
Anticancer Activities of Marine-Derived Phenolic Compounds and Their Derivatives

 , ,
, ,  and
and
Abstract
:
1. Introduction—Marine Natural Products as Anticancer Agents
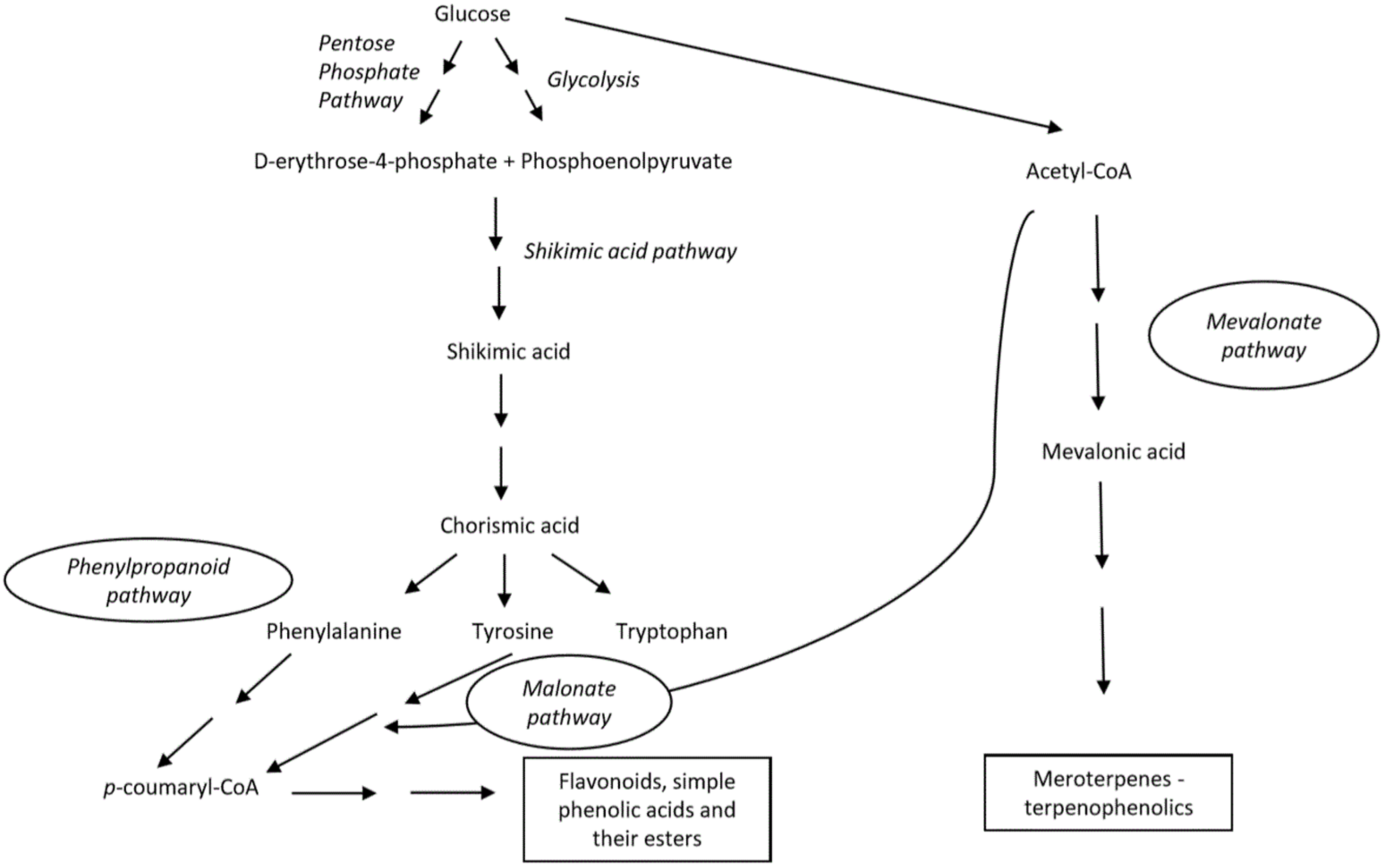
2. Phenolic Compounds of Marine Origin—General Characteristics and Biosynthesis
3. Anticancer Properties of Marine-Derived Phenolic Compounds
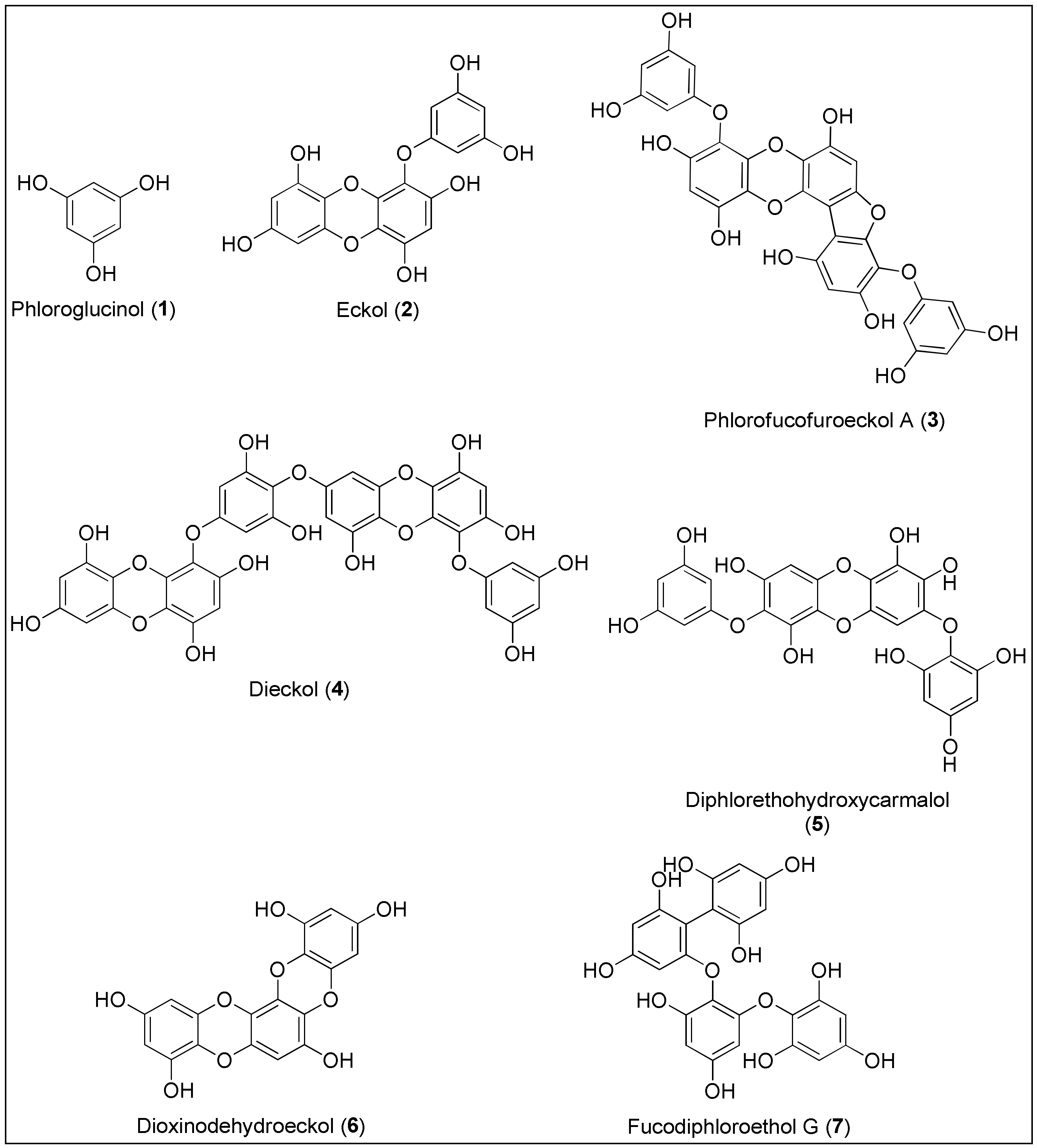
3.1. Phlorotannins
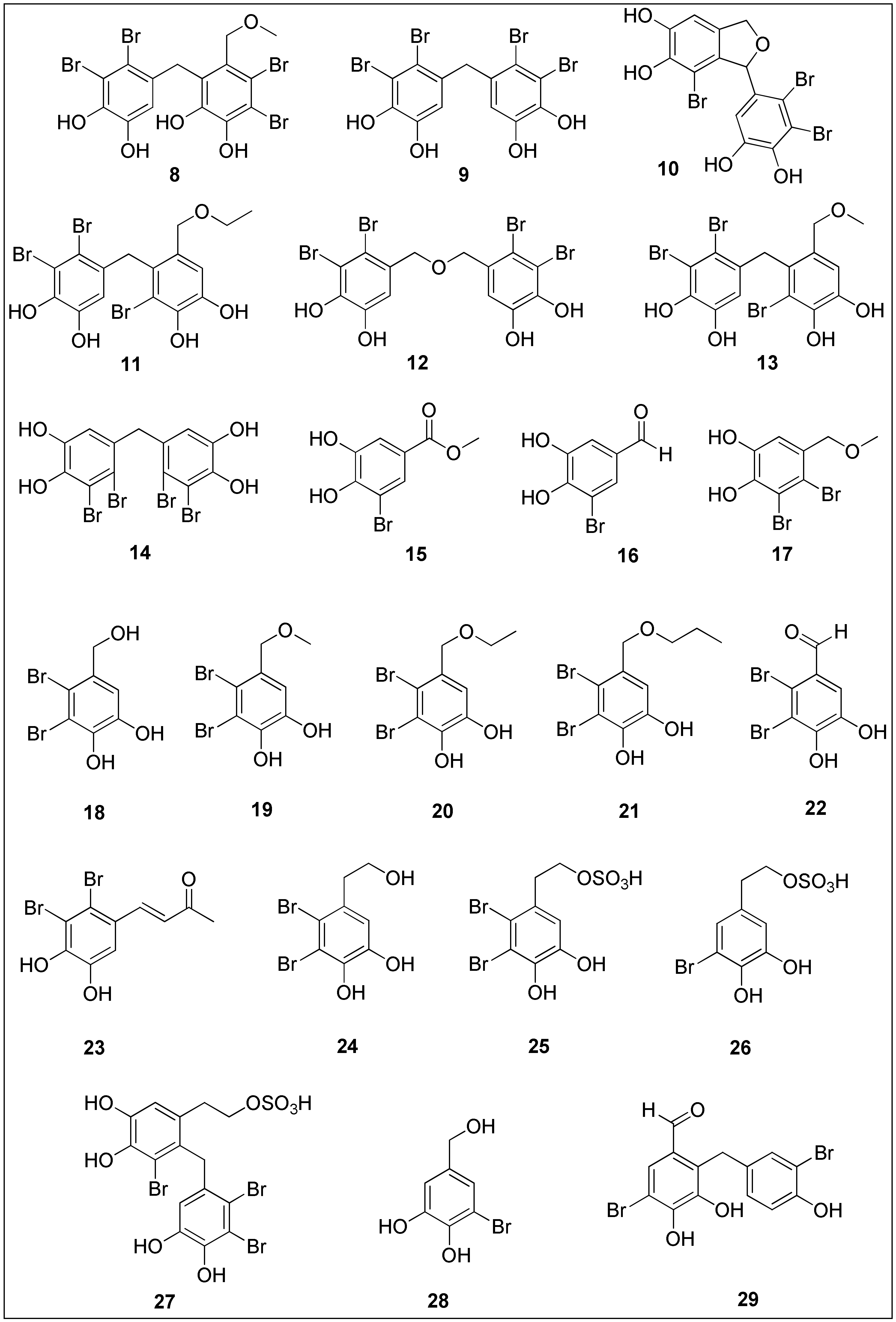
3.2. Bromophenols
3.3. Flavonoids
3.4. Coumarins
3.5. Terpenophenolics
3.6. Quinones and Hydroquinones
3.7. Miscellaneous Compounds
4. Conclusions
List of Abbreviations
| 26-L5 | murine colon adenocarcinoma cell line |
| 3T3-L1 | mouse embryonic fibroblast cell line |
| 4CL | 4-coumaroyl CoA ligase |
| A2780 | human ovarian cancer cell line |
| A375 | human melanoma cell line |
| A431 | epidermoid carcinoma cell line |
| A549 | adenocarcinomic human alveolar basal epithelial cell line |
| A5490 | airway epithelial cells and mitochondrial DNA depleted cancer cell line |
| AKT | serine/threonine-specific protein kinase (PKB) |
| AP-1 | transcription factor |
| APN | aminopeptidase N |
| ATF3 | Activating Transcription Factor 3 |
| AUXB1 | wild type of chinese hamster ovary cells, CHO |
| B16-BL6 | murine melanoma cell line |
| B16F0 | murine melanoma cell line, parent cells |
| B16F10 | murine melanoma cell line from a C57BL/6J mouse |
| Bad | Bcl2-associated agonist of cell death |
| Bax | Bcl-2-associated X protein |
| Bcl-2 | B-cell lymphoma 2 |
| Bcl-xL | B-cell lymphoma-extra large |
| BDDE | bis (2,3-dibromo-4,5-dihydroxybenzyl) ether |
| BDDPM | bis (2,3-dibromo-4,5-dihydroxy-phenyl)-methane |
| Bel7402 | human hepatocellular carcinoma cell line |
| BGC-823 | human gastric carcinoma cell line |
| BT-474 | human breast carcinoma are characterized by the overexpression of human epidermal growth factor receptors 2 (HER-2) and estrogen receptors (ER) |
| C4H | cinnamate 4-hydroxlyase |
| C6 | rat glioma cell line |
| Caco2 | human colorectal adenocarcinoma cells |
| Calu-3 | human lung cancer cell line |
| CCHRC5 | AUXB1 cells resistant to doxorubicin |
| CCRF-CEM | leukemia cell line |
| CD44 | cell-surface glycoprotein |
| CDK2 | Cyclin-dependent Kinase 2 |
| CEM | line of lymphoblastic cells originally derived from a child with acute lymphoblastic leukemia |
| CEM/C2 | camptothecin (CPT) resistant derivative of the human T cell leukemia cell line CCRF-CEM |
| CFU-GM | granulocyte–macrophage progenitor (GMP) |
| cIAP2 | baculoviral IAP repeat-containing protein 3 |
| c-Jun | proto-oncogene, transcription factor AP-1 |
| Colon-38 | human colon cancer cell line |
| COX-2 | cyclooxigenase-2 |
| DLD-1 | human colon cancer cell line |
| DOT1L | disruptor of telomeric silencing-1 like |
| DU-145 | human prostate cancer cell line |
| EA.hy926 | human umbilical vein endothelial cells |
| ECM | Extracellular Matrix |
| ECV-304 | human umbilical vein endothelial cells |
| EGFR-TK | Epidermal Growth Factor Receptor Tyrosine Kinase |
| eNOS | Nitric Oxide Synthase, endothelial |
| ERK | Extracellular signal-Regulated Kinases |
| GXF251L | human gastric carcinoma cells |
| H-125 | human lung cancer cell line |
| H22 | murine hepatic carcinoma cell line |
| H2887 | non-small-cell lung cancer cell line |
| H460 | large cell lung cancer cell line |
| H460/MX20 | large cell lung cancer cell line, derived from H460, mitoxantrone induced ABCG2-overexpressing cells |
| HaCaT | human keratinocyte cells |
| HCC | hepatocellular carcinoma |
| HCT116 | human colon cancer cell line |
| HCT-15 | human colon adenocarcinoma cell line |
| HCT-8 | human ileocaecal adenocarcinoma cell line |
| HDAC | histone deacetylase |
| HeLa | human cervical cancer cell line |
| HELF | embryo lung fibroblasts |
| Hep3B | human hepatoma cell line |
| HepG2 | human hepatoma cell line |
| HIF-1α | hypoxia-inducible factor 1-alpha |
| HL-60 | leukemia cells |
| HT-1080 | fibrosarcoma cell line |
| HT-29 | human colon cancer cell line |
| Huh-7 | human hepatoma cell lines |
| HUVEC | human endothelial cells |
| IFN-γ | Interferon gamma |
| IGF-1R | Insulin-like Growth Factor 1 Receptor |
| IKK | IκB kinase |
| K562 | human myelogenous leukemia cells |
| KB | epidermoid carcinoma cell lines |
| KBv200 | epidermoid carcinoma cell lines, multi-drug resistant (MDR) cell line |
| L1210 | leukemia cells |
| L5178Y | human leukemia monocytic cell line |
| LNCaP | androgen-sensitive human prostate adenocarcinoma cell line |
| LoVo | human colon cancer cell line, supraclavicular lymph node metastasis |
| LXF529L | lung cancer cell line |
| MAPK | mitogen-activated protein kinase |
| MARCHF1 | E3 ubiquitin-protein ligase MARCHF1 |
| MAXF401NL | human mammary cancer cell line |
| MCA | Mycothiol-S-conjugate Amidase |
| MCF-7 | human breast cancer cell line |
| MCF-7/ADR | Adriamycin-resistant human breast cancer cell line |
| MDA-MB-231 | epithelial, human breast cancer cell line |
| MEL28 | human melanoma cell line |
| MEXF462NL | human melanoma cell line |
| MITF | microphthalmia-associated transcription factor |
| MMP3 | Matrix Metalloproteinase-3 |
| MRC-5 | normal fibroblast cell line |
| mTOR | Mammalian Target of Rapamycin |
| MTT | 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide, reagent |
| NCI-H460 | human lung carcinoma epithelial cells |
| NDEA | N-nitrosodiethylamine |
| Neuro-2a | neuroblastoma cell line |
| NF-κB | Nuclear Factor kappa-light-chain-enhancer of activated B cells |
| NIH-3T3 | murine fibroblast cell line |
| NIK | NF-κB inducing kinase |
| Notch2 | Neurogenic Locus Notch Homolog Protein 2 |
| NSCLC-N6 | non-small cell lung cancer cell line |
| Oct4 | Octamer-binding transcription factor 4 |
| OVCAR-5 | human epithelial ovary carcinoma cell line |
| P-338 | sensitive murine leukemia cells |
| p38 | mitogen-activated protein kinases |
| P-388 | bipotential murine pre-B cell lymphoma |
| P388CPT5 | murine leukemia cells resistant to the reference topoisomerase I poison camptothecin (CPT) |
| p70S6K | ribosomal protein S6 kinase beta-1 |
| PAL | Phenylalanine Ammonia-Lyase |
| PANC-1 | human pancreatic cancer cell line |
| PARP | Poly (ADP-ribose) Polymerase |
| PAXF1657L | human pancreatic tumor cell line |
| PC3 | bone metastasis of a grade IV prostatic adenocarcinoma cell line |
| PCNA | proliferating cell nuclear antigen |
| PI3K | Phosphoinositide 3-Kinase |
| PKS | Polyketide Synthase |
| Rac1 | Ras-related C3 botulinum toxin substrate 1 |
| RAF | Rapidly Accelerated Fibrosarcoma, protein kinase |
| RAS | Rat Sarcoma Virus, GTP-ase |
| RD | human rhabdomyosarcoma cell line |
| Reg3A | regenerating family member 3 alpha |
| Reg3g | regenerating islet-derived protein 3 gamma |
| RhoGDI2 | Rho GDP dissociation inhibitor 2 |
| RKO | colon carcinoma cell line |
| ROS | Reactive Oxygen Species |
| RXF486L | human renal cancer cell line |
| S1 | human colon cancer cell line |
| S1-M1-80 | human colon cancer cell line, derived from S1 cells, mitoxantrone-selected ABCG2-overexpressing cells |
| SHG-44 | human glioma cell line |
| SH-SY5Y | cell line human neuroblast from neural tissue |
| SJCRH30 | rhabdomycosarcoma cancer cell line |
| SKBR3 | human breast cancer cell line, hypertriploid |
| SK-Hep1 | human hepatic adenocarcinoma cell line |
| SKOV3 | ovarian carcinoma cells |
| SLUG | transcription factor (SNAI2) |
| SNU638 | human stomach carcinoma cell line |
| Sox2 | (sex determining region Y)-box 2, trancription factor |
| Src | proto-oncogene tyrosine-protein kinase Src |
| STAT3 | Signal Transducer and Activator of Transcription 3 |
| SW1990 | spleen metastasis of a grade II pancreatic adenocarcinoma |
| SW480 | primary colon adenocarcinoma cell line |
| TAL | Tyrosine Ammonia Lyase |
| THP-1 | human leukemia monocytic cell line |
| TIMP-1 | TIMP Metallopeptidase Inhibitor 1 |
| TLR-4 | Toll-like receptor 4 |
| TNF-α | Tumor Necrosis Factor alpha |
| TOP-2 | DNA topoisomerase 2-alpha |
| TRP-1 | tyrosinase and tyrosinase-linked proteins |
| TRP-2 | tyrosinase and tyrosinase-linked proteins |
| U251 | glioblastoma-derived human cell line |
| U87 | human primary glioblastoma cell line, synonym: U87-MG |
| V79 | chinese hamster fibroblasts |
| VDAC1 | Voltage-dependent Anion-selective Channel 1 |
| VEGF | Vascular Endothelial Growth Factor |
| VEGFR | Vascular Endothelial Growth Factor Receptor |
| WAVE2 | Wiskott–Aldrich syndrome protein family member 2 |
| WST-1 | cell proliferation reagent |
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Torre, L.A.; Bray, F.; Siegel, R.L.; Ferlay, J.; Lortet-Tieulent, J.; Jemal, A. Global cancer statistics, 2012. CA Cancer J. Clin. 2015, 65, 87–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lefranc, F.; Koutsaviti, A.; Ioannou, E.; Kornienko, A.; Roussis, V.; Kiss, R.; Newman, D. Algae metabolites: From in vitro growth inhibitory effects to promising anticancer activity. Nat. Prod. Rep. 2019, 36, 810–841. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [Green Version]
- Al-Khater, K.M.; Almofty, S.; Ravinayagam, V.; Alrushaid, N.; Rehman, S. Role of a metastatic suppressor gene KAI1/CD82 in the diagnosis and prognosis of breast cancer. Saudi J. Biol. Sci. 2021, 28, 3391–3398. [Google Scholar] [CrossRef]
- Fife, C.M.; McCarroll, J.A.; Kavallaris, M. Movers and shakers: Cell cytoskeleton in cancer metastasis. Br. J. Pharmacol. 2014, 171, 5507–5523. [Google Scholar] [CrossRef] [Green Version]
- Rocha, D.H.A.; Seca, A.M.L.; Pinto, D.C.G.A. Seaweed Secondary Metabolites: In Vitro and In Vivo Anticancer Activity. Mar. Drugs 2018, 16, 410. [Google Scholar] [CrossRef] [Green Version]
