
Optimal Secretory Expression of Acetaldehyde Dehydrogenase from Issatchenkia terricola in Bacillus subtilis through a Combined Strategy




Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Plasmids
2.2. Media and Growth Conditions
2.3. Construction of a Plasmid Library of istALDH Fused with Different SPs
2.4. Construction of Plasmids Expressing the istALDH with Different Promoter
2.5. Assay and Detection of Secreted istALDH
2.6. Quantitative Real-Time PCR Analysis of istALDH Expression in Recombinant B. subtilis
3. Results and Discussion
3.1. Heterologous Production of Secreted istALDH in B. subtilis Strain RIK1285
3.2. Effect of the Signal Peptides on the Secretion of istALDH
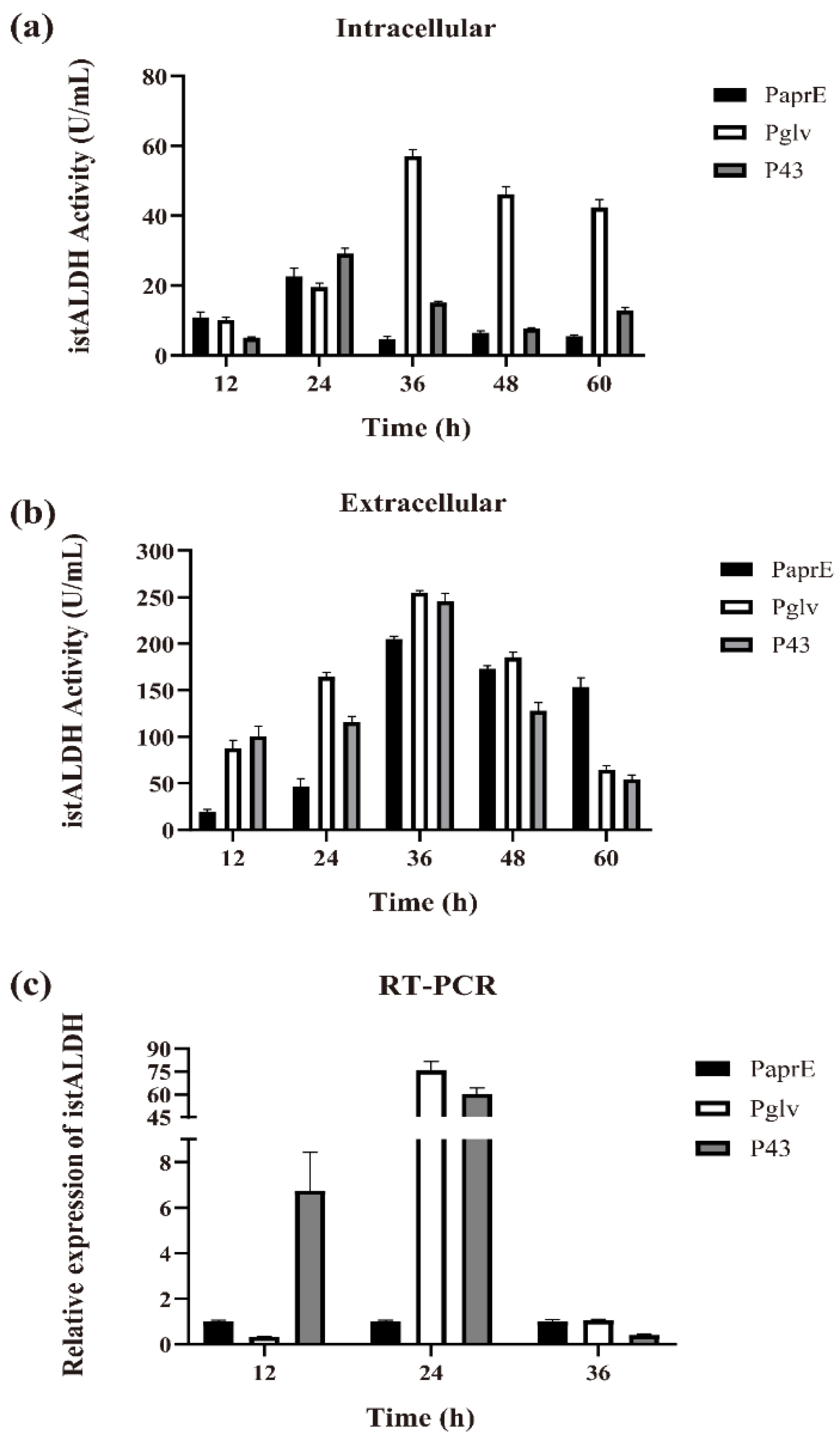
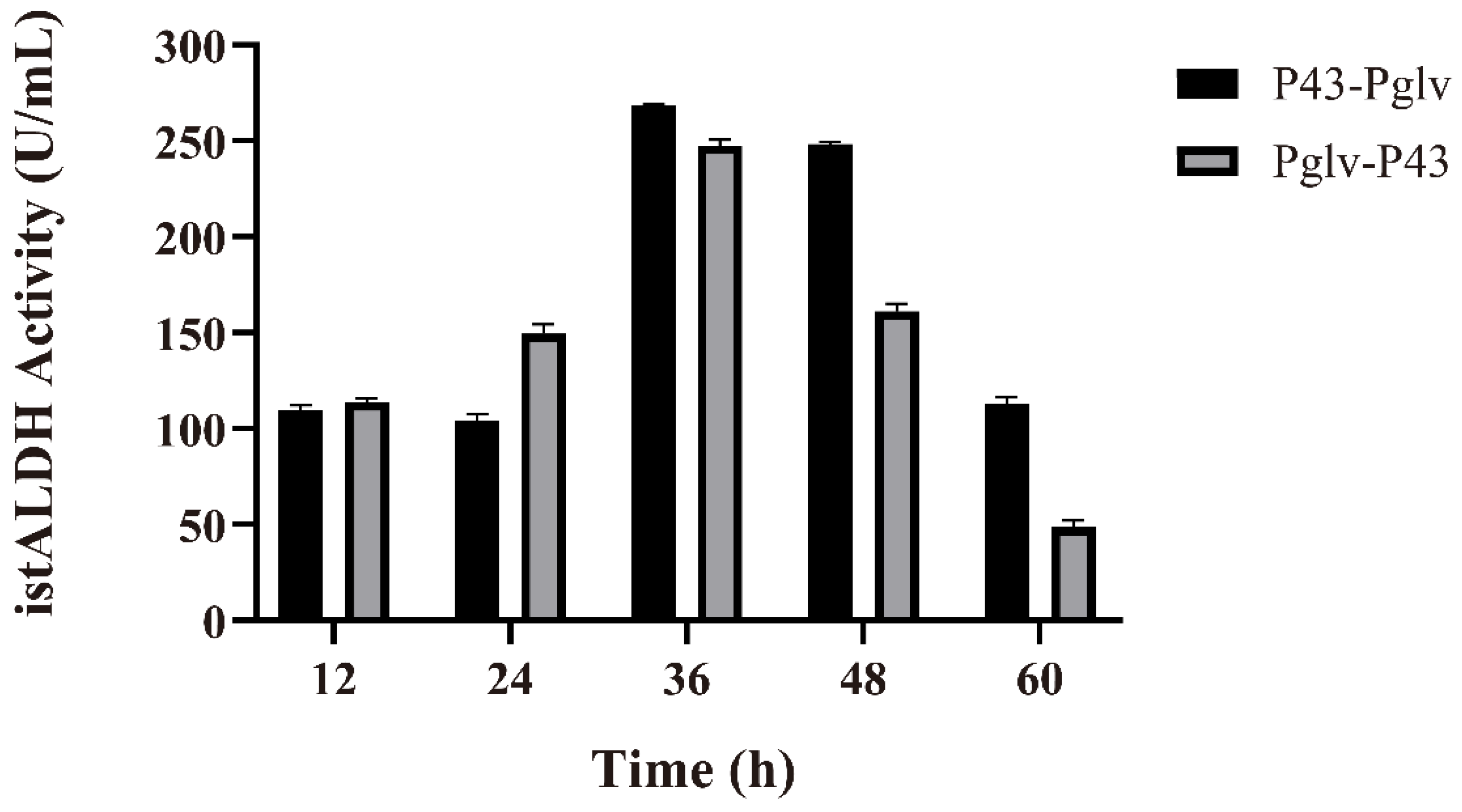
3.3. Effect of the Promoter on Secretion of istALDH
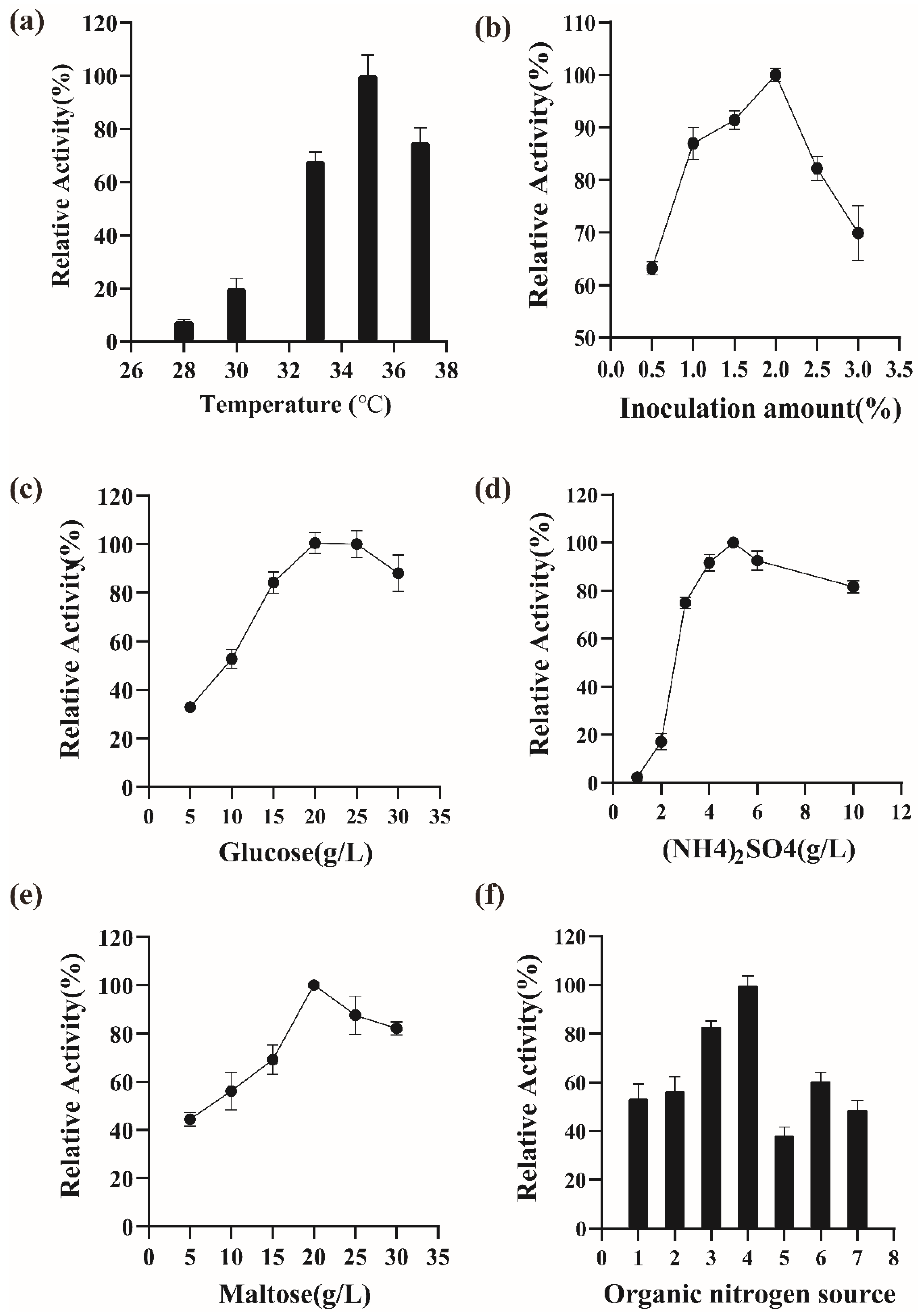
3.4. Effect on the Fermentation Conditions on the Secretion of istALDH
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sophos, N.A.; Pappa, A.; Ziegler, T.L.; Vasiliou, V. Aldehyde dehydrogenase gene superfamily: The 2000 update. Chem. Biol. Interact. 2003, s143–s144, 323–337. [Google Scholar] [CrossRef]
- Gonzálezsegura, L.; Riverosrosas, H.; Juliánsánchez, A.; Muñozclares, R.A. Residues that influence coenzyme preference in the aldehyde dehydrogenases. Chem. Biol. Interact. 2015, 234, 59–74. [Google Scholar] [CrossRef]
- Dellarco, V.L. A mutagenicity assessment of acetaldehyde. Mutat. Res. Rev. Genet. Toxicol. 1988, 195, 1–20. [Google Scholar] [CrossRef]
- Maffei, F.; Fimognari, C.; Castelli, E.; Stefanini, G.F.; Forti, G.C.; Hrelia, P. Increased cytogenetic damage detected by FISH analysis on micronuclei in peripheral lymphocytes from alcoholics. Mutagenesis 2000, 15, 517–523. [Google Scholar] [CrossRef] [Green Version]
- Lachenmeier, D.W.; Sohnius, E.M. The role of acetaldehyde outside ethanol metabolism in the carcinogenicity of alcoholic beverages: Evidence from a large chemical survey. Food Chem. Toxicol. 2008, 46, 2903–2911. [Google Scholar] [CrossRef] [PubMed]
