
Rationalisation of Antifungal Properties of α-Helical Pore-Forming Peptide, Mastoparan B

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Peptides and Cell Lines
2.2. Circular Dichroism (CD) Spectropolarimetry
2.3. Zeta-Potential (ζ) Measurements
2.4. Calculation of Physicochemical Parameters
2.5. Determination of the Minimum Inhibitory Concentrations (MICs)
2.6. Mammalian Cell Viability Assays
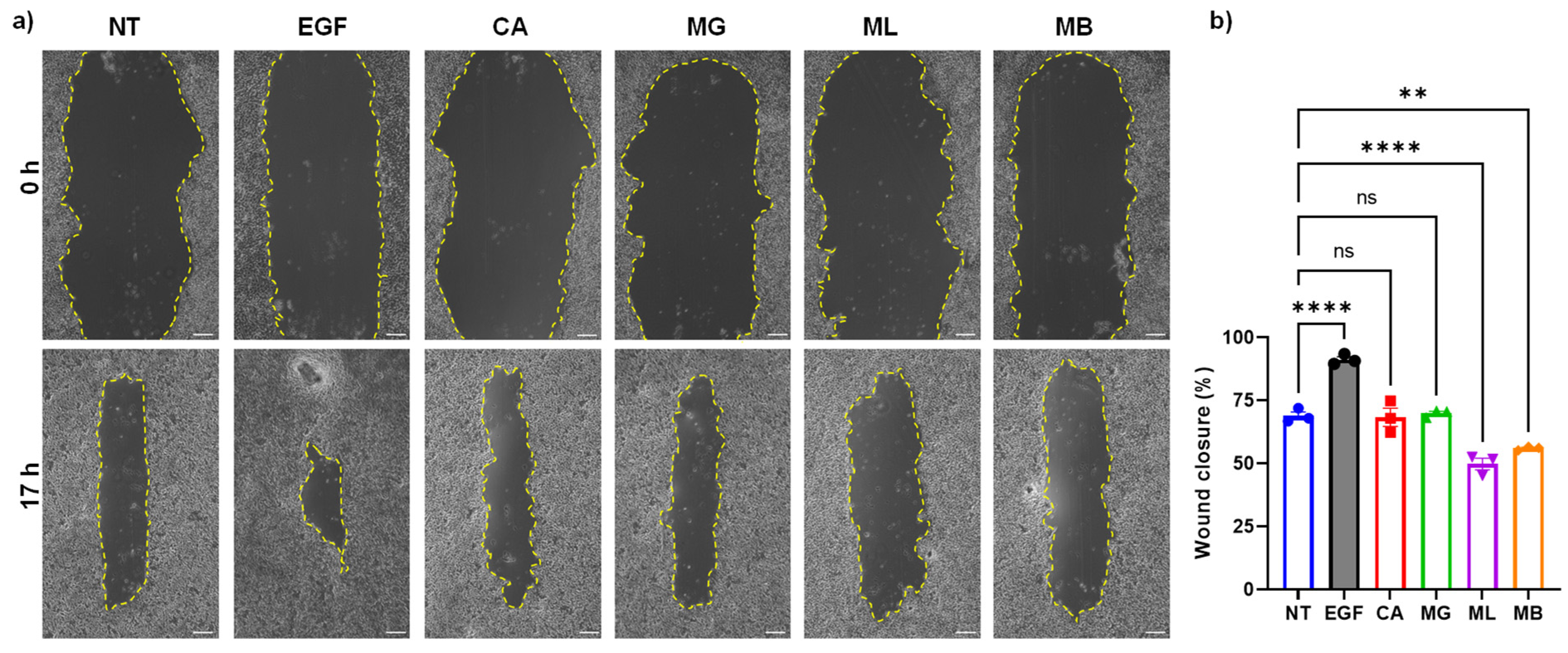
2.7. Cell Migration Assay
2.8. Sytox™ Green Uptake Assay
2.9. Time–Kill Kinetics Studies (TKS)
3. Results
3.1. Physicochemical Properties of Pore-Forming Peptides
3.2. Antifungal Activity of Pore-Forming Peptides
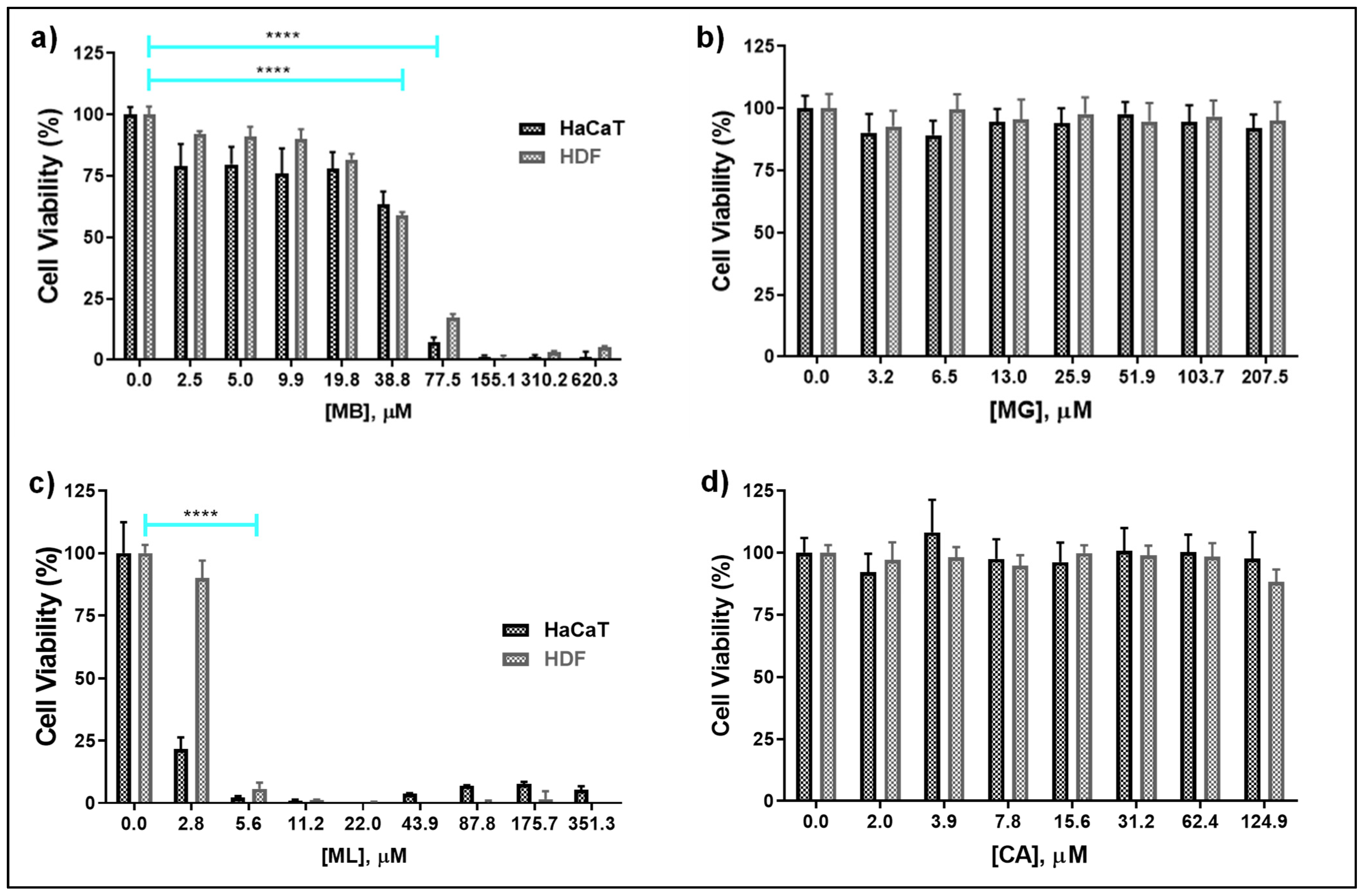
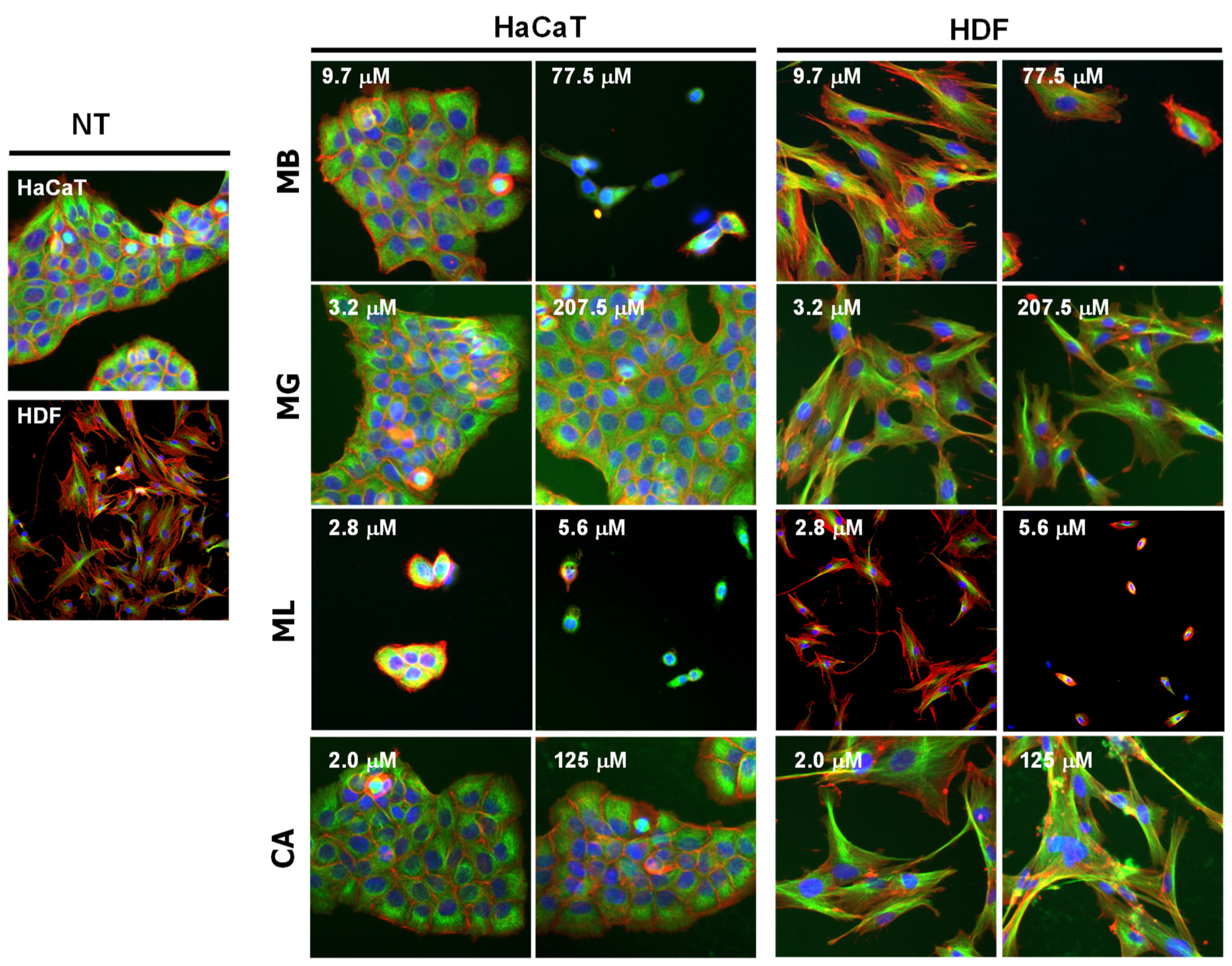
3.3. Host Cell Cytocompatibility of Pore-Forming Peptides
3.4. Antifungal Activity of MB in the Presence of Mono and Divalent Cations
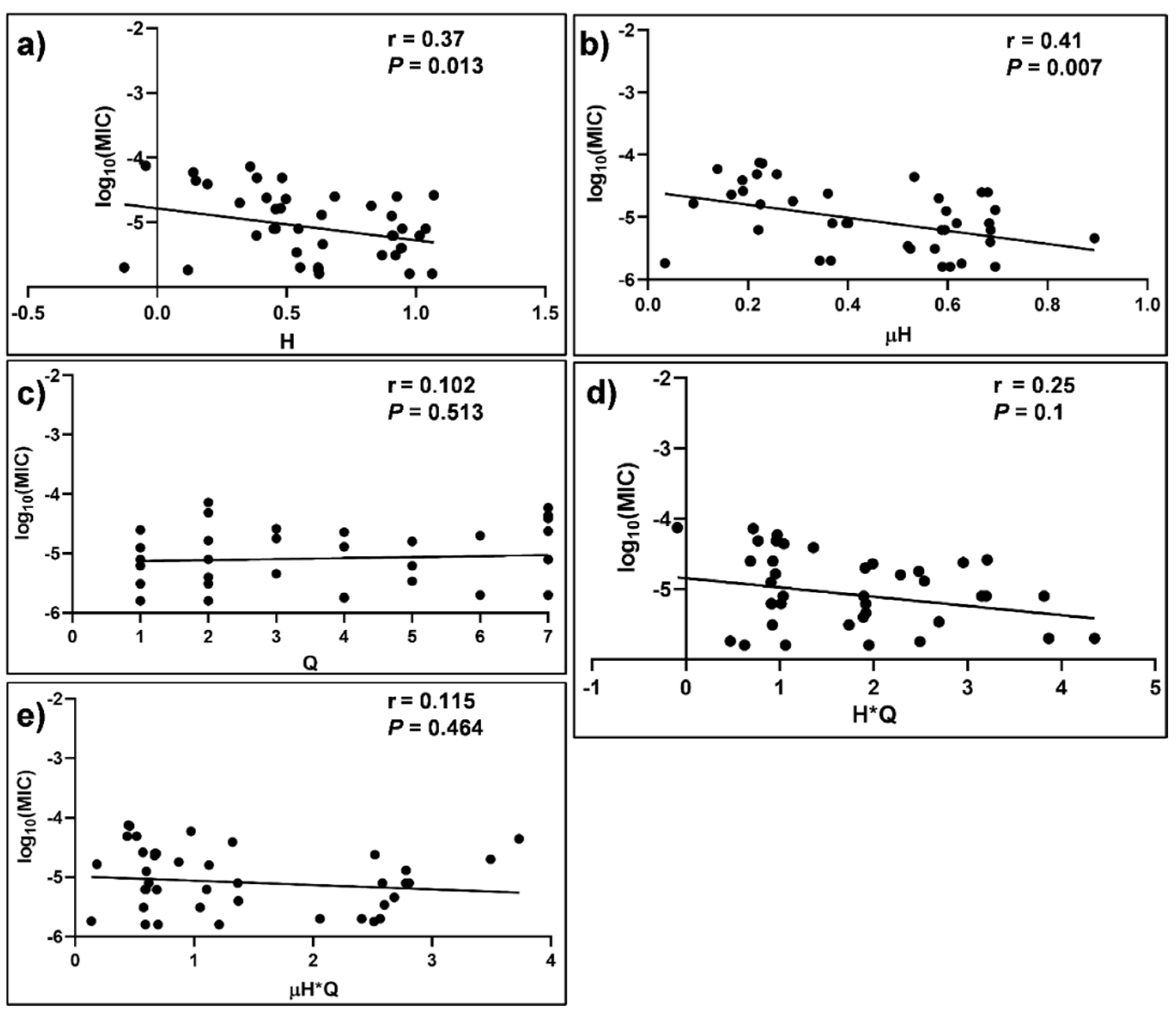
3.5. Correlation of Physicochemical Properties of MB with Anti-C. glabrata Activity
4. Discussion and Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Pappas, P.G.; Lionakis, M.S.; Arendrup, M.C.; Ostrosky-Zeichner, L.; Kullberg, B.J. Invasive candidiasis. Nat. Rev. Dis. Primers 2018, 4, 18026. [Google Scholar] [CrossRef] [PubMed]
- Kullberg, B.J.; Arendrup, M.C. Invasive candidiasis. N. Engl. J. Med. 2015, 373, 1445–1456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whaley, S.G.; Berkow, E.L.; Rybak, J.M.; Nishimoto, A.T.; Barker, K.S.; Rogers, P.D. Azole antifungal resistance in Candida albicans and emerging non-albicans Candida species. Front. Microbiol. 2017, 7, 2173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papon, N.; Courdavault, V.; Clastre, M.; Bennett, R.J. Emerging and emerged pathogenic Candida species: Beyond the Candida albicans paradigm. PLoS Pathog. 2013, 9, e1003550. [Google Scholar] [CrossRef] [Green Version]