- Arnedos, M.; Soria, J.C.; Andre, F.; Tursz, T. Personalized treatments of cancer patients: A reality in daily practice, a costly dream or a shared vision of the future from the oncology community? Cancer Treat. Rev. 2014, 40, 1192–1198. [Google Scholar] [CrossRef]
- Apetoh, L.; Ladoire, S.; Coukos, G.; Ghiringhelli, F. Combining immunotherapy and anticancer agents: The right path to achieve cancer cure? Ann. Oncol. 2015, 26, 1813–1823. [Google Scholar] [CrossRef]
- Ramsay, R.R.; Popovic-Nikolic, M.R.; Nikolic, K.; Uliassi, E.; Bolognesi, M.L. A perspective on multi-target drug discovery and design for complex diseases. Clin. Transl. Med. 2018, 7, 31–61. [Google Scholar] [CrossRef] [Green Version]
- Ercolano, G.; De Cicco, P.; Ianaro, A. New Drugs from the Sea: Pro-Apoptotic Activity of Sponges and Algae Derived Compounds. Mar. Drugs 2019, 17, 31. [Google Scholar] [CrossRef] [Green Version]
- Palanisamy, S.K.; Rajendran, N.M.; Marino, A. Natural Products Diversity of Marine Ascidians (Tunicates; Ascidiacea) and Successful Drugs in Clinical Development. Nat. Prod. Bioprospect. 2017, 7, 1795995. [Google Scholar] [CrossRef] [Green Version]
- Hamed, I.; Özogul, F.; Özogul, Y.; Regenstein, J.M. Marine Bioactive Compounds and Their Health Benefits: A Review. Compr. Rev. Food Sci. Food Saf. 2015, 14, 446–465. [Google Scholar] [CrossRef]
- Suleria, H.A.R.; Gobe, G.; Masci, P.; Osborne, S.A. Marine bioactive compounds and health promoting perspectives; innovation pathways for drug discovery. Trends Food Sci. Technol. 2016, 50, 44–55. [Google Scholar] [CrossRef]
- Matulja, D.; Markovic, M.K.; Ambrožić, G.; Laclef, S.; Pavelić, S.K.; Marković, D. Secondary metabolites from gorgonian corals of the genus Eunicella: Structural characterizations, biological activities, and synthetic approaches. Molecules 2020, 25, 129. [Google Scholar] [CrossRef] [Green Version]
- Matulja, D.; Wittine, K.; Malatesti, N.; Laclef, S.; Turks, M.; Markovic, M.K.; Ambrožić, G.; Marković, D. Marine Natural Products with High Anticancer Activities. Curr. Med. Chem. 2020, 27, 1243–1307. [Google Scholar] [CrossRef]
- Calado, R.; Leal, M.C.; Gaspar, H.; Santos, S.; Marques, A.; Nunes, M.L.; Vieira, H. How to Succeed in Marketing Marine Natural Products for Nutraceutical, Pharmaceutical and Cosmeceutical Markets. In Grand Challenges in Marine Biotechnology; Rampelotto, P.H., Trincone, A., Eds.; Springer International Publishing: Manhattan, NY, USA, 2018; pp. 317–403. ISBN 978-3-319-69075-9. [Google Scholar]
- Jiménez, C. Marine Natural Products in Medicinal Chemistry. ACS Med. Chem. Lett. 2018, 9, 959–961. [Google Scholar] [CrossRef] [Green Version]
- Alves, C.; Silva, J.; Pinteus, S.; Gaspar, H.; Alpoim, M.C.; Botana, L.M.; Pedrosa, R. From marine origin to therapeutics: The antitumor potential of marine algae-derived compounds. Front. Pharmacol. 2018, 9, 777–800. [Google Scholar] [CrossRef] [Green Version]
- Deeks, E.D. Polatuzumab Vedotin: First Global Approval. Drugs 2019, 79, 1467–1475. [Google Scholar] [CrossRef] [Green Version]
- Chang, E.; Weinstock, C.; Zhang, L.; Charlab, R.; Dorff, S.E.; Gong, Y.; Hsu, V.; Li, F.; Ricks, T.K.; Song, P.; et al. FDA Approval Summary: Enfortumab Vedotin for Locally Advanced or Metastatic Urothelial Carcinoma. Clin. Cancer Res. 2020, 27, 922–927. [Google Scholar] [CrossRef]
- Barreca, M.; Spanò, V.; Montalbano, A.; Cueto, M.; Díaz Marrero, A.R.; Deniz, I.; Erdoğan, A.; Lukić Bilela, L.; Moulin, C.; Taffin-de-Givenchy, E.; et al. Marine Anticancer Agents: An Overview with a Particular Focus on Their Chemical Classes. Mar. Drugs 2020, 18, 619. [Google Scholar] [CrossRef]
- Kepp, O.; Zitvogel, L.; Kroemer, G. Lurbinectedin: An FDA-approved inducer of immunogenic cell death for the treatment of small-cell lung cancer. Oncoimmunology 2020, 9, 1–3. [Google Scholar] [CrossRef]
- Markham, A. Belantamab Mafodotin: First Approval. Drugs 2020, 80, 1607–1613. [Google Scholar] [CrossRef]
- Jimenez, P.C.; Wilke, D.V.; Branco, P.C.; Bauermeister, A.; Rezende-Teixeira, P.; Gaudêncio, S.P.; Costa-Lotufo, L.V. Enriching cancer pharmacology with drugs of marine origin. Br. J. Pharmacol. 2020, 177, 3–27. [Google Scholar] [CrossRef] [Green Version]
- Cotas, J.; Leandro, A.; Monteiro, P.; Pacheco, D.; Figueirinha, A.; Goncąlves, A.M.M.; Da Silva, G.J.; Pereira, L. Seaweed phenolics: From extraction to applications. Mar. Drugs 2020, 18, 384. [Google Scholar] [CrossRef]
- Mekinić, I.G.; Skroza, D.; Šimat, V.; Hamed, I.; Čagalj, M.; Perković, Z.P. Phenolic content of brown algae (Pheophyceae) species: Extraction, identification, and quantification. Biomolecules 2019, 9, 244. [Google Scholar] [CrossRef] [Green Version]
- Utkina, N.K.; Makarchenko, A.E.; Shchelokova, O.V.; Virovaya, M.V. Antioxidant activity of phenolic metabolites from marine sponges. Chem. Nat. Compd. 2004, 40, 373–377. [Google Scholar] [CrossRef]
- Cichewicz, R.H.; Clifford, L.J.; Lassen, P.R.; Cao, X.; Freedman, T.B.; Nafie, L.A.; Deschamps, J.D.; Kenyon, V.A.; Flanary, J.R.; Holman, T.R.; et al. Stereochemical determination and bioactivity assessment of (S)-(+)-curcuphenol dimers isolated from the marine sponge Didiscus aceratus and synthesized through laccase biocatalysis. Bioorganic Med. Chem. 2005, 13, 5600–5612. [Google Scholar] [CrossRef]
- Putra, M.Y.; Murniasih, T.; Swasono, R.T.; Wibowo, J.T.; Saputri, A.N.C.; Widhiana, M.R.; Arlyza, I.S. Secondary metabolites and their biological activities in Indonesian soft coral of the genus Lobophytum. Asian Pac. J. Trop. Biomed. 2016, 6, 909–913. [Google Scholar] [CrossRef] [Green Version]
- Deghrigue, M.; Dellai, A.; Akremi, N.; Le Morvan, V.; Robert, J.; Bouraoui, A. Evaluation of antiproliferative and antioxidant activities of the organic extract and its polar fractions from the Mediterranean gorgonian Eunicella singularis. Environ. Toxicol. Pharmacol. 2013, 36, 339–346. [Google Scholar] [CrossRef]
- Deghrigue, M.; Dellai, A.; Bouraoui, A.; Akremi, N.; Le Morvan, V.; Robert, J.; Bouraoui, A. In Vitro Antiproliferative and Antioxidant Activities of the Organic Extract and Its Semi-Purified Fractions from the Mediterranean Gorgonian Eunicella singularis. Int. J. Pharm. Pharm. Sci. 2013, 5, 432–439. [Google Scholar]
- Matulja, D.; Grbčić, P.; Bojanić, K.; Topić-Popović, N.; Čož-Rakovac, R.; Laclef, S.; Šmuc, T.; Jović, O.; Marković, D.; Pavelić, S.K. Chemical evaluation, antioxidant, antiproliferative, anti-inflammatory and antibacterial activities of organic extract and semi-purified fractions of the Adriatic sea fan, Eunicella cavolini. Molecules 2021, 26, 5751. [Google Scholar] [CrossRef] [PubMed]
- Hussein, H.A.; Abdullah, M.A. Anticancer Compounds Derived from Marine Diatoms. Mar. Drugs 2020, 18, 356. [Google Scholar] [CrossRef]
- Zheng, Y.; Chen, X.; Chen, L.; Shen, L.; Fu, X.; Chen, Q.; Chen, M.; Wang, C. Isolation and Neuroprotective Activity of Phenolic Derivatives from the Marine-Derived Fungus Penicillium janthinellum. J. Ocean Univ. China 2020, 19, 700–706. [Google Scholar] [CrossRef]
- Kodzius, R.; Gojobori, T. Marine metagenomics as a source for bioprospecting. Mar. Genomics 2015, 24, 21–30. [Google Scholar] [CrossRef] [Green Version]
- Metsämuuronen, S.; Sirén, H. Bioactive phenolic compounds, metabolism and properties: A review on valuable chemical compounds in Scots pine and Norway spruce. Phytochem. Rev. 2019, 18, 623–664. [Google Scholar] [CrossRef] [Green Version]
- Bertelli, A.; Biagi, M.; Corsini, M.; Baini, G.; Cappellucci, G.; Miraldi, E. Polyphenols: From Theory to Practice. Foods 2021, 10, 2595. [Google Scholar] [CrossRef] [PubMed]
- Mageroy, M.H.; Jancsik, S.; Saint Yuen, M.M.; Fischer, M.; Withers, S.G.; Paetz, C.; Schneider, B.; Mackay, J.; Bohlmann, J. A conifer UDP-sugar dependent glycosyltransferase contributes to acetophenone metabolism and defense against insects. Plant Physiol. 2017, 175, 641–651. [Google Scholar] [CrossRef]
- Mateos, R.; Pérez-Correa, J.R.; Domínguez, H. Bioactive properties of marine phenolics. Mar. Drugs 2020, 18, 501. [Google Scholar] [CrossRef]
- Vacca, R.A.; Valenti, D.; Caccamese, S.; Daglia, M.; Braidy, N.; Nabavi, S.M. Plant polyphenols as natural drugs for the management of Down syndrome and related disorders. Neurosci. Biobehav. Rev. 2016, 71, 865–877. [Google Scholar] [CrossRef]
- Taskin, D.; Ozdemir, M.; Yalcin, B. LC-ESI-tandem MS and in silico ADMET analysis of polyphenols from Rhus coriaria L. and Micromeria fruticosa (L.) Druce ssp. brachycalyx P. H. Davis. Futur. J. Pharm. Sci. 2021, 7, 168. [Google Scholar] [CrossRef]
- Getachew, A.T.; Jacobsen, C.; Holdt, S.L. Emerging technologies for the extraction of marine phenolics: Opportunities and challenges. Mar. Drugs 2020, 18, 389. [Google Scholar] [CrossRef]
- Bhatia, P.; Chugh, A. Role of marine bioprospecting contracts in developing access and benefit sharing mechanism for marine traditional knowledge holders in the pharmaceutical industry. Glob. Ecol. Conserv. 2015, 3, 176–187. [Google Scholar] [CrossRef] [Green Version]
- Berni, R.; Cai, G.; Hausman, J.F.; Guerriero, G. Plant fibers and phenolics: A review on their synthesis, analysis and combined use for biomaterials with new properties. Fibers 2019, 7, 80. [Google Scholar] [CrossRef] [Green Version]
- Stewart, A.J.; Stewart, R.F. Phenols. In Encyclopedia of Ecology; Jørgensen, S.E., Fath, B.D., Eds.; Academic Press: Oxford, UK, 2008; pp. 2682–2689. ISBN 9780080914565. [Google Scholar]
- Anantharaju, P.G.; Gowda, P.C.; Vimalambike, M.G.; Madhunapantula, S.V. An overview on the role of dietary phenolics for the treatment of cancers. Nutr. J. 2016, 15, 99. [Google Scholar] [CrossRef] [Green Version]
