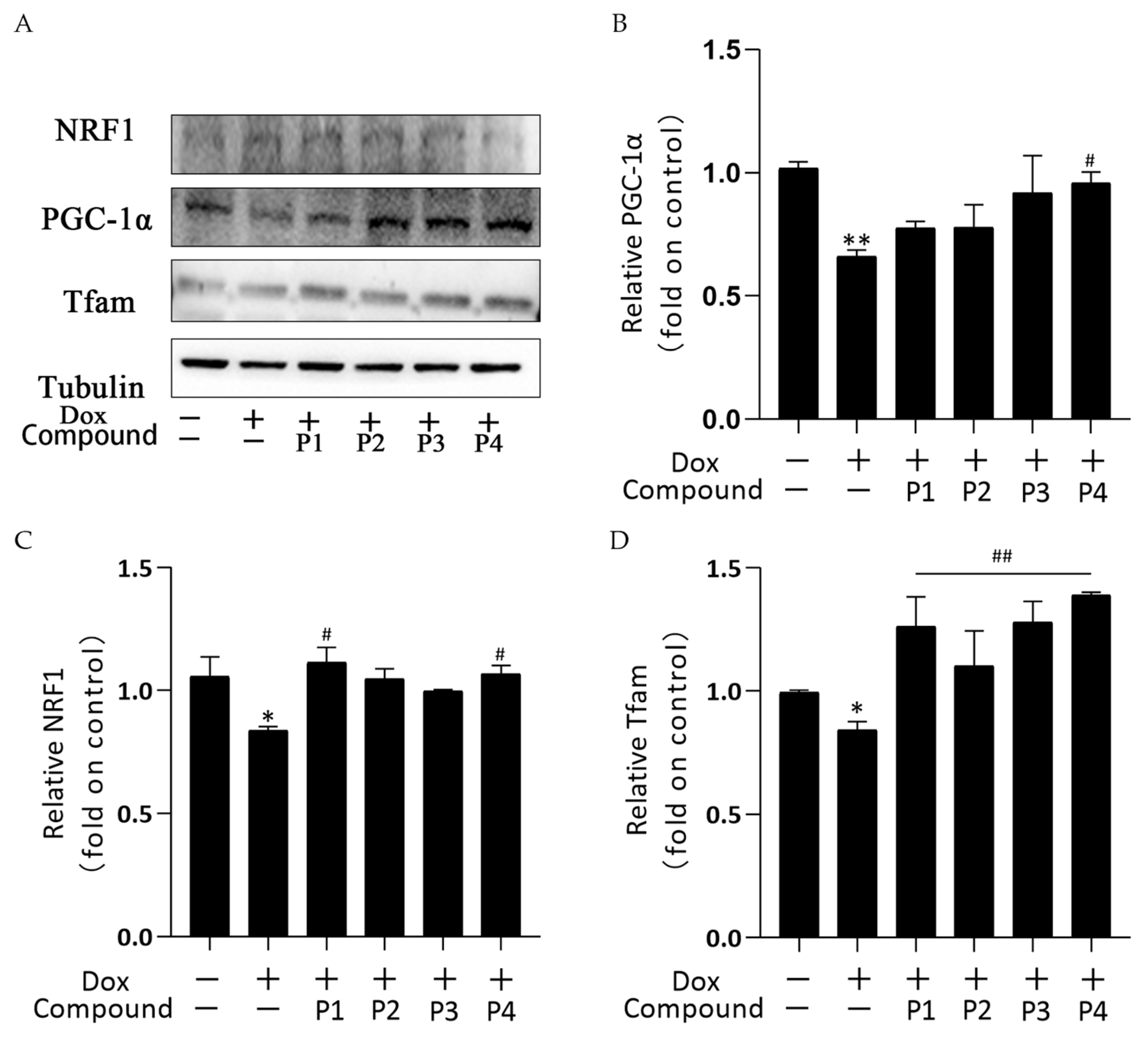
2.6. Effects of H. rhamnoides L. Phenylpropanoids on Mitochondrial Biogenesis in Zebrafish
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Zebrafish Maintenance
4.3. Zebrafish Treatment
4.4. Observation of Pericardial Morphology
4.5. Quantification of the Pericardial Area
4.6. Quantification of Cardiac Cyclization
4.7. Heart Rate Measurement
4.8. Locomotor Activity Assessment
4.9. AO Staining
4.10. Western Blotting
4.11. Assay to Determine ATP Content
4.12. Assay to Determine MMP
4.13. Mitochondrial Copy Number Assay
4.14. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Arcamone, F.; Cassinelli, G.; Fantini, G.; Grein, A.; Orezzi, P.; Pol, C.; Spalla, C. Adriamycin, 14-hydroxydaunomycin, a new antitumor antibiotic from S. peucetius var. caesius. Biotechnol. Bioeng. 1969, 11, 1101–1110. [Google Scholar] [CrossRef] [PubMed]
- van der Zanden, S.Y.; Qiao, X.; Neefjes, J. New insights into the activities and toxicities of the old anticancer drug doxorubicin. FEBS J. 2021, 288, 6095–6111. [Google Scholar] [CrossRef] [PubMed]
- Lotrionte, M.; Biondi-Zoccai, G.; Abbate, A.; Lanzetta, G.; D'Ascenzo, F.; Malavasi, V.; Peruzzi, M.; Frati, G.; Palazzoni, G. Review and Meta-Analysis of Incidence and Clinical Predictors of Anthracycline Cardiotoxicity. Am. J. Cardiol. 2013, 112, 1980–1984. [Google Scholar] [CrossRef] [PubMed]
- Lipshultz, S.E.; Colan, S.D.; Gelber, R.D.; Perez-Atayde, A.R.; Sallan, S.E.; Sanders, S.P. Late cardiac effects of doxorubicin therapy for acute lymphoblastic leukemia in childhood. N. Engl. J. Med. 1991, 324, 808–815. [Google Scholar] [CrossRef] [PubMed]
- Lipshultz, S.E.; Anderson, L.M.; Miller, T.L.; Gerschenson, M.; Stevenson, K.E.; Neuberg, D.S.; Franco, V.I.; LiButti, D.E.; Silverman, L.B.; Vrooman, L.M.; et al. Impaired mitochondrial function is abrogated by dexrazoxane in doxorubicin-treated childhood acute lymphoblastic leukemia survivors. Cancer 2016, 122, 946–953. [Google Scholar] [CrossRef] [Green Version]
- Govender, J.; Loos, B.; Marais, E.; Engelbrecht, A.M. Mitochondrial catastrophe during doxorubicin-induced cardiotoxicity: A review of the protective role of melatonin. J. Pineal Res. 2014, 57, 367–380. [Google Scholar] [CrossRef]
- He, L.; Liu, F.X.; Li, J.X. Mitochondrial Sirtuins and Doxorubicin-induced Cardiotoxicity. Cardiovasc. Toxicol. 2021, 21, 179–191. [Google Scholar] [CrossRef]