- McCarty, T.P.; Pappas, P.G. Invasive candidiasis. Infect. Dis. Clin. N. Am. 2016, 30, 103–124. [Google Scholar] [CrossRef] [PubMed]
- Wisplinghoff, H.; Bischoff, T.; Tallent, S.M.; Seifert, H.; Wenzel, R.P.; Edmond, M.B. Nosocomial bloodstream infections in US hospitals: Analysis of 24,179 cases from a prospective nationwide surveillance study. Clin. Infect. Dis. 2004, 39, 309–317. [Google Scholar] [CrossRef] [Green Version]
- Pristov, K.E.; Ghannoum, M.A. Resistance of Candida to azoles and echinocandins worldwide. Clin. Microbiol. Infect. 2019, 25, 792–798. [Google Scholar] [CrossRef]
- Ostrosky-Zeichner, L.; Casadevall, A.; Galgiani, J.N.; Odds, F.C.; Rex, J.H. An insight into the antifungal pipeline: Selected new molecules and beyond. Nat. Rev. Drug. Discov. 2010, 9, 719–727. [Google Scholar] [CrossRef]
- Ciociola, T.; Giovati, L.; Conti, S.; Magliani, W.; Santinoli, C.; Polonelli, L. Natural and synthetic peptides with antifungal activity. Future Med. Chem. 2016, 8, 1413–1433. [Google Scholar] [CrossRef]
- Wimley, W.C.; Hristova, K. Antimicrobial peptides: Successes, challenges and unanswered questions. J. Membr Biol. 2011, 239, 27–34. [Google Scholar] [CrossRef] [Green Version]
- Haney, E.F.; Straus, S.K.; Hancock, R.E.W. Reassessing the host defense peptide landscape. Front. Chem. 2019, 7, 43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duncan, V.M.S.; O’Neil, D.A. Commercialization of antifungal peptides. Fungal Biol. Rev. 2013, 26, 156–165. [Google Scholar] [CrossRef]
- Buda De Cesare, G.; Cristy, S.A.; Garsin, D.A.; Lorenz, M.C. Antimicrobial peptides: A new frontier in antifungal therapy. mBio. 2020, 11, e02123-20. [Google Scholar] [CrossRef]