- Saltveit, M.E. Synthesis and metabolism of phenolic compounds. In Fruit and Vegetable Phytochemicals: Chemistry and Human Health; Yahia, E.M., Ed.; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2017; pp. 115–124. ISBN 9781119158042. [Google Scholar]
- Francenia Santos-Sánchez, N.; Salas-Coronado, R.; Hernández-Carlos, B.; Villanueva-Cañongo, C. Shikimic Acid Pathway in Biosynthesis of Phenolic Compounds. In Plant Physiological Aspects of Phenolic Compounds; Soto-Hernández, S., García-Mateos, R., PalmaTenango, M., Eds.; IntechOpen: London, UK, 2019; Chapter 3; pp. 35–50. [Google Scholar]
- Tohge, T.; De Souza, L.P.; Fernie, A.R. Current understanding of the pathways of flavonoid biosynthesis in model and crop plants. J. Exp. Bot. 2017, 68, 4013–4028. [Google Scholar] [CrossRef]
- Babenko, L.M.; Smirnov, O.E.; Romanenko, K.O.; Trunova, O.K.; Kosakivska, I.V. Phenolic compounds in plants: Biogenesis and functions. Ukr. Biochem. J. 2019, 91, 5–18. [Google Scholar] [CrossRef]
- Kumari, S.; Priya, P.; Misra, G.; Yadav, G. Structural and biochemical perspectives in plant isoprenoid biosynthesis. Phytochem. Rev. 2013, 12, 255–291. [Google Scholar] [CrossRef]
- Mandrekar, V.K.; Gawas, U.B.; Majik, M.S. Brominated Molecules From Marine Algae and Their Pharmacological Importance. In Studies in Natural Products Chemistry; Atta-ur-Rahman, Ed.; Elsevier B.V.: Amsterdam, The Netherlands, 2018; Volume 61, pp. 461–490. ISBN 978-0-444-64183-0. [Google Scholar]
- Bertoni, G. A key step in phlorotannin biosynthesis revealed. Plant Cell 2013, 25, 2770. [Google Scholar] [CrossRef] [Green Version]
- Meslet-Cladière, L.; Delage, L.; Leroux, C.J.J.; Goulitquer, S.; Leblanc, C.; Creis, E.; Gall, E.A.; Stiger-Pouvreau, V.; Czjzek, M.; Potin, P. Structure/function analysis of a type III polyketide synthase in the brown alga Ectocarpus siliculosus reveals a biochemical pathway in phlorotannin monomer biosynthesis. Plant Cell 2013, 25, 3089–3103. [Google Scholar] [CrossRef] [Green Version]
- Jimenez-Lopez, C.; Pereira, A.G.; Lourenço-Lopes, C.; Garcia-Oliveira, P.; Cassani, L.; Fraga-Corral, M.; Prieto, M.A.; Simal-Gandara, J. Main bioactive phenolic compounds in marine algae and their mechanisms of action supporting potential health benefits. Food Chem. 2021, 341, 128262. [Google Scholar] [CrossRef] [PubMed]
- Di Meo, F.; Lemaur, V.; Cornil, J.; Lazzaroni, R.; Duroux, J.L.; Olivier, Y.; Trouillas, P. Free radical scavenging by natural polyphenols: Atom versus electron transfer. J. Phys. Chem. A 2013, 117, 2082–2092. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, V.; Tuli, H.S.; Varol, A.; Thakral, F.; Yerer, M.B.; Sak, K.; Varol, M.; Jain, A.; Khan, M.A.; Sethi, G. Role of reactive oxygen species in cancer progression: Molecular mechanisms and recent advancements. Biomolecules 2019, 9, 735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guedes, É.A.C.; Da Silva, T.G.; Aguiar, J.S.; De Barros, L.D.; Pinotti, L.M.; Sant’Ana, A.E.G. Cytotoxic activity of marine algae against cancerous cells. Brazilian J. Pharmacogn. 2013, 23, 668–673. [Google Scholar] [CrossRef] [Green Version]
- Abu-Khudir, R.; Ismail, G.A.; Diab, T. Antimicrobial, Antioxidant, and Anti-Tumor Activities of Sargassum linearifolium and Cystoseira crinita from Egyptian Mediterranean Coast. Nutr. Cancer 2021, 73, 829–844. [Google Scholar] [CrossRef]
- Namvar, F.; Baharara, J.; Mahdi, A.A. Antioxidant and anticancer activities of selected Persian gulf algae. Indian J. Clin. Biochem. 2014, 29, 13–20. [Google Scholar] [CrossRef] [Green Version]
- Mhadhebi, L.; Mhadhebi, A.; Robert, J.; Bouraoui, A. Antioxidant, anti-inflammatory and antiproliferative effects of aqueous extracts of three Mediterranean brown seaweeds of the Genus Cystoseira. Iran. J. Pharm. Res. 2014, 13, 207–220. [Google Scholar] [CrossRef]
- Premarathna, A.D.; Ranahewa, T.H.; Wijesekera, S.K.; Harishchandra, D.L.; Karunathilake, K.J.K.; Waduge, R.N.; Wijesundara, R.R.M.K.K.; Jayasooriya, A.P.; Wijewardana, V.; Rajapakse, R.P.V.J. Preliminary screening of the aqueous extracts of twenty-three different seaweed species in Sri Lanka with in-vitro and in-vivo assays. Heliyon 2020, 6, e03918. [Google Scholar] [CrossRef]
- Abdelhamid, A.; Lajili, S.; Elkaibi, M.A.; Ben Salem, Y.; Abdelhamid, A.; Muller, C.D.; Majdoub, H.; Kraiem, J.; Bouraoui, A. Optimized Extraction, Preliminary Characterization and Evaluation of the in Vitro Anticancer Activity of Phlorotannin-Rich Fraction from the Brown Seaweed, Cystoseira sedoides. J. Aquat. Food Prod. Technol. 2019, 28, 892–909. [Google Scholar] [CrossRef]
- Zenthoefer, M.; Geisen, U.; Hofmann-Peiker, K.; Fuhrmann, M.; Kerber, J.; Kirchhöfer, R.; Hennig, S.; Peipp, M.; Geyer, R.; Piker, L.; et al. Isolation of polyphenols with anticancer activity from the Baltic Sea brown seaweed Fucus vesiculosus using bioassay-guided fractionation. J. Appl. Phycol. 2017, 29, 2021–2037. [Google Scholar] [CrossRef]
- Park, C.; Lee, H.; Hwangbo, H.; Ji, S.Y.; Kim, M.Y.; Kim, S.Y.; Hong, S.H.; Kim, G.Y.; Choi, Y.H. Ethanol extract of Hizikia fusiforme induces apoptosis in B16F10 mouse melanoma cells through ROS-dependent inhibition of the PI3K/Akt signaling pathway. Asian Pacific J. Cancer Prev. 2020, 21, 1275–1282. [Google Scholar] [CrossRef]
- Sevimli-Gur, C.; Yesil-Celiktas, O. Cytotoxicity screening of supercritical fluid extracted seaweeds and phenylpropanoids. Mol. Biol. Rep. 2019, 46, 3691–3699. [Google Scholar] [CrossRef]
- Kosanić, M.; Ranković, B.; Stanojković, T. Brown macroalgae from the Adriatic Sea as a promising source of bioactive nutrients. J. Food Meas. Charact. 2019, 13, 330–338. [Google Scholar] [CrossRef]
- Rosa, G.P.; Tavares, W.R.; Sousa, P.M.C.; Pagès, A.K.; Seca, A.M.L.; Pinto, D.C.G.A. Seaweed secondary metabolites with beneficial health effects: An overview of successes in in vivo studies and clinical trials. Mar. Drugs 2020, 18, 8. [Google Scholar] [CrossRef] [Green Version]
- Imbs, T.I.; Zvyagintseva, T.N. Phlorotannins are Polyphenolic Metabolites of Brown Algae. Russ. J. Mar. Biol. 2018, 44, 263–273. [Google Scholar] [CrossRef]
- Erpel, F.; Mateos, R.; Pérez-Jiménez, J.; Pérez-Correa, J.R. Phlorotannins: From isolation and structural characterization, to the evaluation of their antidiabetic and anticancer potential. Food Res. Int. 2020, 137, 109589. [Google Scholar] [CrossRef]
- Singh, I.P.; Sidana, J. Phlorotannins. In Functional Ingredients from Algae for Foods and Nutraceuticals; Domínguez, H., Ed.; Woodhead Publishing: Cambridge, UK, 2013; pp. 181–204. ISBN 9780857095121. [Google Scholar]
- Nahvi, I.; Belkahla, S.; Asiri, S.M.; Rehman, S. Overview and Prospectus of Algal Biogenesis of Nanoparticles. In Microbial Nanotechnology: Green Synthesis and Applications; Ansari, M.A., Rehman, S., Eds.; Springer: Singapore, 2021; pp. 121–134. ISBN 978-981-16-1923-6. [Google Scholar]
- Bai, Y.; Sun, Y.; Gu, Y.; Zheng, J.; Yu, C.; Qi, H. Preparation, Characterization and Antioxidant Activities of Kelp Phlorotannin Nanoparticles. Molecules 2020, 25, 4550. [Google Scholar] [CrossRef]
- Kaushalya, K.G.D.; Gunathilake, K.D.P.P. Encapsulation of phlorotannins from edible brown seaweed in chitosan: Effect of fortification on bioactivity and stability in functional foods. Food Chem. 2022, 377, 132012. [Google Scholar] [CrossRef]
- Singh, I.P.; Sidana, J.; Bharate, S.B.; Foley, W.J. Phloroglucinol compounds of natural origin: Synthetic aspects. Nat. Prod. Rep. 2010, 27, 393–416. [Google Scholar] [CrossRef]
- Pal Singh, I.; Bharate, S.B. Phloroglucinol compounds of natural origin. Nat. Prod. Rep. 2006, 23, 558–591. [Google Scholar] [CrossRef]
- Kang, K.A.; Zhang, R.; Chae, S.; Lee, S.J.; Kim, J.; Kim, J.; Jeong, J.; Lee, J.; Shin, T.; Lee, N.H.; et al. Phloroglucinol (1,3,5-trihydroxybenzene) protects against ionizing radiation-induced cell damage through inhibition of oxidative stress in vitro and in vivo. Chem. Biol. Interact. 2010, 185, 215–226. [Google Scholar] [CrossRef]
- Kwon, Y.H.; Jung, S.Y.; Kim, J.W.; Lee, S.H.; Lee, J.H.; Lee, B.Y.; Kwon, S.M. Phloroglucinol inhibits the bioactivities of endothelial progenitor cells and suppresses tumor angiogenesis in LLC-tumor-bearing mice. PLoS ONE 2012, 7, e33618. [Google Scholar] [CrossRef] [Green Version]
- Kim, R.K.; Suh, Y.; Yoo, K.C.; Cui, Y.H.; Hwang, E.; Kim, H.J.; Kang, J.S.; Kim, M.J.; Lee, Y.Y.; Lee, S.J. Phloroglucinol suppresses metastatic ability of breast cancer cells by inhibition of epithelial-mesenchymal cell transition. Cancer Sci. 2015, 106, 94–101. [Google Scholar] [CrossRef] [Green Version]
- Kang, M.H.; Kim, I.H.; Nam, T.J. Phloroglucinol induces apoptosis via apoptotic signaling pathways in HT-29 colon cancer cells. Oncol. Rep. 2014, 32, 1341–1346. [Google Scholar] [CrossRef] [Green Version]
- Kang, M.H.; Kim, I.H.; Nam, T.J. Phloroglucinol induces apoptosis through the regulation of insulin-like growth factor 1 receptor signaling pathways in human colon cancer HT-29 cells. Int. J. Oncol. 2014, 45, 1036–1042. [Google Scholar] [CrossRef] [Green Version]
- Kim, R.K.; Uddin, N.; Hyun, J.W.; Kim, C.; Suh, Y.; Lee, S.J. Novel anticancer activity of phloroglucinol against breast cancer stem-like cells. Toxicol. Appl. Pharmacol. 2015, 286, 143–150. [Google Scholar] [CrossRef]
- Lopes-Costa, E.; Abreu, M.; Gargiulo, D.; Rocha, E.; Ramos, A.A. Anticancer effects of seaweed compounds fucoxanthin and phloroglucinol, alone and in combination with 5-fluorouracil in colon cells. J. Toxicol. Environ. Health—Part A Curr. Issues 2017, 80, 1–12. [Google Scholar] [CrossRef]