- Olas, B.; Skalski, B.; Ulanowska, K. The Anticancer Activity of Sea Buckthorn Elaeagnus rhamnoides (L.) A. Nelson. Front. Pharmacol. 2018, 9, 232. [Google Scholar] [CrossRef] [Green Version]
- Malik, S.; Goyal, S.; Ojha, S.K.; Bharti, S.; Nepali, S.; Kumari, S.; Singh, V.; Arya, D.S. Seabuckthorn Attenuates Cardiac Dysfunction and Oxidative Stress in Isoproterenol-Induced Cardiotoxicity in Rats. Int. J. Toxicol. 2011, 30, 671–680. [Google Scholar] [CrossRef]
- Masoodi, K.Z.; Wani, W.; Dar, Z.A.; Mansoor, S.; Anam-ul-Haq, S.; Farooq, I.; Hussain, K.; Wani, S.A.; Nehvi, F.A.; Ahmed, N. Sea buckthorn (Hippophae rhamnoides L.) inhibits cellular proliferation, wound healing and decreases expression of prostate specific antigen in prostate cancer cells in vitro. J. Funct. Foods 2020, 73, 104102. [Google Scholar] [CrossRef]
- Olas, B.; Skalski, B. Preparations from Various Organs of Sea Buckthorn (Elaeagnus rhamnoides (L.) A. Nelson) as Important Regulators of Hemostasis and Their Role in the Treatment and Prevention of Cardiovascular Diseases. Nutrients 2022, 14, 991. [Google Scholar] [CrossRef]
- Yue, X.F.; Shang, X.; Zhang, Z.J.; Zhang, Y.N. Phytochemical composition and antibacterial activity of the essential oils from different parts of sea buckthorn (Hippophae rhamnoides L.). J. Food Drug Anal. 2017, 25, 327–332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suchal, K.; Bhatia, J.; Malik, S.; Malhotra, R.K.; Gamad, N.; Goyal, S.; Nag, T.C.; Arya, D.S.; Ojha, S. Seabuckthorn Pulp Oil Protects against Myocardial Ischemia-Reperfusion Injury in Rats through Activation of Akt/eNOS. Front. Pharmacol. 2016, 7, 155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, W.N.; Ouyang, J.; Hu, N.; Li, G.; Wang, H.L. Protective Effect of Two Alkaloids from Hippophae rhamnoides Linn. against Doxorubicin-Induced Toxicity in H9c2 Cardiomyoblasts. Molecules 2021, 26, 1946. [Google Scholar] [CrossRef] [PubMed]
- Shen, R.; Yu, Y.C.; Lan, R.; Yu, R.; Yuan, Z.; Xia, Z.N. The cardiovascular toxicity induced by high doses of gatifloxacin and ciprofloxacin in zebrafish. Environ. Pollut. 2019, 254, 112861. [Google Scholar] [CrossRef]
- Heger, Z.; Cernei, N.; Kudr, J.; Gumulec, J.; Blazkova, I.; Zitka, O.; Eckschlager, T.; Stiborova, M.; Adam, V.; Kizek, R. A novel insight into the cardiotoxicity of antineoplastic drug doxorubicin. Int. J. Mol. Sci. 2013, 14, 21629–21646. [Google Scholar] [CrossRef] [Green Version]