- Matsuzaki, K. Control of cell selectivity of antimicrobial peptides. Biochim. Biophys Acta 2009, 1788, 1687–1692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashby, M.; Petkova, A.; Hilpert, K. Cationic antimicrobial peptides as potential new therapeutic agents in neonates and children: A review. Curr. Opin. Infect. Dis. 2014, 27, 258–267. [Google Scholar] [CrossRef]
- Struyfs, C.; Cammue, B.P.A.; Thevissen, K. Membrane-interacting antifungal peptides. Front. Cell Dev. Biol. 2021, 9, 649875. [Google Scholar] [CrossRef]
- McMillan, K.A.M.; Coombs, M.R.P. Review: Examining the natural role of amphibian antimicrobial peptide magainin. Molecules 2020, 25, 5436. [Google Scholar] [CrossRef]
- Dutta, P.; Sahu, R.K.; Dey, T.; Lahkar, M.D.; Manna, P.; Kalita, J. Beneficial role of insect-derived bioactive components against inflammation and its associated complications (colitis and arthritis) and cancer. Chem. Biol. Interact. 2019, 313, 108824. [Google Scholar] [CrossRef]
- Smits, J.P.H.; Niehues, H.; Rikken, G.; van Vlijmen-Willems, I.M.J.J.; van de Zande, G.W.H.J.F.; Zeeuwen, P.L.J.M.; Schalkwijk, J.; van den Bogaard, E.H. Immortalized N/TERT keratinocytes as an alternative cell source in 3D human epidermal models. Sci. Rep. 2017, 7, 11838. [Google Scholar] [CrossRef]
- Lakshminarayanan, R.; Fan, D.; Du, C.; Moradian-Oldak, J. The role of secondary structure in the entropically driven amelogenin self-assembly. Biophys, J. 2007, 93, 3664–3674. [Google Scholar] [CrossRef] [Green Version]
- Gautier, R.; Douguet, D.; Antonny, B.; Drin, G. HELIQUEST: A web server to screen sequences with specific alpha-helical properties. Bioinformatics 2008, 24, 2101–2102. [Google Scholar] [CrossRef]