- Kang, K.A.; Lee, K.H.; Chae, S.; Zhang, R.; Jung, M.S.; Lee, Y.; Kim, S.Y.; Kim, H.S.; Joo, H.G.; Park, J.W.; et al. Eckol isolated from Ecklonia cava attenuates oxidative stress induced cell damage in lung fibroblast cells. FEBS Lett. 2005, 579, 6295–6304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, E.; Ahn, G.N.; Lee, N.H.; Kim, J.M.; Yun, J.S.; Hyun, J.W.; Jeon, Y.J.; Wie, M.B.; Lee, Y.J.; Park, J.W.; et al. Radioprotective properties of eckol against ionizing radiation in mice. FEBS Lett. 2008, 582, 925–930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hyun, K.H.; Yoon, C.H.; Kim, R.K.; Lim, E.J.; An, S.; Park, M.J.; Hyun, J.W.; Suh, Y.; Kim, M.J.; Lee, S.J. Eckol suppresses maintenance of stemness and malignancies in glioma stem-like cells. Toxicol. Appl. Pharmacol. 2011, 254, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Zhou, W.; Zhao, S.; Li, S.; Yan, D.; Wang, J. Eckol inhibits Reg3A-induced proliferation of human SW1990 pancreatic cancer cells. Exp. Ther. Med. 2019, 18, 2825–2832. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Guo, J.; Hu, X.; Liu, J.; Li, S.; Wang, J. Eckol protects against acute experimental colitis in mice: Possible involvement of Reg3g. J. Funct. Foods 2020, 73, 104088. [Google Scholar] [CrossRef]
- Zhang, M.Y.; Guo, J.; Hu, X.M.; Zhao, S.Q.; Li, S.L.; Wang, J. An in vivo anti-tumor effect of eckol from marine brown algae by improving the immune response. Food Funct. 2019, 10, 4361–4371. [Google Scholar] [CrossRef]
- Cho, S.H.; Kim, H.S.; Lee, W.W.; Han, E.J.; Kim, S.Y.; Fernando, I.P.S.; Ahn, G.; Kim, K.N. Eckol from Ecklonia cava ameliorates TNF-α/IFN-γ-induced inflammatory responses via regulating MAPKs and NF-κB signaling pathway in HaCaT cells. Int. Immunopharmacol. 2020, 82, 106146. [Google Scholar] [CrossRef]
- Eo, H.J.; Kwon, T.H.; Park, G.H.; Song, H.M.; Lee, S.J.; Park, N.H.; Jeong, J.B. In vitro anticancer activity of phlorofucofuroeckol a via upregulation of activating transcription factor 3 against human colorectal cancer cells. Mar. Drugs 2016, 14, 69. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.J.; Park, J.H.; Park, S.A.; Joo, N.R.; Lee, B.H.; Lee, K.B.; Oh, S.M. Dieckol or phlorofucofuroeckol extracted from Ecklonia cava suppresses lipopolysaccharide-mediated human breast cancer cell migration and invasion. J. Appl. Phycol. 2020, 32, 631–640. [Google Scholar] [CrossRef]
- Manandhar, B.; Wagle, A.; Seong, S.H.; Paudel, P.; Kim, H.R.; Jung, H.A.; Choi, J.S. Phlorotannins with potential anti-tyrosinase and antioxidant activity isolated from the marine seaweed Ecklonia stolonifera. Antioxidants 2019, 8, 240. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Li, Y.; Qian, Z.J.; Lee, S.H.; Li, Y.X.; Kim, S. Dieckol from Ecklonia cava regulates invasion of human fibrosarcoma cells and modulates mmp-2 and mmp-9 expression via NF-b pathway. Evid.-Based Complement. Altern. Med. 2011, 2011, 140462. [Google Scholar] [CrossRef] [Green Version]
- Oh, S.M.; Park, C.G.; Kang, J.H.; Kim, E.J.; Chee, H.Y.; Lee, B.H.; Lee, K.B. Dieckol inhibits 12-O-tetradecanoylphorbol-13-acetate-induced SK-Hep1 human hepatoma cell motility through suppression of matrix metalloproteinase-9 activity. J. Appl. Biol. Chem. 2011, 54, 376–381. [Google Scholar] [CrossRef]
- Wang, L.; Lee, W.; Jayawardena, T.U.; Cha, S.-H.; Jeon, Y.-J. Dieckol, an algae-derived phenolic compound, suppresses airborne particulate matter-induced skin aging by inhibiting the expressions of pro-inflammatory cytokines and matrix metalloproteinases through regulating NF-κB, AP-1, and MAPKs signaling pathways. Food Chem. Toxicol. 2020, 146, 111823. [Google Scholar] [CrossRef]
- Park, S.J.; Kim, Y.T.; Jeon, Y.J. Antioxidant dieckol downregulates the Rac1/ROS signaling pathway and inhibits Wiskott-Aldrich syndrome protein (WASP)-family verprolin-homologous protein 2 (WAVE2)-mediated invasive migration of B16 mouse melanoma cells. Mol. Cells 2012, 33, 363–369. [Google Scholar] [CrossRef] [Green Version]
- Park, S.J.; Jeon, Y.J. Dieckol from Ecklonia cava suppresses the migration and invasion of HT1080 cells by inhibiting the focal adhesion kinase pathway downstream of Rac1-ROS signaling. Mol. Cells 2012, 33, 141–149. [Google Scholar] [CrossRef] [Green Version]
- Takenawa, T.; Miki, H. WASP and WAVE family proteins: Key molecules for rapid rearrangement of cortical actin filaments and cell movement. J. Cell Sci. 2001, 114, 1801–1809. [Google Scholar] [CrossRef]
- Yoon, J.S.; Kasin Yadunandam, A.; Kim, S.J.; Woo, H.C.; Kim, H.R.; Kim, G. Do Dieckol, isolated from Ecklonia stolonifera, induces apoptosis in human hepatocellular carcinoma Hep3B cells. J. Nat. Med. 2013, 67, 519–527. [Google Scholar] [CrossRef]
- Li, Y.X.; Li, Y.; Je, J.Y.; Kim, S.K. Dieckol as a novel anti-proliferative and anti-angiogenic agent and computational anti-angiogenic activity evaluation. Environ. Toxicol. Pharmacol. 2015, 39, 259–270. [Google Scholar] [CrossRef]
- Ahn, J.H.; Yang, Y.I.; Lee, K.T.; Choi, J.H. Dieckol, isolated from the edible brown algae Ecklonia cava, induces apoptosis of ovarian cancer cells and inhibits tumor xenograft growth. J. Cancer Res. Clin. Oncol. 2014, 141, 255–268. [Google Scholar] [CrossRef]
- Kim, E.K.; Tang, Y.; Kim, Y.S.; Hwang, J.W.; Choi, E.J.; Lee, J.H.; Lee, S.H.; Jeon, Y.U.J.; Park, P.J. First evidence that Ecklonia cava-derived dieckol attenuates MCF-7 human breast carcinoma cell migration. Mar. Drugs 2015, 13, 1785–1797. [Google Scholar] [CrossRef] [Green Version]
- You, S.H.; Kim, J.-S.; Kim, Y.-S. Apoptosis and Cell Cycle Arrest in Two Human Breast Cancer Cell Lines by Dieckol Isolated from Ecklonia cava. J. Breast Dis. 2018, 6, 39–45. [Google Scholar] [CrossRef] [Green Version]
- Sadeeshkumar, V.; Duraikannu, A.; Ravichandran, S.; Kodisundaram, P.; Fredrick, W.S.; Gobalakrishnan, R. Modulatory efficacy of dieckol on xenobiotic-metabolizing enzymes, cell proliferation, apoptosis, invasion and angiogenesis during NDEA-induced rat hepatocarcinogenesis. Mol. Cell. Biochem. 2017, 433, 195–204. [Google Scholar] [CrossRef]
- Sadeeshkumar, V.; Duraikannu, A.; Ravichandran, S.; Fredrick, W.S.; Sivaperumal, R.; Kodisundaram, P. Protective effects of dieckol on N-nitrosodiethylamine induced hepatocarcinogenesis in rats. Biomed. Pharmacother. 2016, 84, 1810–1819. [Google Scholar] [CrossRef]
- Wang, C.H.; Li, X.F.; Jin, L.F.; Zhao, Y.; Zhu, G.J.; Shen, W.Z. Dieckol inhibits non-small–cell lung cancer cell proliferation and migration by regulating the PI3K/AKT signaling pathway. J. Biochem. Mol. Toxicol. 2019, 33, e22346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J.W.; Yan, Y.; Wang, L.; Wu, D.; Ye, N.K.; Chen, S.H.; Li, F. Marine bioactive compound dieckol induces apoptosis and inhibits the growth of human pancreatic cancer cells PANC-1. J. Biochem. Mol. Toxicol. 2021, 35, e22648. [Google Scholar] [CrossRef] [PubMed]
- Park, E.; Ahn, G.; Yun, J.S.; Kim, M.J.; Bing, S.J.; Kim, D.S.; Lee, J.; Lee, N.H.; Park, J.W.; Jee, Y. Dieckol rescues mice from lethal irradiation by accelerating hemopoiesis and curtailing immunosuppression. Int. J. Radiat. Biol. 2010, 86, 848–859. [Google Scholar] [CrossRef] [PubMed]
- Sadeeshkumar, V.; Duraikannu, A.; Aishwarya, T.; Jayaram, P.; Ravichandran, S.; Ganeshamurthy, R. Radioprotective efficacy of dieckol against gamma radiation-induced cellular damage in hepatocyte cells. Naunyn. Schmiedebergs. Arch. Pharmacol. 2019, 392, 1031–1041. [Google Scholar] [CrossRef] [PubMed]
- Toume, K.; Miyata, M.; Egawa, K.; Nose, K.; Hayashi, M.; Komiyama, K.; Ishibashi, M. Isolation of diphlorethohydroxycarmalol from a brown alga Ishige okamurae. Nat. Med. 2004, 58, 79–80. [Google Scholar]
- Kang, S.M.; Kim, A.D.; Heo, S.J.; Kim, K.N.; Lee, S.H.; Ko, S.C.; Jeon, Y.J. Induction of apoptosis by diphlorethohydroxycarmalol isolated from brown alga, Ishige okamurae. J. Funct. Foods 2012, 4, 433–439. [Google Scholar] [CrossRef]
- Park, M.H.; Jeon, Y.J.; Kim, H.J.; Han, J.S. Effect of diphlorethohydroxycarmalol isolated from Ishige okamurae on apoptosis in 3 t3-L1 preadipocytes. Phyther. Res. 2013, 27, 931–936. [Google Scholar] [CrossRef]
- Piao, M.J.; Kumara, M.H.S.R.; Kim, K.C.; Kang, K.A.; Kang, H.K.; Lee, N.H.; Hyun, J.W. Diphlorethohydroxycarmalol suppresses ultraviolet B-induced matrix metalloproteinases via inhibition of JNK and ERK signaling in human keratinocytes. Biomol. Ther. 2015, 23, 557–563. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Kim, H.S.; Oh, J.Y.; Je, J.G.; Jeon, Y.J.; Ryu, B.M. Protective effect of diphlorethohydroxycarmalol isolated from Ishige okamurae against UVB-induced damage in vitro in human dermal fibroblasts and in vivo in zebrafish. Food Chem. Toxicol. 2020, 136, 110963. [Google Scholar] [CrossRef]
- Piao, M.J.; Hewage, S.R.K.M.; Han, X.; Kang, K.A.; Kang, H.K.; Lee, N.H.; Hyun, J.W. Protective effect of diphlorethohydroxycarmalol against ultraviolet B radiation-induced DNA damage by inducing the nucleotide excision repair system in HaCaT human keratinocytes. Mar. Drugs 2015, 13, 5629–5641. [Google Scholar] [CrossRef] [Green Version]
- Park, C.; Lee, H.; Hong, S.H.; Kim, J.H.; Park, S.K.; Jeong, J.W.; Kim, G.Y.; Hyun, J.W.; Yun, S.J.; Kim, B.W.; et al. Protective effect of diphlorethohydroxycarmalol against oxidative stress-induced DNA damage and apoptosis in retinal pigment epithelial cells. Cutan. Ocul. Toxicol. 2019, 38, 298–308. [Google Scholar] [CrossRef]
- Kong, C.S.; Kim, J.A.; Yoon, N.Y.; Kim, S.K. Induction of apoptosis by phloroglucinol derivative from Ecklonia cava in MCF-7 human breast cancer cells. Food Chem. Toxicol. 2009, 47, 1653–1658. [Google Scholar] [CrossRef]
- Yoon, N.Y.; Eom, T.K.; Kim, M.M.; Kim, S.K. Inhibitory effect of phlorotannins isolated from Ecklonia cava on mushroom tyrosinase activity and melanin formation in mouse B16F10 melanoma cells. J. Agric. Food Chem. 2009, 57, 4124–4129. [Google Scholar] [CrossRef]