- Yao, Z.; Lu, F.; Bie, X.; Lu, Z. Gene cloning, expression, and characterization of a novel acetaldehyde dehydrogenase from Issatchenkia terricola strain XJ-2. Appl. Microbiol. Biotechnol. 2012, 93, 1999–2009. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Zhu, X.; Zhang, C.; Lu, F.; Lu, Z.; Lu, Y. Co-expression of alcohol dehydrogenase and aldehyde dehydrogenase in Bacillus subtilis for alcohol detoxification. Food Chem. Toxicol. 2020, 135, 110890. [Google Scholar] [CrossRef] [PubMed]
- Schädlich, L.; Senger, T.; Kirschning, C.J.; Müller, M.; Gissmann, L. Refining HPV 16 L1 purification from E. coli: Reducing endotoxin contaminations and their impact on immunogenicity. Vaccine 2009, 27, 1511–1522. [Google Scholar] [CrossRef] [PubMed]
- Kai, H.; Xiao, G.; Bo, J.; Li, S. Construction of a food-grade arginase expression system and its application in L-ornithine production with whole cell biocatalyst. Process Biochem. 2018, 73, 94–101. [Google Scholar] [CrossRef]
- Vavrová, L.; Muchová, K.; Barák, I.; Felice, M.D.; Fouet, A.; Barák, I.; Cutting, S.; Ricca, E. Comparison of different Bacillus subtilis expression systems. Res. Microbiol. 2010, 161, 791–797. [Google Scholar] [CrossRef] [PubMed]
- Harwood, C.R.; Cranenburgh, R. Bacillus protein secretion: An unfolding story. Trends Microbiol. 2008, 16, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Miao, Y.; Hu, L.-p.; Kai, W.; Zhu, R. Immunization of mice against alpha, beta, and epsilon toxins of Clostridium perfringens using recombinant rCpa-b-x expressed by Bacillus subtilis. Mol. Immunol. 2020, 123, 88–96. [Google Scholar] [CrossRef] [PubMed]
- Dong, H.; Zhang, D. Current development in genetic engineering strategies of Bacillus species. Microb. Cell Factories 2014, 13, 63. [Google Scholar] [CrossRef]