- Giangaspero, A.; Sandri, L.; Tossi, A. Amphipathic alpha helical antimicrobial peptides. Eur. J. Biochem. 2001, 268, 5589–5600. [Google Scholar] [CrossRef] [PubMed]
- Torres, M.D.T.; Pedron, C.N.; Higashikuni, Y.; Kramer, R.M.; Cardoso, M.H.; Oshiro, K.G.N.; Franco, O.L.; Silva, P.I., Jr.; Silva, F.D.; Oliveira, V.X., Jr.; et al. Structure-function-guided exploration of the antimicrobial peptide polybia-CP identifies activity determinants and generates synthetic therapeutic candidates. Commun. Biol. 2018, 1, 221. [Google Scholar] [CrossRef] [Green Version]
- Mojsoska, B.; Jenssen, H. Peptides and peptidomimetics for antimicrobial drug design. Pharmaceuticals (Basel) 2015, 8, 366–415. [Google Scholar] [CrossRef] [PubMed]
- Porto, W.F.; Irazazabal, L.; Alves, E.S.F.; Ribeiro, S.M.; Matos, C.O.; Pires, A.S.; Fensterseifer, I.C.M.; Miranda, V.J.; Haney, E.F.; Humblot, V.; et al. In silico optimization of a guava antimicrobial peptide enables combinatorial exploration for peptide design. Nat. Commun. 2018, 9, 1490. [Google Scholar] [CrossRef] [PubMed]
- Torres, M.T.; de la Fuente-Nunez, C. Toward computer-made artificial antibiotics. Curr. Opin. Microbiol. 2019, 51, 30–38. [Google Scholar] [CrossRef]
- Yount, N.Y.; Weaver, D.C.; Lee, E.Y.; Lee, M.W.; Wang, H.; Chan, L.C.; Wong, G.C.L.; Yeaman, M.R. Unifying structural signature of eukaryotic alpha-helical host defense peptides. Proc. Natl. Acad. Sci. USA. 2019, 116, 6944–6953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfaller, M.A.; Diekema, D.J. Epidemiology of invasive candidiasis: A persistent public health problem. Clin. Microbiol. Rev. 2007, 20, 133–163. [Google Scholar] [CrossRef] [Green Version]