- Lee, M.S.; Yoon, H.D.; Kim, J.I.; Choi, J.S.; Byun, D.S.; Kim, H.R. Dioxinodehydroeckol inhibits melanin synthesis through PI3K/Akt signalling pathway in α-melanocyte-stimulating hormone-treated B16F10 cells. Exp. Dermatol. 2012, 21, 417–473. [Google Scholar] [CrossRef] [Green Version]
- Ryu, B.M.; Ahn, B.N.; Kang, K.H.; Kim, Y.S.; Li, Y.X.; Kong, C.S.; Kim, S.K.; Kim, D.G. Dioxinodehydroeckol protects human keratinocyte cells from UVB-induced apoptosis modulated by related genes Bax/Bcl-2 and caspase pathway. J. Photochem. Photobiol. B Biol. 2015, 153, 352–357. [Google Scholar] [CrossRef]
- Ham, Y.-M.; Baik, J.-S.; Hyun, J.-W.; Lee, N.-H. Isolation of a new phlorotannin, fucodiphlorethol G, from a brown alga Ecklonia cava. Bull. Korean Chem. Soc. 2007, 28, 1595–1597. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Qian, Z.J.; Kim, M.M.; Kim, S.K. Cytotoxic activities of phlorethol and fucophlorethol derivatives isolated from Laminariaceae Ecklonia cava. J. Food Biochem. 2011, 35, 357–369. [Google Scholar] [CrossRef]
- Li, Y.; Qian, Z.J.; Ryu, B.M.; Lee, S.H.; Kim, M.M.; Kim, S.K. Chemical components and its antioxidant properties in vitro: An edible marine brown alga, Ecklonia cava. Bioorganic Med. Chem. 2009, 17, 1963–1973. [Google Scholar] [CrossRef]
- Li, Y.X.; Li, Y.; Qian, Z.J.; Ryu, B.; Kim, S.K. Suppression of vascular endothelial growth factor (VEGF) induced angiogenic responses by fucodiphloroethol G. Process Biochem. 2011, 46, 1095–1103. [Google Scholar] [CrossRef]
- Gribble, G.W. Biological activity of recently discovered halogenated marine natural products. Mar. Drugs 2015, 13, 4044–4136. [Google Scholar] [CrossRef] [Green Version]
- Cavallo, G.; Metrangolo, P.; Milani, R.; Pilati, T.; Priimagi, A.; Resnati, G.; Terraneo, G. The halogen bond. Chem. Rev. 2016, 116, 2478–2601. [Google Scholar] [CrossRef] [Green Version]
- Mantri, R.V.; Sanghvi, R.; Zhu, H.J. Solubility of pharmaceutical solids. In Developing Solid Oral Dosage Forms: Pharmaceutical Theory and Practice, 2nd ed.; Qiu, Y., Chen, Y., Zhang, G.G.Z., Yu, L., Mantri, R.V., Eds.; Elsevier Inc.: Amsterdam, The Netherlands, 2017; pp. 3–22. ISBN 9780128024478. [Google Scholar]
- Shibata, T.; Miyasaki, T.; Miyake, H.; Tanaka, R.; Kawaguchi, S. The Influence of Phlorotannins and Bromophenols on the Feeding Behavior of Marine Herbivorous Gastropod Turbo cornutus. Am. J. Plant Sci. 2014, 5, 387–392. [Google Scholar] [CrossRef] [Green Version]
- Dong, H.; Dong, S.; Hansen, P.E.; Stagos, D.; Lin, X.; Liu, M. Progress of bromophenols in marine algae from 2011 to 2020: Structure, bioactivities, and applications. Mar. Drugs 2020, 18, 411. [Google Scholar] [CrossRef]
- Shinada, N.K.; De Brevern, A.G.; Schmidtke, P. Halogens in Protein-Ligand Binding Mechanism: A Structural Perspective. J. Med. Chem. 2019, 62, 9341–9356. [Google Scholar] [CrossRef]
- Xu, X.; Song, F.; Wang, S.; Li, S.; Xiao, F.; Zhao, J.; Yang, Y.; Shang, S.; Yang, L.; Shi, J. Dibenzyl bromophenols with diverse dimerization patterns from the brown alga Leathesia nana. J. Nat. Prod. 2004, 67, 1661–1666. [Google Scholar] [CrossRef]
- Shi, D.; Li, J.; Guo, S.; Su, H.; Fan, X. The antitumor effect of bromophenol derivatives in vitro and Leathesia nana extract in vivo. Chinese J. Oceanol. Limnol. 2009, 27, 277–282. [Google Scholar] [CrossRef]
- Lijun, H.; Nianjun, X.; Jiangong, S.; Xiaojun, Y.; Chengkui, Z. Isolation and pharmacological activities of bromophenols from Rhodomela confervoides. Chinese J. Oceanol. Limnol. 2005, 23, 226–229. [Google Scholar] [CrossRef]
- Liu, M.; Zhang, W.; Wei, J.; Lin, X. Synthesis and α-glucosidase inhibitory mechanisms of bis(2,3-dibromo-4,5-dihydroxybenzyl) ether, a potential marine bromophenol α-glucosidase inhibitor. Mar. Drugs 2011, 9, 1554–1565. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Zhang, W.; Wei, J.; Qiu, L.; Lin, X. Marine bromophenol bis(2,3-dibromo-4,5-dihydroxybenzyl) ether, induces mitochondrial apoptosis in K562 cells and inhibits topoisomerase I in vitro. Toxicol. Lett. 2012, 211, 126–134. [Google Scholar] [CrossRef]
- Liu, M.; Wang, G.; Xiao, L.; Xu, A.; Liu, X.; Xu, P.; Lin, X. Bis(2,3-dibromo-4,5-dihydroxybenzyl) ether, a marine algae derived bromophenol, inhibits the growth of Botrytis cinerea and interacts with DNA molecules. Mar. Drugs 2014, 12, 3838–3851. [Google Scholar] [CrossRef]
- Qi, X.; Liu, G.; Qiu, L.; Lin, X.; Liu, M. Marine bromophenol bis(2,3-dibromo-4,5-dihydroxybenzyl) ether, represses angiogenesis in HUVEC cells and in zebrafish embryos via inhibiting the VEGF signal systems. Biomed. Pharmacother. 2015, 75, 58–66. [Google Scholar] [CrossRef] [PubMed]
- Wu, N.; Luo, J.; Jiang, B.; Wang, L.; Wang, S.; Wang, C.; Fu, C.; Li, J.; Shi, D. Marine bromophenol bis (2,3-dibromo-4,5-dihydroxy-phenyl)-methane inhibits the proliferation, migration, and invasion of hepatocellular carcinoma cells via modulating β1-integrin/FAK signaling. Mar. Drugs 2015, 13, 1010–1025. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Wang, L.J.; Jiang, B.; Wu, N.; Li, X.; Liu, S.; Luo, J.; Shi, D. Anti-angiogenic properties of BDDPM, a bromophenol from marine red alga Rhodomela confervoides, with multi receptor tyrosine kinase inhibition effects. Int. J. Mol. Sci. 2015, 16, 13548–13560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hodgkin, J.H.; Craigie, J.S.; McInnes, A.G. The occurrence of 2,3-dibromobenzyl alcohol 4,5-disulfate, dipotassium salt, in Polysiphonia lanosa. Can. J. Chem. 1966, 44, 74–78. [Google Scholar] [CrossRef] [Green Version]
- Shoeib, N.A.; Bibby, M.C.; Blunden, G.; Linley, P.A.; Swaine, D.J.; Wheelhouse, R.T.; Wright, C.W. In-vitro cytotoxic activities of the major bromophenols of the red alga Polysiphonia lanosa and some novel synthetic isomers. J. Nat. Prod. 2004, 67, 1445–1449. [Google Scholar] [CrossRef] [PubMed]
- Popplewell, W.L.; Northcote, P.T. Colensolide A: A new nitrogenous bromophenol from the New Zealand marine red alga Osmundaria colensoi. Tetrahedron Lett. 2009, 50, 6814–6817. [Google Scholar] [CrossRef]
- Ma, M.; Zhao, J.; Wang, S.; Li, S.; Yang, Y.; Shi, J.; Fan, X.; He, L. Bromophenols coupled with methyl γ-ureidobutyrate and bromophenol sulfates from the red alga Rhodomela confervoides. J. Nat. Prod. 2006, 69, 206–210. [Google Scholar] [CrossRef]
- Colon, M.; Guevara, P.; Gerwick, W.H.; Ballantine, D. 5’-hydroxyisoavrainvilleol, a new Diphenylmethane Derivative from the Tropical Green Alga Avrainvillea nigricans. J. Nat. Prod. 1987, 50, 368–374. [Google Scholar] [CrossRef]
- Hawas, U.W.; Abou El-Kassem, L.T.; Al-Farawati, R.; Shaher, F.M. Halo-phenolic metabolites and their in vitro antioxidant and cytotoxic activities from the Red Sea alga Avrainvillea amadelpha. Zeitschrift fur Naturforsch. 2021, 76, 213–218. [Google Scholar] [CrossRef]
- Wegener, A.; Miller, K.A. Total Synthesis of Avrainvilleol. J. Org. Chem. 2017, 82, 11655–11658. [Google Scholar] [CrossRef]
- Tauchen, J.; Huml, L.; Rimpelova, S.; Jurášek, M. Flavonoids and related members of the aromatic polyketide group in human health and disease: Do they really work? Molecules 2020, 25, 3846. [Google Scholar] [CrossRef]
- Martins, B.T.; Correia da Silva, M.; Pinto, M.; Cidade, H.; Kijjoa, A. Marine natural flavonoids: Chemistry and biological activities. Nat. Prod. Res. 2019, 33, 3260–3272. [Google Scholar] [CrossRef]
- Goiris, K.; Muylaert, K.; Voorspoels, S.; Noten, B.; De Paepe, D.; E Baart, G.J.; De Cooman, L. Detection of flavonoids in microalgae from different evolutionary lineages. J. Phycol. 2014, 50, 483–492. [Google Scholar] [CrossRef]
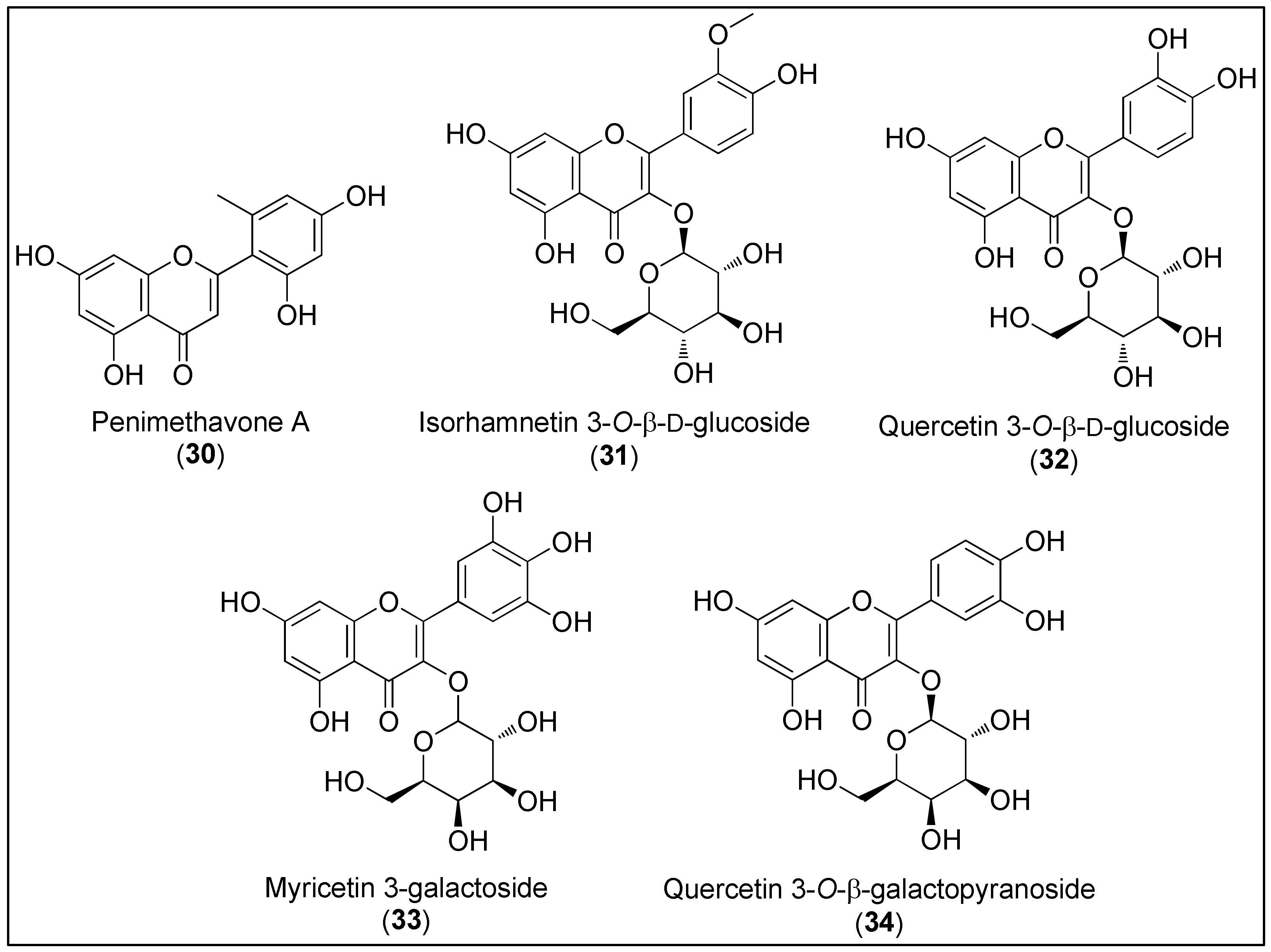
- Hou, X.M.; Wang, C.Y.; Gu, Y.C.; Shao, C.L. Penimethavone A, a flavone from a gorgonian-derived fungus Penicillium chrysogenum. Nat. Prod. Res. 2016, 30, 2274–2277. [Google Scholar] [CrossRef]
- Kong, C.S.; Kim, Y.A.; Kim, M.M.; Park, J.S.; Kim, J.A.; Kim, S.K.; Lee, B.J.; Nam, T.J.; Seo, Y. Flavonoid glycosides isolated from Salicornia herbacea inhibit matrix metalloproteinase in HT1080 cells. Toxicol. Vitr. 2008, 22, 1742–1748. [Google Scholar] [CrossRef]
- Mohammed, M.M.D.; El-Sharkawy, E.R.; Matloub, A.A. Cytotoxic flavonoids from Diplotaxis harra (Forssk.) Boiss. growing in Sinai. J. Med. Plant Res. 2011, 5, 5099–5103. [Google Scholar]
- Bae, M.J.; Karadeniz, F.; Oh, J.H.; Yu, G.H.; Jang, M.S.; Nam, K.H.; Seo, Y.; Kong, C.S. MMP-Inhibitory Effects of Flavonoid Glycosides from Edible Medicinal Halophyte Limonium tetragonum. Evid.-Based Complement. Altern. Med. 2017, 2017, 6750274. [Google Scholar] [CrossRef] [Green Version]
- Önder, A. Anticancer activity of natural coumarins for biological targets. In Studies in Natural Products Chemistry; Rahman, A., Ed.; Elsevier: Amsterdam, The Netherlands, 2020; Volume 64, pp. 85–109. ISBN 978-0-12-817903-1. [Google Scholar]
- Vazquez-Rodriguez, S.; Matos, M.; Borges, F.; Uriarte, E.; Santana, L. Bioactive Coumarins from Marine Sources: Origin, Structural Features and Pharmacological Properties. Curr. Top. Med. Chem. 2015, 15, 1755–1766. [Google Scholar] [CrossRef]
- Matos, M.J.; Santana, L.; Uriarte, E.; Abreu, O.A.; Molina, E.; Yordi, E.G. Coumarins—An Important Class of Phytochemicals. In Phytochemicals—Isolation, Characterisation and Role in Human Health; Rao, A.V., Rao, L.G., Eds.; IntechOpen: London, UK, 2015; pp. 113–140. ISBN 978-953-51-2170-1. [Google Scholar]
- Shejwalkar, P. Applications of Coumarins as Cardiovascular and Anti-Cancer Agents: A Short Review. J. Cardiol. Cardiovasc. Ther. 2017, 8, 1–6. [Google Scholar] [CrossRef]
- Katsori, A.M.; Hadjipavlou-Litina, D. Coumarin derivatives: An updated patent review (2012–2014). Expert Opin. Ther. Pat. 2014, 24, 1323–1347. [Google Scholar] [CrossRef]