- OuYang, J.; Zhou, W.N.; Li, G.; Wang, X.Y.; Ding, C.X.; Suo, Y.R.; Wang, H.L. Three New Alkaloids from Hippophae rhamnoides LINN. subsp sinensis ROUSI. Helv. Chim. Acta 2015, 98, 1287–1291. [Google Scholar] [CrossRef]
- Zhou, W.N.; Yuan, Z.Z.; Li, G.; Ouyang, J.; Suo, Y.R.; Wang, H.L. Isolation and structure determination of a new flavone glycoside from seed residues of seabuckthorn (Hippophae rhamnoides L.). Nat. Prod. Res. 2018, 32, 892–897. [Google Scholar] [CrossRef]
- Olas, B. Sea buckthorn as a source of important bioactive compounds in cardiovascular diseases. Food Chem. Toxicol. 2016, 97, 199–204. [Google Scholar] [CrossRef]
- Huang, S.R.; Huang, M.; Tian, S.N.; Meng, Z.Y.; Yan, S.; Teng, M.M.; Zhou, Z.Q.; Diao, J.L.; Zhu, W.T. Imazalil and its metabolite imazalil-M caused developmental toxicity in zebrafish (Danio rerio) embryos via cell apoptosis mediated by metabolic disorders. Pestic. Biochem. Physiol. 2022, 184, 105113. [Google Scholar] [CrossRef]
- Li, W.J.; Zhang, X.Y.; Wu, R.T.; Song, Y.H.; Xie, M.Y. Ganoderma atrum polysaccharide improves doxorubicin-induced cardiotoxicity in mice by regulation of apoptotic pathway in mitochondria. Carbohydr. Polym. 2018, 202, 581–590. [Google Scholar] [CrossRef] [PubMed]
- Wenningmann, N.; Knapp, M.; Ande, A.; Vaidya, T.R.; Ait-Oudhia, S. Insights into Doxorubicin-induced Cardiotoxicity: Molecular Mechanisms, Preventive Strategies, and Early Monitoring. Mol. Pharmacol. 2019, 96, 219–232. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.Q.; Zhen, D.; Bai, M.R.; Xuan, T.Q.; Wang, Y.; Liu, M.J.; Yu, L.J.; Bai, D.S.; Fu, D.N.; Wei, C.X. Ethanol extracts of Rhaponticum uniflorum (L.) DC flowers attenuate doxorubicin-induced cardiotoxicity via alleviating apoptosis and regulating mitochondrial dynamics in H9c2 cells. J. Ethnopharmacol. 2022, 288, 114936. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.Y.; Wang, S.Y.; Wang, L.F.; Ceylan, A.F.; Ren, J.; Zhang, Y.M. Mitophagy inhibitor liensinine suppresses doxorubicin-induced cardiotoxicity through inhibition of Drp1-mediated maladaptive mitochondrial fission. Pharmacol. Res. 2020, 157, 104846. [Google Scholar] [CrossRef]