- Vannini, M.; Emery, S.; Lieutier-Colas, F.; Legueult, K.; Mondain, V.; Retur, N.; Gastaud, L.; Pomares, C.; Hasseine, L. Epidemiology of candidemia in NICE area, France: A five-year study of antifungal susceptibility and mortality. J. Mycol. Med. 2021, 32, 101210. [Google Scholar] [CrossRef]
- Memariani, H.; Memariani, M. Anti-fungal properties and mechanisms of melittin. Appl. Microbiol. Biotechnol. 2020, 104, 6513–6526. [Google Scholar] [CrossRef]
- Healey, K.R.; Zhao, Y.; Perez, W.B.; Lockhart, S.R.; Sobel, J.D.; Farmakiotis, D.; Kontoyiannis, D.P.; Sanglard, D.; Taj-Aldeen, S.J.; Alexander, B.D.; et al. Prevalent mutator genotype identified in fungal pathogen Candida glabrata promotes multi-drug resistance. Nat. Commun. 2016, 7, 11128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanglard, D. Emerging threats in antifungal-resistant fungal pathogens. Front. Med. 2016, 3, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfaller, M.A.; Castanheira, M.; Lockhart, S.R.; Ahlquist, A.M.; Messer, S.A.; Jones, R.N. Frequency of decreased susceptibility and resistance to echinocandins among fluconazole-resistant bloodstream isolates of Candida glabrata. J. Clin. Microbiol. 2012, 50, 1199–1203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmalreck, A.F.; Lackner, M.; Becker, K.; Fegeler, W.; Czaika, V.; Ulmer, H.; Lass-Flörl, C. Phylogenetic relationships matter: Antifungal susceptibility among clinically relevant yeasts. Antimicrob. Agents Chemother. 2014, 58, 1575–1585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López-García, B.; Gandía, M.; Muñoz, A.; Carmona, L.; Marcos, J.F. A genomic approach highlights common and diverse effects and determinants of susceptibility on the yeast Saccharomyces cerevisiae exposed to distinct antimicrobial peptides. BMC Microbiol. 2010, 10, 289. [Google Scholar] [CrossRef] [Green Version]