- Detsi, A.; Kontogiorgis, C.; Hadjipavlou-Litina, D. Coumarin derivatives: An updated patent review (2015–2016). Expert Opin. Ther. Pat. 2017, 27, 1201–1226. [Google Scholar] [CrossRef]
- Tan, N.; Tao, Y.; Pan, J.; Wang, S.; Xu, F.; She, Z.; Lin, Y.; Gareth Jones, E.B. Isolation, structure elucidation, and mutagenicity of four alternariol derivatives produced by the mangrove endophytic fungus No. 2240. Chem. Nat. Compd. 2008, 44, 296–300. [Google Scholar] [CrossRef]
- Hawas, U.W.; El-Desouky, S.; Abou El-Kassem, L.; Elkhateeb, W. Alternariol derivatives from Alternaria alternata, an endophytic fungus residing in Red sea soft coral, inhibit HCV NS3/4A protease. Appl. Biochem. Microbiol. 2015, 51, 579–584. [Google Scholar] [CrossRef]
- Fukuda, T.; Ishibashi, F.; Iwao, M. Lamellarin alkaloids: Isolation, synthesis, and biological activity. In Alkaloids: Chemistry and Biology; Knölker, H.-J., Ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2020; Volume 83, pp. 1–112. ISBN 9780128209813. [Google Scholar]
- Bailly, C. Anticancer properties of lamellarins. Mar. Drugs 2015, 13, 1105–1123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Facompré, M.; Tardy, C.; Bal-Mahieu, C.; Colson, P.; Perez, C.; Manzanares, I.; Cuevas, C.; Bailly, C. Lamellarin D: A Novel Potent Inhibitor of Topoisomerase I. Cancer Res. 2003, 63, 7392–7399. [Google Scholar] [PubMed]
- Ballot, C.; Kluza, J.; Martoriati, A.; Nyman, U.; Formstecher, P.; Joseph, B.; Bailly, C.; Marchetti, P. Essential role of mitochondria in apoptosis of cancer cells induced by the marine alkaloid Lamellarin D. Mol. Cancer Ther. 2009, 8, 3307–3317. [Google Scholar] [CrossRef] [Green Version]
- Ballot, C.; Martoriati, A.; Jendoubi, M.; Buche, S.; Formstecher, P.; Mortier, L.; Kluza, J.; Marchetti, P. Another facet to the anticancer response to lamellarin D: Induction of cellular senescence through inhibition of topoisomerase i and intracellular ROS production. Mar. Drugs 2014, 12, 779–798. [Google Scholar] [CrossRef] [Green Version]
- Ballot, C.; Kluza, J.; Lancel, S.; Martoriati, A.; Hassoun, S.M.; Mortier, L.; Vienne, J.C.; Briand, G.; Formstecher, P.; Bailly, C.; et al. Inhibition of mitochondrial respiration mediates apoptosis induced by the anti-tumoral alkaloid lamellarin D. Apoptosis 2010, 15, 769–781. [Google Scholar] [CrossRef]
- Colligs, V.; Hansen, S.P.; Imbri, D.; Seo, E.J.; Kadioglu, O.; Efferth, T.; Opatz, T. Synthesis and biological evaluation of a D-ring-contracted analogue of lamellarin D. Bioorganic Med. Chem. 2017, 25, 6137–6148. [Google Scholar] [CrossRef]
- Lade, D.M.; Pawar, A.B.; Mainkar, P.S.; Chandrasekhar, S. Total Synthesis of Lamellarin D Trimethyl Ether, Lamellarin D, and Lamellarin H. J. Org. Chem. 2017, 82, 4998–5004. [Google Scholar] [CrossRef]
- Quesada, A.R.; García Grávalos, M.D.; Fernández Puentes, J.L. Polyaromatic alkaloids from marine invertebrates as cytotoxic compounds and inhibitors of multidrug resistance caused by P-glycoprotein. Br. J. Cancer 1996, 74, 677–682. [Google Scholar] [CrossRef] [Green Version]
- Du, L.; Zhu, T.; Fang, Y.; Liu, H.; Gu, Q.; Zhu, W. Aspergiolide A, a novel anthraquinone derivative with naphtho[1,2,3-de]chromene-2,7-dione skeleton isolated from a marine-derived fungus Aspergillus glaucus. Tetrahedron 2007, 63, 1085–1088. [Google Scholar] [CrossRef]
- Wang, Y.; Qi, X.; Li, D.; Zhu, T.; Mo, X.; Li, J. Anticancer efficacy and absorption, distribution, metabolism, and toxicity studies of aspergiolide A in early drug development. Drug Des. Devel. Ther. 2014, 8, 1965–1977. [Google Scholar] [CrossRef] [Green Version]
- Du, L.; Zhu, T.; Liu, H.; Fang, Y.; Zhu, W.; Gu, Q. Cytotoxic polyketides from a marine-derived fungus Aspergillus glaucus. J. Nat. Prod. 2008, 71, 1837–1842. [Google Scholar] [CrossRef]
- Watty, M.; Syahdi, R.R.; Yanuar, A. Database compilation and virtual screening of secondary metabolites derived from marine fungi as epidermal growth factor receptor tyrosine kinase inhibitors. Asian J. Pharm. Clin. Res. 2017, 10, 142–147. [Google Scholar] [CrossRef] [Green Version]
- De La Mare, J.A.; Lawson, J.C.; Chiwakata, M.T.; Beukes, D.R.; Edkins, A.L.; Blatch, G.L. Quinones and halogenated monoterpenes of algal origin show anti-proliferative effects against breast cancer cells in vitro. Investig. New Drugs 2012, 30, 2187–2200. [Google Scholar] [CrossRef]
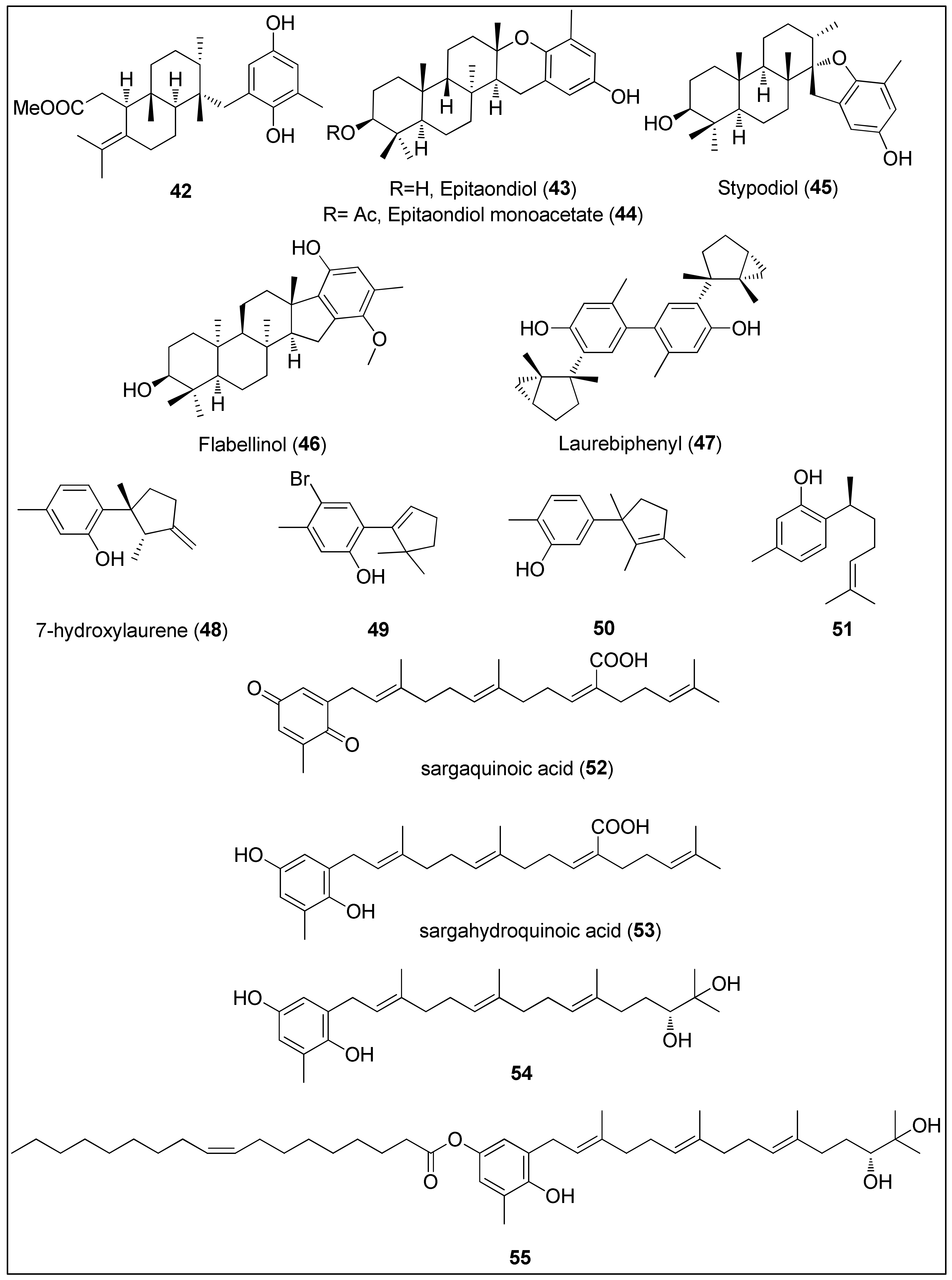
- Le Bideau, F.; Kousara, M.; Chen, L.; Wei, L.; Dumas, F. Tricyclic sesquiterpenes from marine origin. Chem. Rev. 2017, 117, 6110–6159. [Google Scholar] [CrossRef]
- Wright, A.E.; Pomponi, S.A.; McConnell, O.J.; Kohmoto, S.; McCarthy, P.J. (+)-Curcuphenol and (+)-curcudiol, sesquiterpene phenols from shallow and deep water collections of the marine sponge Didiscus flavus. J. Nat. Prod. 1987, 50, 976–978. [Google Scholar] [CrossRef]
- Dorta, E.; Cueto, M.; Brito, I.; Darias, J. New terpenoids from the brown alga Stypopodium zonale. J. Nat. Prod. 2002, 65, 1727–1730. [Google Scholar] [CrossRef]
- Pereira, D.M.; Cheel, J.; Areche, C.; San-Martin, A.; Rovirosa, J.; Silva, L.R.; Valentao, P.; Andrade, P.B. Anti-proliferative activity of meroditerpenoids isolated from the brown alga Stypopodium flabelliforme against several cancer cell lines. Mar. Drugs 2011, 9, 852–862. [Google Scholar] [CrossRef]
- Depix, M.S.; Martínez, J.; Santibañez, F.; Rovirosa, J.; Martín, A.S.; Maccioni, R.B. The compound 14-keto stypodiol diacetate from the algae Stypopodium flabelliforme inhibits microtubules and cell proliferation in DU-145 human prostatic cells. Mol. Cell. Biochem. 1998, 187, 191–199. [Google Scholar] [CrossRef]
- Sabry, O.M.M.; Andrews, S.; McPhail, K.L.; Goeger, D.E.; Yokochi, A.; LePage, K.T.; Murray, T.F.; Gerwick, W.H. Neurotoxic meroditerpenoids from the tropical marine brown alga Stypopodium flabelliforme. J. Nat. Prod. 2005, 68, 1022–1030. [Google Scholar] [CrossRef]
- Sun, J.; Shi, D.; Ma, M.; Li, S.; Wang, S.; Han, L.; Yang, Y.; Fan, X.; Shi, J.; He, L. Sesquiterpenes from the red alga Laurencia tristicha. J. Nat. Prod. 2005, 68, 915–919. [Google Scholar] [CrossRef] [PubMed]
- Shizuri, Y.; Yamada, K. Laurebiphenyl, a dimeric sesquiterpene of the cyclolaurane-type from the red alga Laurencia nidifica. Phytochemistry 1985, 24, 1385–1386. [Google Scholar] [CrossRef]
- Kladi, M.; Xenaki, H.; Vagias, C.; Papazafiri, P.; Roussis, V. New cytotoxic sesquiterpenes from the red algae Laurencia obtusa and Laurencia microcladia. Tetrahedron 2006, 62, 182–189. [Google Scholar] [CrossRef]
- Kladi, M.; Vagias, C.; Furnari, G.; Moreau, D.; Roussakis, C.; Roussis, V. Cytotoxic cuparene sesquiterpenes from Laurencia microcladia. Tetrahedron Lett. 2005, 46, 5723–5726. [Google Scholar] [CrossRef]
- Rodrigo, G.; Almanza, G.R.; Cheng, Y.; Peng, J.; Hamann, M.; Duan, R.D.; Åkesson, B. Antiproliferative effects of curcuphenol, a sesquiterpene phenol. Fitoterapia 2010, 81, 762–766. [Google Scholar] [CrossRef]
- Taşdemir, D.; Bugni, T.S.; Mangalindan, G.C.; Concepción, G.P.; Harper, M.K.; Ireland, C.M. Bisabolane type sesquiterpenes from a marine Didiscus sponge. Turkish J. Chem. 2003, 27, 273–279. [Google Scholar]
- Reddy, P.; Urban, S. Meroditerpenoids from the southern Australian marine brown alga Sargassum fallax. Phytochemistry 2009, 70, 250–255. [Google Scholar] [CrossRef]
- Hur, S.; Lee, H.; Kim, Y.; Lee, B.H.; Shin, J.; Kim, T.Y. Sargaquinoic acid and sargachromenol, extracts of Sargassum sagamianum, induce apoptosis in HaCaT cells and mice skin: Its potentiation of UVB-induced apoptosis. Eur. J. Pharmacol. 2008, 582, 1–11. [Google Scholar] [CrossRef]
- Mori, J.; Iwashima, M.; Wakasugi, H.; Saito, H.; Matsunaga, T.; Ogasawara, M.; Takahashi, S.; Suzuki, H.; Hayashi, T. New plastoquinones isolated from the brown alga, Sargassum micracanthum. Chem. Pharm. Bull. 2005, 53, 1159–1163. [Google Scholar] [CrossRef] [Green Version]
- Bertanha, C.S.; Januário, A.H.; Alvarenga, T.A.; Pimenta, L.P.; E Silva, M.L.A.; Cunha, W.R.; Pauletti, P.M. Quinone and hydroquinone metabolites from the ascidians of the genus Aplidium. Mar. Drugs 2014, 12, 3608–3633. [Google Scholar] [CrossRef] [Green Version]
- García, P.A.; Hernández, Á.P.; San Feliciano, A.; Castro, M.A.Á. Bioactive prenyl- and terpenyl-quinones/ Hydroquinones of marine origin. Mar. Drugs 2018, 16, 292. [Google Scholar] [CrossRef] [Green Version]
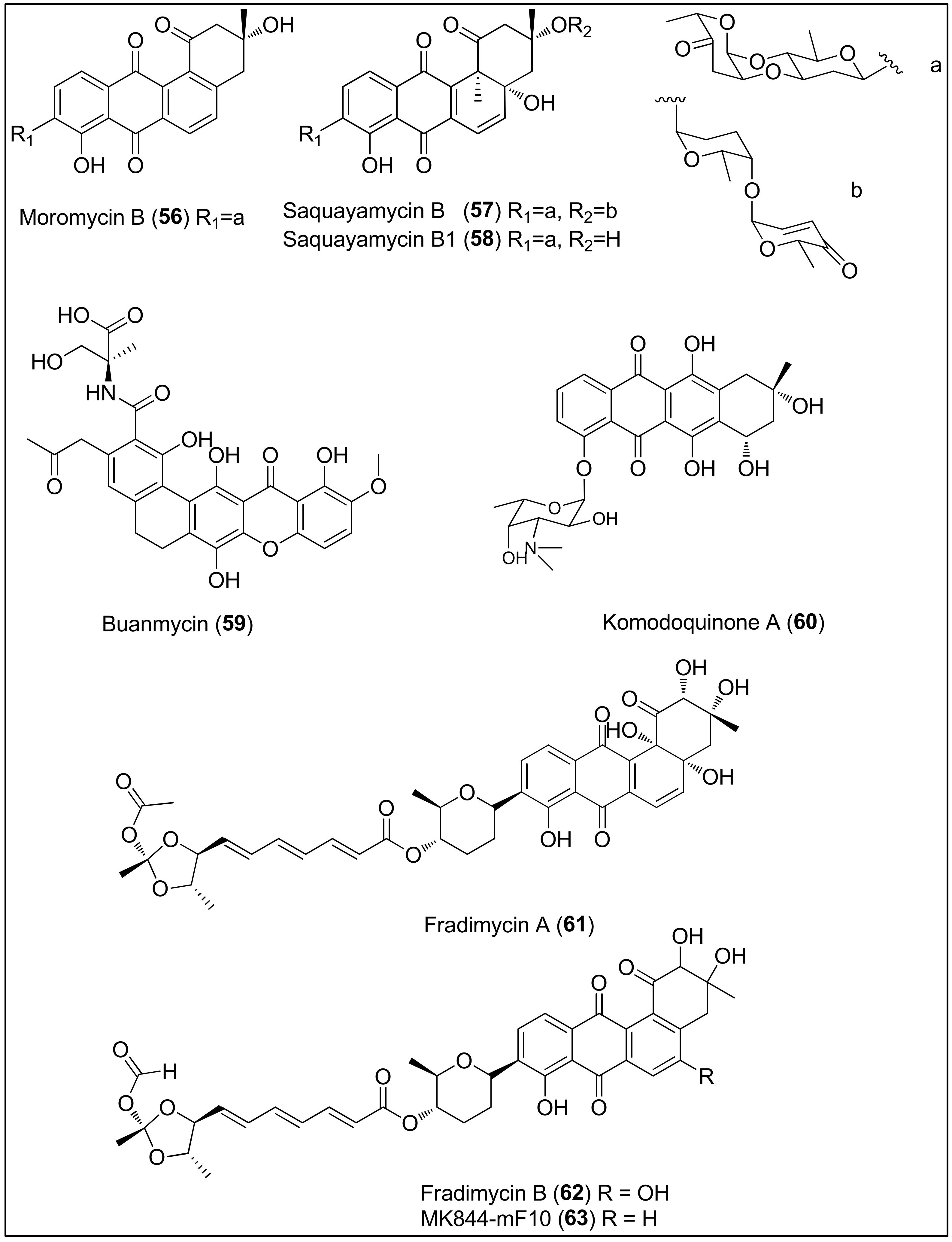
- Qu, X.Y.; Ren, J.W.; Peng, A.H.; Lin, S.Q.; Lu, D.D.; Du, Q.Q.; Liu, L.; Li, X.; Li, E.W.; Xie, W.D. Cytotoxic, anti-migration, and anti-invasion activities on breast cancer cells of angucycline glycosides isolated from a marine-derived Streptomyces sp. Mar. Drugs 2019, 17, 277. [Google Scholar] [CrossRef] [Green Version]
- Moon, K.; Chung, B.; Shin, Y.; Rheingold, A.L.; Moore, C.E.; Park, S.J.; Park, S.; Lee, S.K.; Oh, K.B.; Shin, J.; et al. Pentacyclic antibiotics from a tidal mud flat-derived actinomycete. J. Nat. Prod. 2015, 78, 524–529. [Google Scholar] [CrossRef]
- Itoh, T.; Kinoshita, M.; Wei, H.; Kobayashi, M. Stereostructure of komodoquinone A, a neuritogenic anthracycline, from marine Streptomyces sp. KS3. Chem. Pharm. Bull. 2003, 51, 1402–1404. [Google Scholar] [CrossRef] [Green Version]
- Itoh, T.; Kinoshita, M.; Aoki, S.; Kobayashi, M. Komodoquinone A, a Novel Neuritogenic Anthracycline, from Marine Streptomyces sp. KS3. J. Nat. Prod. 2003, 66, 1373–1377. [Google Scholar] [CrossRef]
- Xin, W.; Ye, X.; Yu, S.; Lian, X.Y.; Zhang, Z. New capoamycin-type antibiotics and polyene acids from marine Streptomyces fradiae PTZ0025. Mar. Drugs 2012, 10, 2388–2402. [Google Scholar] [CrossRef] [Green Version]
- Ganesan, S.; Velsamy, G.; Sivasudha, T.; Manoharan, N. MALDI-TOF mass spectrum profiling, antibacterial and anticancer activity of marine Streptomyces fradiae BDMS1. World J. Pharm. Pharm. Sci. 2013, 2, 5148–5165. [Google Scholar]