- Zhou, C.; Ye, B.; Cheng, S.; Zhao, L.; Liu, Y.; Jiang, J.; Yan, X. Promoter engineering enables overproduction of foreign proteins from a single copy expression cassette in Bacillus subtilis. Microb. Cell Factories 2019, 18, 111. [Google Scholar] [CrossRef] [Green Version]
- Mao, S.; Zhang, Z.; Ma, X.; Tian, H.; Liu, Y. Efficient secretion expression of phospholipase D in Bacillus subtilis and its application in synthesis of phosphatidylserine by enzyme immobilization. Int. J. Biol. Macromol. 2021, 169, 282–289. [Google Scholar] [CrossRef]
- Peng, C.; Shi, C.; Cao, X.; Li, Y.; Liu, F.; Lu, F. Factors influencing recombinant protein secretion efficiency in Gram-positive bacteria: Signal peptide and beyond. Front. Bioeng. Biotechnol. 2019, 7, 139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, Y.; Liu, S.; Jiao, Y.; Gao, H.; Wang, M.; Du, G.; Chen, J. Enhanced extracellular production of L-asparaginase from Bacillus subtilis 168 by B. subtilis WB600 through a combined strategy. Appl. Microbiol. Biotechnol. 2017, 101, 1509–1520. [Google Scholar] [CrossRef]
- Su, L.; Li, Y.; Wu, J. Efficient secretory expression of Bacillus stearothermophilus α/β-cyclodextrin glycosyltransferase in Bacillus subtilis. J. Biotechnol. 2021, 331, 74–82. [Google Scholar] [CrossRef] [PubMed]
- Guan, C.; Cui, W.; Cheng, J.; Rui, L.; Liu, Z.; Li, Z.; Zhou, Z. Construction of a highly active secretory expression system via an engineered dual promoter and a highly efficient signal peptide in Bacillus subtilis. New Biotechnol. 2016, 33, 372–379. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Xie, M.; Yang, Y.; Liu, T.; Liqin, D.U.; Liang, Z.; Wei, Y. High Level Secretion Expression of Maltogenicα-amylase from Saccharomonospora viridis in Bacillus subtilis. Guangxi Sci. 2016, 23, 12–18. [Google Scholar]
- Gao, L.; Han, J.; Liu, H.; Qu, X.; Lu, Z.; Bie, X. Plipastatin and surfactin coproduction by Bacillus subtilis pB2-L and their effects on microorganisms. Antonie Van Leeuwenhoek 2017, 110, 1007–1018. [Google Scholar] [CrossRef] [PubMed]