- Andrä, J.; Berninghausen, O.; Leippe, M. Cecropins, antibacterial peptides from insects and mammals, are potently fungicidal against Candida albicans. Med. Microbiol. Immunol. 2001, 189, 169–173. [Google Scholar] [CrossRef]
- Kim, D.H.; Lee, D.G.; Kim, K.L.; Lee, Y. Internalization of tenecin 3 by a fungal cellular process is essential for its fungicidal effect on Candida albicans. Eur. J. Biochem. 2001, 268, 4449–4458. [Google Scholar] [CrossRef] [Green Version]
- De Groot, P.W.; Kraneveld, E.A.; Yin, Q.Y.; Dekker, H.L.; Gross, U.; Crielaard, W.; de Koster, C.G.; Bader, O.; Klis, F.M.; Weig, M. The cell wall of the human pathogen Candida glabrata: Differential incorporation of novel adhesin-like wall proteins. Eukaryot. Cell. 2008, 7, 1951–1964. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptide | Amino Acid Sequence | Net Charge | Hydrophobicity (H) | Hydrophobic Moment (μH) | pI | α-Helicity (%) in | ζ-Potential (mV) | |
|---|---|---|---|---|---|---|---|---|
| PBS | TFE | |||||||
| MB | LKLKSIVSWAKKVL-NH2 | +5 | 0.561 | 0.404 | 14.0 | 6.0 | 38.2 | 6.8 ± 2.5 |
| MG | GIGKFLHSAGKFGKAFVGEIMKS | +3 | 0.42 | 0.515 | 10.0 | 1.6 | 35.2 | 5.6 ± 1.7 |
| ML | GIGAVLKVLTTGLPALISWIKRKRQQ-NH2 | +6 | 0.511 | 0.394 | 14.0 | 5.6 | 47.5 | 10.8 ± 0.4 |
| CA | KWKLFKKIEKVGQNIRDGIIKAGPAVAVVGQATQIAK-NH2 | +7 | 0.312 | 0.202 | 11.4 | 2.5 | 18.8 | 7.8 ± 2.1 |
| Strains | Source | MIC of Peptides in μM | ||||
|---|---|---|---|---|---|---|
| MB | MG | ML | CA | Amphotericin B | ||
| C. albicans | ||||||
| ATCC 10231 | - | 2.52 | 26.48 | 2.82 | 2.00 | 0.14 |
| ATCC 2091 | - | 2.52 | 26.48 | 1.41 | 2.00 | 0.14 |
| ATCC 24433 | - | 1.26 | 13.24 | 1.41 | 2.00 | 0.14 |
| 1976R | Colon | 0.62 | 6.63 | 1.41 | 0.50 | 0.54 |