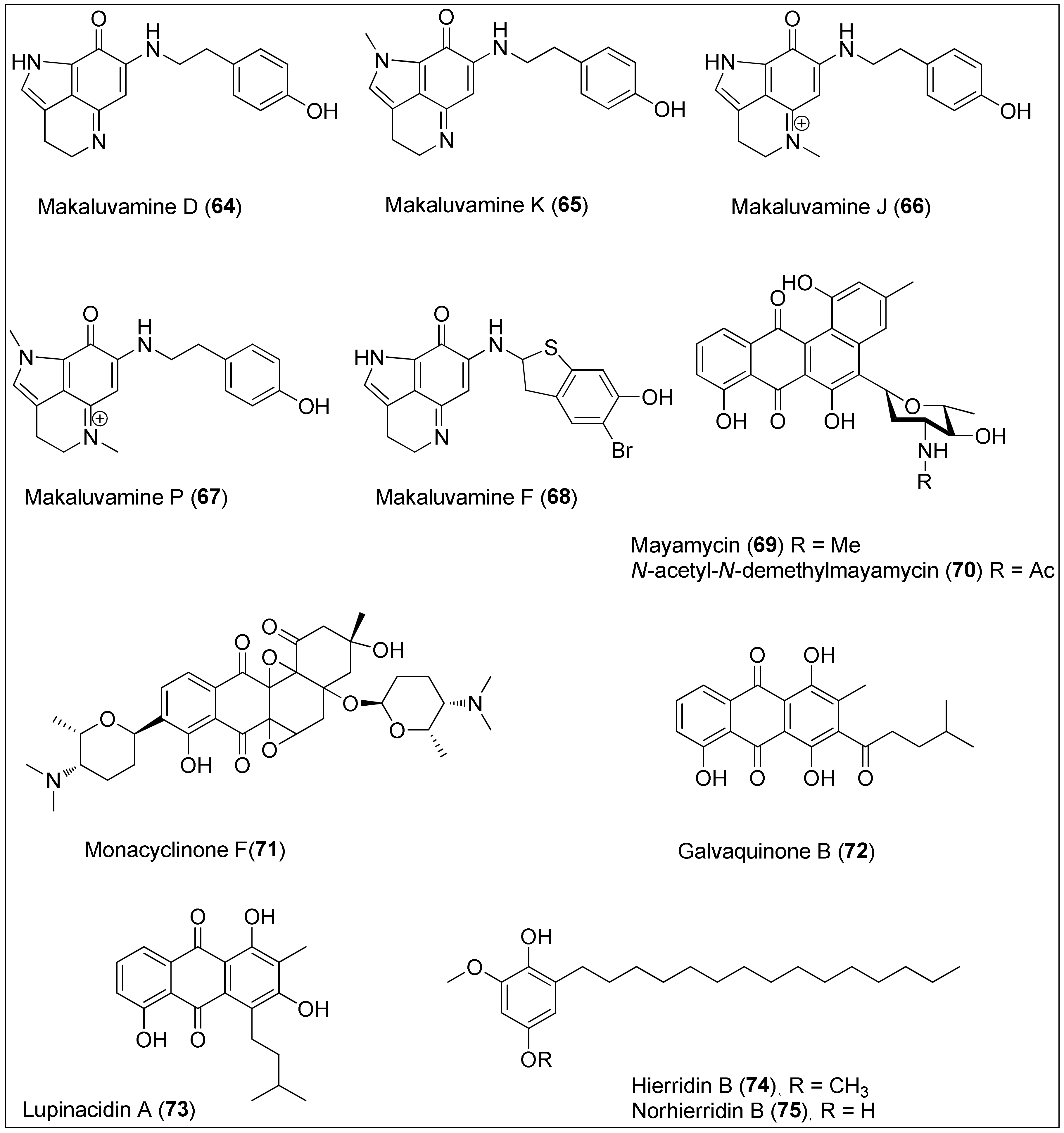
- Kita, Y.; Fujioka, H. Marine pyrroloiminoquinone alkaloids. In Topics in Current Chemistry; Springer: Berlin/Heidelberg, Germany, 2012; Volume 309, pp. 131–162. ISBN 9783642255281. [Google Scholar]
- Kalinski, J.C.J.; Krause, R.W.M.; Parker-Nance, S.; Waterworth, S.C.; Dorrington, R.A. Unlocking the Diversity of Pyrroloiminoquinones Produced by Latrunculid Sponge Species. Mar. Drugs 2021, 19, 68. [Google Scholar] [CrossRef]
- Lin, S.; McCauley, E.P.; Lorig-Roach, N.; Tenney, K.; Naphen, C.N.; Yang, A.M.; Johnson, T.A.; Hernadez, T.; Rattan, R.; Valeriote, F.A.; et al. Another look at pyrroloiminoquinone alkaloids-perspectives on their therapeutic potential from known structures and semisynthetic analogues. Mar. Drugs 2017, 15, 98. [Google Scholar] [CrossRef] [Green Version]
- Dijoux, M.G.; Schnabel, P.C.; Hallock, Y.F.; Boswell, J.L.; Johnson, T.R.; Wilson, J.A.; Ireland, C.M.; Van Soest, R.; Boyd, M.R.; Barrows, L.R.; et al. Antitumor activity and distribution of pyrroloiminoquinones in the sponge genus Zyzzya. Bioorganic Med. Chem. 2005, 13, 6035–6044. [Google Scholar] [CrossRef]
- Radisky, D.C.; Radisky, E.S.; Copp, B.R.; Ireland, C.M.; Barrows, L.R.; Kramer, R.A. Novel Cytotoxic Topoisomerase II Inhibiting Pyrroloiminoquinones from Fijian Sponges of the Genus Zyzzya. J. Am. Chem. Soc. 1993, 115, 1632–1638. [Google Scholar] [CrossRef]
- Goey, A.K.L.; Chau, C.H.; Sissung, T.M.; Cook, K.M.; Venzon, D.J.; Castro, A.; Ransom, T.R.; Henrich, C.J.; McKee, T.C.; McMahon, J.B.; et al. Screening and Biological Effects of Marine Pyrroloiminoquinone Alkaloids: Potential Inhibitors of the HIF-1α/p300 Interaction. J. Nat. Prod. 2016, 79, 1267–1275. [Google Scholar] [CrossRef] [PubMed]
- Aburjania, Z.; Whitt, J.D.; Jang, S.; Nadkarni, D.H.; Chen, H.; Rose, J.B.; Velu, S.E.; Jaskula-Sztul, R. Synthetic Makaluvamine Analogs Decrease c-Kit Expression and Are Cytotoxic to Neuroendocrine Tumor Cells. Molecules 2020, 25, 4940. [Google Scholar] [CrossRef] [PubMed]
- Boucle, S.; Melin, C.; Clastre, M.; Guillard, J. Design, synthesis and evaluation of new marine alkaloid-derived pentacyclic structures with anti-tumoral potency. Mar. Drugs 2015, 13, 655–665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Backenköhler, J.; Spindler, S.; Spiteller, P. Total Synthesis of Damirone C, Makaluvamine O, Makaluvone, Batzelline C and Batzelline D. ChemistrySelect 2017, 2, 2589–2592. [Google Scholar] [CrossRef]
- Schneemann, I.; Kajahn, I.; Ohlendorf, B.; Zinecker, H.; Erhard, A.; Nagel, K.; Wiese, J.; Imhoff, J.F. Mayamycin, a cytotoxic polyketide from a Streptomyces strain isolated from the marine sponge Halichondria panicea. J. Nat. Prod. 2010, 73, 1309–1312. [Google Scholar] [CrossRef]
- Liang, Y.; Xie, X.; Chen, L.; Yan, S.; Ye, X.; Anjum, K.; Huang, H.; Lian, X.; Zhang, Z. Bioactive Polycyclic Quinones from Marine Streptomyces sp. 182SMLY. Mar. Drugs 2016, 14, 10–20. [Google Scholar] [CrossRef] [Green Version]
- Wu, K.; Wang, M.; Yao, Q.; Zhang, A. Synthesis study toward mayamycin. Chinese J. Chem. 2013, 31, 93–99. [Google Scholar] [CrossRef]
- Chakraborty, S.; Mal, D. A Representative Synthetic Route for C5 Angucycline Glycosides: Studies Directed toward the Total Synthesis of Mayamycin. J. Org. Chem. 2018, 83, 1328–1339. [Google Scholar] [CrossRef]
- Vicente, J.; Stewart, A.K.; Van Wagoner, R.M.; Elliott, E.; Bourdelais, A.J.; Wright, J.L.C. Monacyclinones, new angucyclinone metabolites isolated from Streptomyces sp. M7-15 associated with the Puerto Rican Sponge Scopalina ruetzleri. Mar. Drugs 2015, 13, 4682–4700. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Martinez, E.D.; MacMillan, J.B. Anthraquinones from a marine-derived Streptomyces spinoverrucosus. J. Nat. Prod. 2012, 75, 1759–1764. [Google Scholar] [CrossRef] [Green Version]
- Sottorff, I.; Künzel, S.; Wiese, J.; Lipfert, M.; Preußke, N.; Sönnichsen, F.D.; Imhoff, J.F. Antitumor anthraquinones from an Easter island sea anemone: Animal or bacterial origin? Mar. Drugs 2019, 17, 154. [Google Scholar] [CrossRef] [Green Version]
- Papendorf, O.; König, G.M.; Wright, A.D. Hierridin B and 2,4-dimethoxy-6-heptadecyl-phenol, secondary metabolites from the cyanobacterium Phormidium ectocarpi with antiplasmodial activity. Phytochemistry 1998, 49, 2383–2386. [Google Scholar] [CrossRef]
- Leão, P.N.; Costa, M.; Ramos, V.; Pereira, A.R.; Fernandes, V.C.; Domingues, V.F.; Gerwick, W.H.; Vasconcelos, V.M.; Martins, R. Antitumor Activity of Hierridin B, a Cyanobacterial Secondary Metabolite Found in both Filamentous and Unicellular Marine Strains. PLoS ONE 2013, 8, e69562. [Google Scholar] [CrossRef] [Green Version]
- Freitas, S.; Martins, R.; Costa, M.; Leão, P.N.; Vitorino, R.; Vasconcelos, V.; Urbatzka, R. Hierridin B Isolated from a Marine Cyanobacterium Alters VDAC1, Mitochondrial Activity, and Cell Cycle Genes on HT-29 Colon Adenocarcinoma Cells. Mar. Drugs 2016, 14, 158. [Google Scholar] [CrossRef] [Green Version]
- Brandão, P.; Moreira, J.; Almeida, J.; Nazareth, N.; Sampaio-Dias, I.E.; Vasconcelos, V.; Martins, R.; Leão, P.; Pinto, M.; Saraíva, L.; et al. Norhierridin B, a new hierridin B-based hydroquinone with improved antiproliferative activity. Molecules 2020, 25, 1578. [Google Scholar] [CrossRef] [Green Version]
- Tian, Y.; Lin, X.; Zhou, X.; Liu, Y. Phenol derivatives from the sponge-derived fungus Didymellaceae sp. SCSIO F46. Front. Chem. 2018, 6, 536–543. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.X.; Wang, X.F.; Ren, G.W.; Yuan, X.L.; Deng, N.; Ji, G.X.; Li, W.; Zhang, P. Prenylated diphenyl ethers from the marine algal-derived endophytic fungus Aspergillus tennesseensis. Molecules 2018, 23, 2368. [Google Scholar] [CrossRef] [Green Version]
- El-Kashef, D.H.; Youssef, F.S.; Reimche, I.; Teusch, N.; Müller, W.E.G.; Lin, W.; Frank, M.; Liu, Z.; Proksch, P. Polyketides from the marine-derived fungus Aspergillus falconensis: In silico and in vitro cytotoxicity studies. Bioorganic Med. Chem. 2021, 29, 115883. [Google Scholar] [CrossRef]
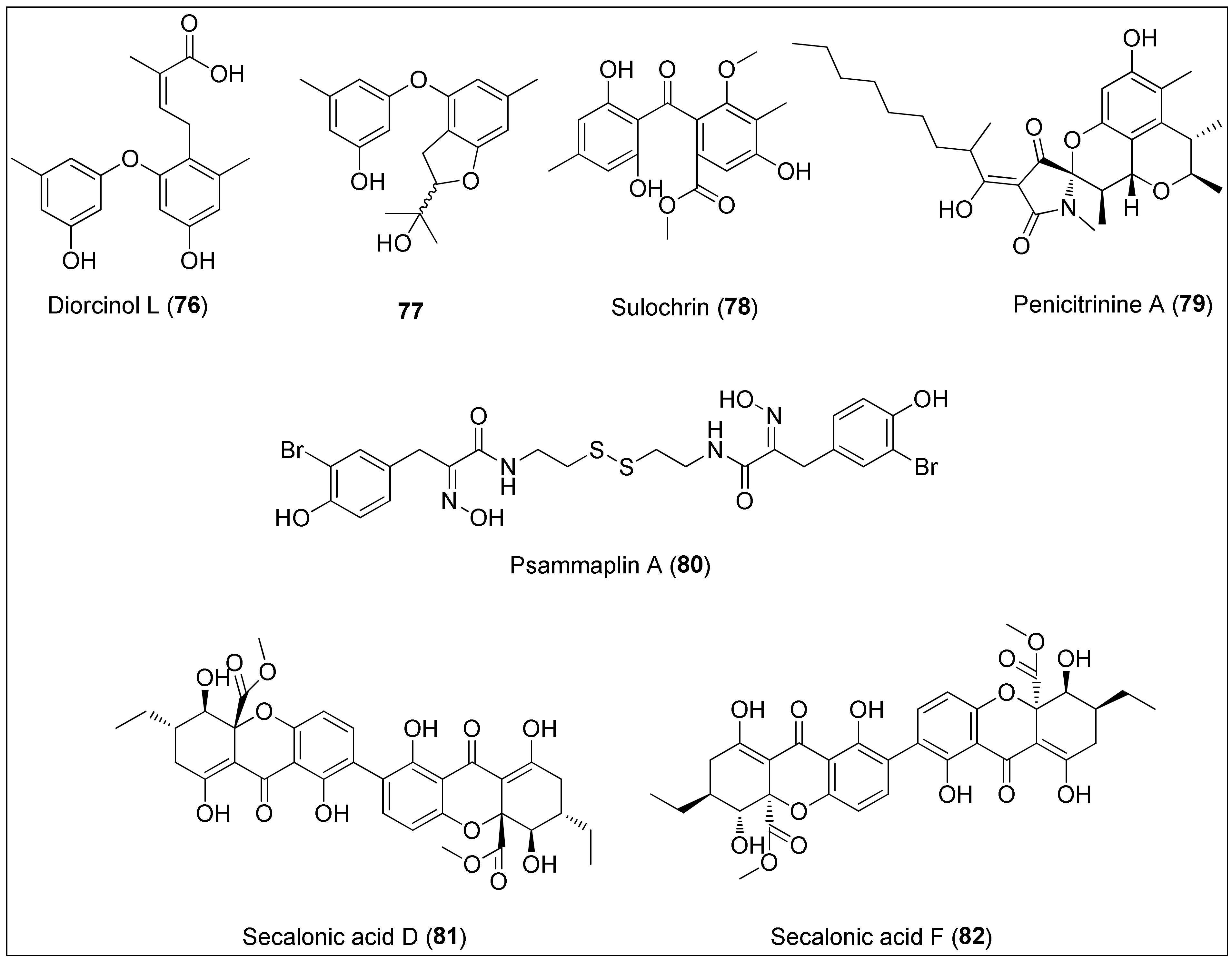
- Liu, Q.Y.; Zhou, T.; Zhao, Y.Y.; Chen, L.; Gong, M.W.; Xia, Q.W.; Ying, M.G.; Zheng, Q.H.; Zhang, Q.Q. Antitumor effects and related mechanisms of Penicitrinine A, a novel alkaloid with a unique spiro skeleton from the marine fungus Penicillium citrinum. Mar. Drugs 2015, 13, 4733–4753. [Google Scholar] [CrossRef]
- Jing, Q.; Hu, X.; Ma, Y.; Mu, J.; Liu, W.; Xu, F.; Li, Z.; Bai, J.; Hua, H.; Li, D. Marine-derived natural lead compound disulfide-linked dimer psammaplin A: Biological activity and structural modification. Mar. Drugs 2019, 17, 384. [Google Scholar] [CrossRef] [Green Version]
- Bao, Y.; Xu, Q.; Wang, L.; Wei, Y.; Hu, B.; Wang, J.; Liu, D.; Zhao, L.; Jing, Y. Studying Histone Deacetylase Inhibition and Apoptosis Induction of Psammaplin A Monomers with Modified Thiol Group. ACS Med. Chem. Lett. 2021, 12, 39–47. [Google Scholar] [CrossRef]
- Ju Han, H.; Sub Byun, W.; Ho Lee, G.; Kyung Kim, W.; Jang, K.; Yang, S.; Yang, J.; Woo Ha, M.; Hong, S.; Lee, J.; et al. Synthesis and biological activity of selenopsammaplin A and its analogues as antitumor agents with DOT1L inhibitory activity. Bioorganic Med. Chem. 2021, 35, 116072. [Google Scholar] [CrossRef]
- Tang, R.; Kimishima, A.; Setiawan, A.; Arai, M. Secalonic acid D as a selective cytotoxic substance on the cancer cells adapted to nutrient starvation. J. Nat. Med. 2020, 74, 495–500. [Google Scholar] [CrossRef]
- Zhang, H.; Huang, L.; Tao, L.; Zhang, J.; Wang, F.; Zhang, X.; Fu, L. Secalonic acid D induces cell apoptosis in both sensitive and ABCG2-overexpressing multidrug resistant cancer cells through upregulating c-Jun expression. Acta Pharm. Sin. B 2019, 9, 516–525. [Google Scholar] [CrossRef]
- Guru, S.K.; Pathania, A.S.; Kumar, S.; Ramesh, D.; Kumar, M.; Rana, S.; Kumar, A.; Malik, F.; Sharma, P.R.; Chandan, B.K.; et al. Secalonic Acid-D represses HIF1α/VEGF-mediated angiogenesis by regulating the Akt/mTOR/p70S6K signaling cascade. Cancer Res. 2015, 75, 2886–2896. [Google Scholar] [CrossRef] [Green Version]
- Sun, H.L.; Zhang, J.; Pan, X.H.; Yan, M.M.; Liu, D.S.; Gao, X.; Zhang, J.R. Secalonic acid- F inhibited cell growth more effectively than 5-fluorouracil on hepatocellular carcinoma in vitro and in vivo. Neoplasma 2017, 64, 344–350. [Google Scholar] [CrossRef]
- Li, N.; Yi, Z.; Wang, Y.; Zhang, Q.; Zhong, T.; Qiu, Y.; Wu, Z.; Tang, X. Differential proteomic analysis of HL60 cells treated with secalonic acid F reveals caspase 3-induced cleavage of Rho GDP dissociation inhibitor 2. Oncol. Rep. 2012, 28, 2016–2022. [Google Scholar] [CrossRef]
- Zeng, Y.; Ren, M.; Li, Y.; Liu, Y.; Chen, C.; Su, J.; Su, B.; Xia, H.; Liu, F.; Jiang, H.; et al. Knockdown of RhoGDI2 represses human gastric cancer cell proliferation, invasion and drug resistance via the Rac1/Pak1/LIMK1 pathway. Cancer Lett. 2020, 492, 136–146. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.; Li, M.; Liu, D.; Wang, X.; Wang, P.; Dai, H.; Yang, W.; Liu, W.; Hu, X.; Zhao, M. Secalonic acid-F, a novel mycotoxin, represses the progression of hepatocellular carcinoma via MARCH1 regulation of the PI3K/AKT/β-catenin signaling pathway. Molecules 2019, 24, 393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Özenver, N.; Dawood, M.; Fleischer, E.; Klinger, A.; Efferth, T. Chemometric and transcriptomic profiling, microtubule disruption and cell death induction by secalonic acid in tumor cells. Molecules 2020, 25, 3224. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Class | Anticancer Properties Displayed by In Vitro, In Vivo or In Silico Experiments | Ref. |