- Mayer, K.M.; Arnold, F.H. A colorimetric assay to quantify dehydrogenase activity in crude cell lysates. J. Biomol. Screen. 2002, 7, 135. [Google Scholar] [CrossRef] [Green Version]
- Bostian, K.A.; Betts, G.F. Kinetics and reaction mechanism of potassium-activated aldehyde dehydrogenase from Saccharomyces cerevisiae. Biochem. J. 1978, 173, 787–798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jing, L.; Yunbin, L.; Mingtong, L.; Jing, S.; Zhenghua, H.; Fengxia, L.; Zhaoxin, L. Alleviating acute alcoholic liver injury in mice with Bacillus subtilis co-expressing alcohol dehydrogenase and acetaldehyde dehydrogenase. J. Funct. Foods 2018, 49, 342–350. [Google Scholar] [CrossRef]
- Zanen, G.; Houben, E.N.G.; Meima, R.; Tjalsma, H.; Jongbloed, J.D.; Westers, H.; Oudega, B.; Luirink, J.; van Dijl, J.M.; Quax, W.J. Signal peptide hydrophobicity is critical for early stages in protein export by Bacillus subtilis. FEBS J. 2005, 272, 4617. [Google Scholar] [CrossRef]
- Ismail, N.F.; Hamdan, S.; Mahadi, N.M.; Ama, M.; Rabu, A.; Fda, B.; Klappa, P.; Illias, R.M. A mutant l-asparaginase II signal peptide improves the secretion of recombinant cyclodextrin glucanotransferase and the viability of Escherichia coli. Biotechnol. Lett. 2011, 33, 999–1005. [Google Scholar] [CrossRef] [PubMed]
- Caspers, M.; Brockmeier, U.; Degering, C.; Eggert, T.; Freudl, R. Improvement of Sec-dependent secretion of a heterologous model protein in Bacillus subtilis by saturation mutagenesis of the N-domain of the AmyE signal peptide. Appl. Microbiol. Biotechnol. 2010, 86, 1877–1885. [Google Scholar] [CrossRef]
- Inouye, M.; Halegoua, S.; Beckwith, J. Secretion and membrane Localization of Proteins in Escherichia Coli. CRC Crit. Rev. Biochem. 1980, 7, 339–371. [Google Scholar] [CrossRef]
- Nielsen, H. Predicting Secretory Proteins with SignalP. Methods Mol. Biol. 2017, 1611, 59–73. [Google Scholar]
- Wang, G.; Dong, Y.; Chen, H.; Zhang, H.; Song, Y.; Chen, W. High incidence of disorder-promoting amino acids in the amino terminus of mature proteins in Bacillus subtilis. Am. J. Biochem. Biotechnol. 2012, 7, 172–178. [Google Scholar]