| 2672R | Urine | 0.62 | 13.24 | 1.41 | 2.00 | 0.14 |
| C. tropicalis | ||||||
| CT 1001 | Blood | 0.62 | 0.82 | 2.82 | 2.00 | 0.54 |
| CT 1002 | Blood | 0.31 | 0.82 | 5.64 | 1.00 | 0.54 |
| CT 1003 | Blood | 1.24 | 3.31 | 2.82 | 4.00 | 0.54 |
| CT 1004 | Blood | 0.62 | 0.82 | 5.62 | 1.00 | 0.54 |
| CT 1005 | Blood | 0.62 | 0.82 | 2.82 | 1.00 | 0.54 |
| CT 1006 | Blood | 1.24 | 0.82 | 5.64 | 1.00 | 0.54 |
| CT 1007 | Blood | 1.24 | 3.31 | 22.56 | 4.00 | 0.54 |
| CT 1008 | Blood | 0.62 | 3.31 | 11.24 | 4.00 | 0.54 |
| CT 1009 | Blood | 0.62 | 0.82 | 2.82 | 1.00 | 0.54 |
| CT 1010 | Blood | 0.31 | 0.82 | 2.82 | 1.00 | 0.54 |
| C. parapsilosis | ||||||
| CP 1001 | Peritoneal | 2.48 | 6.62 | 5.62 | NA | 1.08 |
| CP 1002 | Blood | 4.96 | 6.62 | 11.24 | NA | 1.08 |
| CP 1003 | Blood | 4.96 | 6.62 | 11.24 | NA | 1.08 |
| CP 1004 | Others | 0.62 | 3.31 | 22.48 | NA | 0.54 |
| CP 1005 | Blood | 4.96 | 13.24 | 5.62 | NA | 1.08 |
| CP 1006 | Blood | 4.96 | 13.24 | 5.62 | NA | 1.08 |
| CP 1007 | Blood | 4.96 | 13.24 | 5.62 | NA | 1.08 |
| CP 1008 | Blood | 2.48 | 13.24 | >22.48 | NA | 1.08 |
| CP 1009 | Blood | 1.24 | 13.24 | 11.24 | NA | 1.08 |
| CP 1010 | Blood | 4.96 | 26.48 | 5.62 | NA | 1.08 |
| C. glabrata | ||||||
| CG 1001 | Blood | 9.92 | 26.48 | >22.48 | NA | 0.27 |
| CG 1002 | Pleural fluid | 9.92 | 26.48 | >22.48 | NA | 0.27 |
| CG 1003 | Blood | 4.96 | 26.48 | >22.48 | NA | 0.27 |
| CG 1004 | Blood | 79.40 | >26.48 | >22.48 | NA | 0.14 |
| CG 1005 | Blood | 39.70 | >26.48 | >22.48 | NA | 0.27 |
| CG1006 | Blood | >79.40 | >26.48 | >22.48 | NA | 0.27 |
| CG 1007 | Blood | 4.96 | 13.24 | 11.28 | NA | 0.27 |
| CG 1008 | Blood | 2.98 | 6.62 | 11.28 | NA | 0.27 |
| CG 1009 | Peritoneal fluid | 4.96 | 26.48 | >22.48 | NA | 0.27 |