- Suliman, H.B.; Carraway, M.S.; Ali, A.S.; Reynolds, C.M.; Welty-Wolf, K.E.; Piantadosi, C.A. The CO/HO system reverses inhibition of mitochondrial biogenesis and prevents murine doxorubicin cardiomyopathy. J. Clin. Investig. 2007, 117, 3730–3741. [Google Scholar] [CrossRef] [Green Version]
- Hao, E.K.; Mukhopadhyay, P.; Cao, Z.X.; Erdelyi, K.; Holovac, E.; Liaudet, L.; Lee, W.S.; Hasko, G.; Mechoulam, R.; Pacher, P. Cannabidiol Protects against Doxorubicin-Induced Cardiomyopathy by Modulating Mitochondrial Function and Biogenesis. Mol. Med. 2015, 21, 38–45. [Google Scholar] [CrossRef]
- Miyagawa, K.; Emoto, N.; Widyantoro, B.; Nakayama, K.; Yagi, K.; Rikitake, Y.; Suzuki, T.; Hirata, K. Attenuation of Doxorubicin-Induced Cardiomyopathy by Endothelin-Converting Enzyme-1 Ablation through Prevention of Mitochondrial Biogenesis Impairment. Hypertension 2010, 55, 738–746. [Google Scholar] [CrossRef] [Green Version]
- Duan, J.C.; Yu, Y.B.; Li, Y.; Yu, Y.; Sun, Z.W. Cardiovascular toxicity evaluation of silica nanoparticles in endothelial cells and zebrafish model. Biomaterials 2013, 34, 5853–5862. [Google Scholar] [CrossRef]
- Zhao, Y.; Han, Y.J.; Wang, Z.H.; Chen, T.R.; Qian, H.W.; He, J.; Li, J.; Han, B.; Wang, T. Rosmarinic acid protects against 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine-induced dopaminergic neurotoxicity in zebrafish embryos. Toxicol. Vitr. 2020, 65, 104823. [Google Scholar] [CrossRef]
- Li, M.; Yao, L.; Chen, H.; Ni, X.; Xu, Y.; Dong, W.J.; Fang, M.L.; Chen, D.; Xu, L.; Zhao, B.; et al. Chiral toxicity of muscone to embryonic zebrafish heart. Aquat. Toxicol. 2020, 222, 105451. [Google Scholar] [CrossRef]
- Antkiewicz, D.S.; Burns, C.G.; Carney, S.A.; Peterson, R.E.; Heideman, W. Heart malformation is an early response to TCDD in embryonic zebrafish. Toxicol. Sci. 2005, 84, 368–377. [Google Scholar] [CrossRef] [PubMed]
- Incardona, J.P.; Collier, T.K.; Scholz, N.L. Defects in cardiac function precede morphological abnormalities in fish embryos exposed to polycyclic aromatic hydrocarbons. Toxicol. Appl. Pharmacol. 2004, 196, 191–205. [Google Scholar] [CrossRef] [PubMed]
- Choi, V.W.Y.; Ng, C.Y.P.; Cheng, S.H.; Yu, K.N. alpha-Particle Irradiated Zebrafish Embryos Rescued by Bystander Unirradiated Zebrafish Embryos. Environ. Sci. Technol. 2012, 46, 226–231. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.-S.; Kim, M.-J.; Koo, T.-H.; Kim, J.-D.; Koun, S.; Ham, H.J.; Lee, Y.M.; Rhee, M.; Yeo, S.-Y.; Huh, T.-L. Histone deacetylase is required for the activation of Wnt/β-catenin signaling crucial for heart valve formation in zebrafish embryos. Biochem. Biophys. Res. Commun. 2012, 423, 140–146. [Google Scholar] [CrossRef]