- Miao, H.; Jiang, R.; Han, N.; Ma, Y.; Wu, Q.; Mu, Y.; Huang, Z. Enhanced extracellular expression of α-Amylase DL3-4-1 in Bacillus subtilis via systematic screening of optimal signal peptides. Process Biochem. 2021, 108, 176–184. [Google Scholar] [CrossRef]
- Ming-Ming, Y.; Wei-Wei, Z.; Xi-Feng, Z.; Pei-Lin, C. Construction and characterization of a novel maltose inducible expression vector in Bacillus subtilis. Biotechnol. Lett. 2006, 28, 1713–1718. [Google Scholar] [CrossRef]
- Yang, M.M.; Zhang, W.W.; Chen, Y.L.; Gong, Y.S. Development of a Bacillus subtilis expression system using the improved P glv promoter. Microb. Cell Factories 2010, 9, 55. [Google Scholar]
- Kim, J.H.; Hwang, B.Y.; Roh, J.W.; Lee, J.K.; Kim, K.; Wong, S.L.; Yun, H.D.; Lee, S.G.; Kim, B.G. Camparison of PaprE, PamyE, and P43 promoter strength for β-galactosidase and staphylokinase expression in Bacillus subtilis. Biotechnol. Bioprocess Eng. 2008, 13, 313. [Google Scholar] [CrossRef]
- Kang, H.K.; Jang, J.H.; Shim, J.H.; Park, J.T.; Kim, Y.W.; Park, K.H. Efficient constitutive expression of thermostable 4-α-glucanotransferase in Bacillus subtilis using dual promoters. World J. Microbiol. Biotechnol. 2010, 26, 1915–1918. [Google Scholar] [CrossRef]
- Kang, Z.; Su, L.; Duan, X.; Liu, L.; Jing, W. High-level extracellular protein production in Bacillus subtilis using an optimized dual-promoter expression system. Microb. Cell Factories 2017, 16, 32. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains/Plasmids | Characteristics * | Source |
|---|---|---|
| Strains | ||
| E.coli DH5α | deoR endA1 gyrA96 hsdR17 (rk-mk+)recA1 relA1 supE44 thi-1 Δ(lacZYA-argF)U169 Φ80lacZ ΔM15F-λ- | Vazyme Biotech |
| B. subtilis RIK1285 | Marburg 168 derivative; trpC2, lys1, aprEΔ3, nprR2, nprE18 | Takara |
| B. subtilis BS000 | derived from B. subtilis 168, amyE::promoter-signal peptide | laboratory stock |