| CG 1010 | Blood | 9.92 | 26.48 | 22.48 | NA | 0.27 |
| Bacterial Viability in CFU/mL | ||
|---|---|---|
| No Peptide | MB | |
| SD broth | 184,250 ± 50,500 | 500 |
| +25 mM NaCl | 161,000 ± 21,000 | 3350 ± 250 |
| +50 mM NaCl | 8400 ± 1100 | |
| +100 mM NaCl | 21,900 ± 1500 | |
| +150 mM NaCl | 65,500 ± 3000 | |
| +0.5 mM CaCl2 | 157,000 ± 27,000 | 900 |
| +1.0 mM CaCl2 | 3100 ± 200 | |
| +1.5 mM CaCl2 | 6800 ± 500 | |
| +2.0 mM CaCl2 | 5900 ± 200 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lim, E.J.; Leng, E.G.T.; Tram, N.D.T.; Periayah, M.H.; Ee, P.L.R.; Barkham, T.M.S.; Poh, Z.S.; Verma, N.K.; Lakshminarayanan, R. Rationalisation of Antifungal Properties of α-Helical Pore-Forming Peptide, Mastoparan B. Molecules 2022, 27, 1438. https://doi.org/10.3390/molecules27041438
Lim EJ, Leng EGT, Tram NDT, Periayah MH, Ee PLR, Barkham TMS, Poh ZS, Verma NK, Lakshminarayanan R. Rationalisation of Antifungal Properties of α-Helical Pore-Forming Peptide, Mastoparan B. Molecules. 2022; 27(4):1438. https://doi.org/10.3390/molecules27041438
Chicago/Turabian StyleLim, Edward Jianyang, Eunice Goh Tze Leng, Nhan Dai Thien Tram, Mercy Halleluyah Periayah, Pui Lai Rachel Ee, Timothy Mark Sebastian Barkham, Zhi Sheng Poh, Navin Kumar Verma, and Rajamani Lakshminarayanan. 2022. "Rationalisation of Antifungal Properties of α-Helical Pore-Forming Peptide, Mastoparan B" Molecules 27, no. 4: 1438. https://doi.org/10.3390/molecules27041438
APA StyleLim, E. J., Leng, E. G. T., Tram, N. D. T., Periayah, M. H., Ee, P. L. R., Barkham, T. M. S., Poh, Z. S., Verma, N. K., & Lakshminarayanan, R. (2022). Rationalisation of Antifungal Properties of α-Helical Pore-Forming Peptide, Mastoparan B. Molecules, 27(4), 1438. https://doi.org/10.3390/molecules27041438