- Chen, J.T.; Zhang, S.; Pan, G.X.; Lin, L.; Liu, D.Y.; Liu, Z.; Mei, S.; Zhang, L.J.; Hu, Z.H.; Chen, J.G.; et al. Modulatory effect of metformin on cardiotoxicity induced by doxorubicin via the MAPK and AMPK pathways. Life Sci. 2020, 249, 117498. [Google Scholar] [CrossRef]
- Sishi, B.J.N.; Loos, B.; van Rooyen, J.; Engelbrecht, A.M. Autophagy upregulation promotes survival and attenuates doxorubicin-induced cardiotoxicity. Biochem. Pharmacol. 2013, 85, 124–134. [Google Scholar] [CrossRef]
- Otten, A.B.C.; Kamps, R.; Lindsey, P.; Gerards, M.; Pendeville-Samain, H.; Muller, M.; van Tienen, F.H.J.; Smeets, H.J.M. Tfam Knockdown Results in Reduction of mtDNA Copy Number, OXPHOS Deficiency and Abnormalities in Zebrafish Embryos. Front. Cell Dev. Biol. 2020, 8, 381. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | Primer Sequence |
|---|---|
| Mitochondrial | Forward: 5′-CAAACACAAGCCTCGCCTGTTTAC-3′ |
| Reverse: 5′-CACTGACTTGATGGGGGAGACAGT-3′ | |
| β-Actin | Forward: 5′-ATGGATGAGGAAATCGCTGCC-3′ |
| Reverse: 5′-CTCCCTGATGTCTGGGTCGTC-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, G.; Chu, M.; Tong, Y.; Liang, Y.; Wang, S.; Ma, C.; Wang, Z.; Zhou, W. Protective Effects of Hippophae rhamnoides L. Phenylpropanoids on Doxorubicin-Induced Cardiotoxicity in Zebrafish. Molecules 2022, 27, 8858. https://doi.org/10.3390/molecules27248858
Li G, Chu M, Tong Y, Liang Y, Wang S, Ma C, Wang Z, Zhou W. Protective Effects of Hippophae rhamnoides L. Phenylpropanoids on Doxorubicin-Induced Cardiotoxicity in Zebrafish. Molecules. 2022; 27(24):8858. https://doi.org/10.3390/molecules27248858
Chicago/Turabian StyleLi, Gang, Ming Chu, Yingying Tong, Yuexin Liang, Shenghui Wang, Chengjun Ma, Zhenhua Wang, and Wenna Zhou. 2022. "Protective Effects of Hippophae rhamnoides L. Phenylpropanoids on Doxorubicin-Induced Cardiotoxicity in Zebrafish" Molecules 27, no. 24: 8858. https://doi.org/10.3390/molecules27248858
APA StyleLi, G., Chu, M., Tong, Y., Liang, Y., Wang, S., Ma, C., Wang, Z., & Zhou, W. (2022). Protective Effects of Hippophae rhamnoides L. Phenylpropanoids on Doxorubicin-Induced Cardiotoxicity in Zebrafish. Molecules, 27(24), 8858. https://doi.org/10.3390/molecules27248858