| BS-1 | RIK1285 harboring pBE/istALDH | current study |
| BS-2 | RIK1285 harboring pBE/istALDH-1 | current study |
| BS-3 | RIK1285 harboring pPglv/istALDH | current study |
| BS-4 | RIK1285 harboring pP43/istALDH | current study |
| BS-5 | RIK1285 harboring p P43-Pglv /ist-ALDH | current study |
| BS-6 | RIK1285 harboring p Pglv-P43/ist-ALDH | current study |
| Plasmid | ||
| pMD19T | TA clon vector, Amp+ | Takara |
| pHCMC04-pPglv | Pglv promoter | [20] |
| pPSQ | P43 promoter | [21] |
| pET32a/istALDH | istALDH | [6] |
| pBE | Kan+, Amp+, PaprE, SPaprE | Takara |
| pBE/istALDH | Kan+, Amp+, PaprE, SPaprE, istALDH | current study |
| pBE/istALDH-1 | Kan+, Amp+, PaprE, SPyqzG, istALDH | current study |
| pPglv/istALDH | Kan+, Amp+, Pglv, SPyqzG, istALDH | current study |
| pP43/istALDH | Kan+, Amp+, P43, SPyqzG, istALDH | current study |
| pP43-Pglv/istALDH | Kan+, Amp+, P43-Pglv, SPyqzG, istALDH | current study |
| pPglv-P43/istALDH | Kan+, Amp+, Pglv-P43, SPyqzG, istALDH | current study |
| Primer | Sequence (5′ to 3′) |
|---|---|
| P1 | GAGCTCCTTAGAACTGCAACTAGAAC |
| P2 | GGATCCTTGTGGGCCATCGTTAATGGC |
| P3 | CGCGTCCCTCTCCTTTTGCTTAAGTTCAGAGTAG |
| P4 | GGCCGGTGCACATATGGAGCTCCTTAGAACTGC |
| P5 | GCCGTCTGTACGTTCCTAAACTAGTGGCATGTATCCGAATC |
| P6 | GTTTGATCATCATACGCGTACGACCTCCTTGATAACG |
| P7 | ATGATGATCAAACAATGTGTGATTTGTC |
| P8 | TTAGGAACGTACAGACGGCTTAAAAG |
| P9 | CTTTTAAGCCGTCTGTACGTTCCTAAACTAGTATATTCCTTTTGATAGGTGG |
| P10 | GACAAATCACACATTGTTTGATCATCATACGCGTATTCCTCTCTTACCTATAATG |
| P11 | GCCGTCTGTACGTTCCTAAACTAGTATATTCCTTTTGATAGGTGGTATG |
| P12 | CTTTTGTACGATTCGGATACATGCCACTAGTATTCCTCTCTTACCTATA |
| P13 | GGCATGTATCCGAATCGTACAAAAG |
| P14 | ACTAGTTTAGGAACGTACAGACGGCTTAAAAG |
| P15 | CTTTTAAGCCGTCTGTACGTTCCTAAACTAGTGGCATGTATCCGAATC |
| P16 | CCACCTATCAAAAGGAATATACTAGTACGACCTCCTTGATAACG |
| ALDH-RT-F | CAGAGGTGAGCACTTGATGAAAC |
| ALDH-RT-F | CAGCCCAACCAGCACAATACTTA |
| 16S rRNA-F | ACGGGAGGCAGCAGTAGGG |
| 16S rRNA-R | ACGGGAGGCAGCAGTAGGG |
| Signal Peptide | Protein Sequence | Len | D-Score a | Charge b | Hydrophilicity c (%) | Activity of istALDH d (U/mL) |
|---|---|---|---|---|---|---|
| yqzG | MMIKQCVICLSLLVFGTTAAHA | 22 | 0.716 | 1 | 63.64 | 204.85 ± 3.31 |
| wapA | MKKRKRRNFKRFIAAFLVLALMISLVPADVLA | 32 | 0.707 | 7 | 65.63 | 184.81 ± 3.59 |
| yngK | MKVCQKSIVRFLVSLIIGTFVISVPFMANA | 30 | 0.677 | 3 | 66.67 | 160.76 ± 3.86 |
| xynA | MFKFKKNFLVGLSAALMSISLFSATASA | 28 | 0.757 | 1 | 64.29 | 147.84 ± 2.90 |
| pbp | MKKSIKLYVAVLLLFVVASVPYMHQAALA | 29 | 0.672 | 3 | 68.97 | 140.20 ± 3.52 |
| aprE | MRSKKLWISLLFALTLIFTMAFSNMSVQA | 29 | 0.644 | 3 | 62.07 | 113.71 ± 3.62 |
| g/L | 1 | 2 | 3 | 4 | 5 | 6 | 7 |
|---|---|---|---|---|---|---|---|
| Peptone | 0 | 10 | 15 | 20 | 25 | 30 | 40 |
| Yeast extract | 40 | 30 | 25 | 20 | 15 | 10 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, J.; Zhao, Y.; Cheng, Y.; Hu, R.; Fang, Y.; Lyu, M.; Wang, S.; Lu, Z. Optimal Secretory Expression of Acetaldehyde Dehydrogenase from Issatchenkia terricola in Bacillus subtilis through a Combined Strategy. Molecules 2022, 27, 747. https://doi.org/10.3390/molecules27030747
Lu J, Zhao Y, Cheng Y, Hu R, Fang Y, Lyu M, Wang S, Lu Z. Optimal Secretory Expression of Acetaldehyde Dehydrogenase from Issatchenkia terricola in Bacillus subtilis through a Combined Strategy. Molecules. 2022; 27(3):747. https://doi.org/10.3390/molecules27030747
Chicago/Turabian StyleLu, Jing, Yu Zhao, Yu Cheng, Rong Hu, Yaowei Fang, MingSheng Lyu, Shujun Wang, and Zhaoxin Lu. 2022. "Optimal Secretory Expression of Acetaldehyde Dehydrogenase from Issatchenkia terricola in Bacillus subtilis through a Combined Strategy" Molecules 27, no. 3: 747. https://doi.org/10.3390/molecules27030747
APA StyleLu, J., Zhao, Y., Cheng, Y., Hu, R., Fang, Y., Lyu, M., Wang, S., & Lu, Z. (2022). Optimal Secretory Expression of Acetaldehyde Dehydrogenase from Issatchenkia terricola in Bacillus subtilis through a Combined Strategy. Molecules, 27(3), 747. https://doi.org/10.3390/molecules27030747