|---|---|---|---|
| Phloroglucinol 1 | Phlorotannin | Inhibition of migration and capillary-tube formation resulting in reduction of Lewis lung carcinoma in mouse model | [79] |
| 50% of MDA-MB-231 cells’ migration at 50 µM by inhibiting epithelial-mesenchymal cell transition via downregulation of SLUG protein | [80] | ||
| Apoptosis of the HT-29 cells through insulin-like growth factor 1 receptor (IGF-1R) and inhibition of downstream proteins | [81,82] | ||
| Upregulation of pro-apoptotic proteins Bax and Bad and downregulation of anti-apoptotic Bcl-2 and Bcl-xL | [81] | ||
| Activity against breast cancer stem-like cells by reducing the expression of CD44, Oct4, Notch2, β-catenin and Sox2 proteins, associated with enhanced sensitivity of cells to the ionizing radiation and standard chemotherapeutic drugs (taxol, cisplatin and etoposide) | [83] | ||
| The higher sensitization of colon cancer cells, HT-29 and HCT116, to 5-fluorouracil (5-FU) | [84] | ||
| Eckol 2 | Exhibition of radioprotective properties in vitro by ROS scavenging, and in vivo through the reduction of side effects caused by gamma ray-irradiation | [85,86] | |
| Decreased expression of the stem-like glioma cells markers, CD133, Nestin and Musashi-1 | [87] | ||
| Increased sensitivity of stem-like glioma cells to cytotoxic prodrug, temozolomide as well as to ionizing radiation via PI3K⁄Akt and Ras⁄Raf-1⁄ERK signalling pathways | |||
| Inhibition of the pancreatic cancer cell line (SW1990) growth | [88] | ||
| Increase of cytotoxic T-lymphocytes required for annihilation of sarcoma (S180) xenograft in mouse model | [90] | ||
| Phlorofucofuroeckol 3 | Decreased HCT116, HT-29, LoVo and SW480 cells’ viabilities for 38%, 31%, 47% and 90%, respectively, at 100 μM | [92] | |
| Induction of apoptosis and enhanced expression of activating transcription factor 3 (ATF3) | |||
| Suppression of migration and invasion of MCF-7 and MDA-MB-231 cells by downregulation of Nf-κB and Toll-like receptor 4 (TLR-4) alongside with reducing MMP-2 and -9 expression | [93] | ||
| Dieckol 4 | Suppression of migration and invasion of MCF-7 and MDA-MB-231 cells by downregulation of Nf-κB and Toll-like receptor 4 (TLR-4) alongside with reducing MMP-2 and -9 expression | [93] | |
| Regulation of human fibrosarcoma cells’ (HT1080) invasion via downregulation of MMP-2 and -9 expression through Nf-κB signalling | [95] | ||
| Inhibition of SK-Hep1 cells’ migration through downregulation of MMP-9 expression | [96] | ||
| Decrease of B16F0 melanoma cells via downregulation of Ras-related C3 botulinum toxin substrate 1 (Rac1) and Wiskott–Aldrich syndrome protein family member 2 (WAVE2) | [98] | ||
| Suppression of HT1080 cells migration via reduced expression and phosphorylation of focal adhesion kinase (FAK) | [99] | ||
| Induction of apoptosis in Hep3B hepatocellular carinoma cell line through cytochrome c release and caspase cascade | [101] | ||
| Binding to MMP-2, MMP-9, MAPK, ERK and p38 via hydrogen bonds revealed by computational studies | [102] | ||
| Suppression of ovarian cancer cells (A2780, SKOV3) growth by inducing caspase-dependent and regulating AKT and p38 signaling | [103] | ||
| Reduced volume of SKOV3 tumor xenograft in mouse model | |||
| Reducing mobility of MCF-7 cells by increasing MMP inhibitors, TIMP-1 and -2, as well as degrading extracellular matrix | [104] | ||
| Increase in Bax/Bcl-2 ratio indicating mitochondrial-mediated apoptosis in SK-BR-3 (breast cancer) and PANC-1 (pancreatic cancer) cells | [105,109] | ||
| Protective and chemopreventing effects against hepatocellular carcinoma (HCC) in rats via upregulation of VEGF, MMP-2/9, PCNA and COX-2 | [106,107] | ||
| Anti-migratory and apoptotic activity in A549 cells associated with inhibition of Pi3K/AKT/mTOR signalling pathways and activation of tumor-suppressor, E-cadherin | [108] | ||
| Protection from gamma radiation and consequent damage, both in vitro and in vivo | [110,111] | ||
| Diphlorethohydroxycarmalol 5 | Activity against murine leukemia (P-388) and vincristine-resistant cells (IC50 = 10.5 and 8.0 µg/mL, respectively) | [112] | |
| Induction of apoptosis in HL60 cells via upregulation of Bax, downregulation of Bcl-2 and increased expression of cleaved caspase-3 and cleaved PARP | [113] | ||
| Photoprotective activity against UVB radiation and side effects, through absorption of radiation, inhibition of MMPs’ expression and scavenging of ROS | [115,116,117,118] | ||
| Dioxinodehydroeckol 6 | Induction of apoptosis in MCF-7 cells via downregulation of p65, IKK (IκB kinase) and NIK (NF-κB inducing kinase) Upregulation of Bax and downregulation of Bcl-2 | [119] | |
| Downregulation of tyrosinase and tyrosinase-linked proteins (TRP-1 and TRP-2) as well as MITF (microphthalmia-associated transcription factor), involved in regulation of melanin formation in B16F10 mealnoma cells | [121] | ||
| Protection against radiation-induced skin damage | [122] | ||
| Fucodiphloroethol G 7 | Activity against HeLa, A549, HT1080 and HT29 cancer cells (IC50 = 298.2, 226.5, 242.5 and 228.5 µM, respectively) | [124] | |
| Suppression of MAPK and Akt signalling pathway necessary for expression of proteinases (MMP-2, MMP-9 and APN) involved in degradation of extracellular matrix | [126] | ||
| Bis (2,3-dibromo-4,5-dihydroxybenzyl) ether (BDDE) 12 | Bromophenol | Actvity against human myelogenous leukemia cells (K562) (IC50 = 13.9 µg/mL) | [137] |
| Induction of mitochondrial pathway-related apoptosis via modulation of Bax, Bcl-2, caspases-3 and -9 in K562 cells | |||
| Topoisomerase I inhibitor by binding in the minor groove of DNA | |||
| Repression of angiogenesis through decrease in the VEGF and VEGFR expression in HUVEC cells associated with inhibition of subintestinal vessel formation in vivo on zebrafish embryos | [139] | ||
| Bis (2,3-dibromo-4,5-dihydroxy-phenyl)-methane (BDDPM) 14 | Anti-proliferative activity against HeLa, RKO, HCT116, Bel7402 and U87 human cancer cell lines (IC50 = 17.63, 11.37, 10.58, 8.7 and 23.69 µg/mL, respectively) | [140] | |
| Mitochondrial-related apoptosis associated with the cleavages of caspases 3 and 9 and PARP | |||
| Inhibition of β1-integrin leading to suppression of Bel7402 cell migration and invasion | |||
| Inhibition of angiogenesis in HUVEC cells through inhibition of several receptor tyrosine kinases and reducing cell invasiveness by lowering the production of NO | [141] | ||
| 18–21 | Antiproliferative activity against DLD-1 (IC50 = 18.3, 14.6, 13.5 and 12.4 µM) and HCT-116 cells (IC50 = 20.4, 14.1, 2.51 and 1.32 µM) | [143] | |
| Lanosol butanone 23 | Antiproliferative activity against HL-60 cells (IC50 = 8.0 µM) | [144] | |
| Bromophenol sulfate 25 | Antiproliferative activity against A2780 cancer cells (IC50 = 9.4 µM) | [145] | |
| Avrainvilleal 29 | Moderate activity against HeLa cancer cells (IC50 = 9.64 ± 1.7 µM) | [147] | |
| Penimethavone A 30 | Flavonoid | Antiproliferative activity against HeLa and RD cancer cells (IC50 = 8.41 and 8.18 µM) | [152] |
| Isorhamnetin-3-O-β-ᴅ-glucoside 31 | Inhibition of MMP-2 and MMP-9 in Ht1080 cells, associated with increase in TIMP-1 protein | [153] | |
| Quercetin-3-O-β-ᴅ-glucoside 32 | |||
| Antiproliferative activity against HCT116 cancer cells (IC50 = 24.3 µM) | [154] | ||
| Myricetin 3-galactoside 33 | Suppression of MMP-2 and MMP-9 expression while simultaneously elevating TIMP-1 and TIMP-2 at the mRNA and protein levels in HT1080 cell line | [155] | |
| Quercetin 3-O-β-ᴅ-galactopyranoside 34 | |||
| Reduced the levels of phosphorylated ERK and p-38 in HT1080 cell lines | |||
| Alternariol 35 | Coumarins | Antiproliferative activity against KB and KBv200 cancer cells (IC50 = 3.17 and 4.82 µM) | [162] |
| Anticancer properties against leukemia cell lines (L1210 and CCRF-CEM) | [163] | ||
| Alternariol methyl ether 36 | Antiproliferative activity against KB and KBv200 cancer cells (IC50 = 3.12 and 4.94 µM) | [162] | |
| Antiproliferative properties against leukemia cell lines (Colon-38 and H-125) | [163] | ||
| Lamellarin D 37 | Antiproliferative activity against LNCaP, DU-145 and K562 cancer cells in the nanomolar range | [164,165] | |
| Antiproliferative activity against CEM and CEM/C2 lines (14 and 969 nM) | [166] | ||
| Topoisomerase I inhibitor due to intercalation at the DNA–topoisomerase I complex | |||
| Induction of apoptosis of P388 and P388CPT5 (topoisomerase I—mutated subclone) cells via increasing the levels of Bax and decreasing the expression of Bcl-2 and cIAP2 along with caspase-3/-9 activation | [167] | ||
| Lamellarin M 38 | Antiproliferative activity against P388, Schabel, AUXB1, CCHRC5, A549, HT29 and MEL28 cancer cells (IC50 = 0.15, 0.17, 0.07, 0.17, 0.06, 0.56 and 0.54 µM) | [172] | |
| Lamellarin K 39 | Antiproliferative activity against P388, Schabel, AUXB1, CCHRC5, A549, HT29 and MEL28 cancer cells (IC50 = 0.19, 0.017, 0.19, 0.75, 0.18, 0.38 and 0.40 µM) | ||
| Aspergiolide A 40 | Antiproliferative activity against A-549, HL-60, BEL-7402 and P388 cancer cells (IC50 = 0.13, 0.28, 7.5 and 35 µM) | [173] | |
| Activity against 11 cancer cells with micromolar IC50 values (2.37–7.07 μM) | [174] | ||
| Induction of caspase-mediated apoptosis of BEL-7402 cancer cells via modulation of Bax and Bcl-2 expression | |||
| Inhibition of DNA topoisomerase II | |||
| Suppressing the increase of H22 and BEL-7402 cancer xenografts in mice without major effects on body weight | |||
| Aspergiolide B 41 | Antiproliferative activity against A-549 and HL-60 cancer cells (IC50 = 0.24 and 0.51 µM) | [175] | |
| Potential EGFR-TK inhibitor displaying low binding free energy in active site containing MET-766, THR-790 and THR-854 amino acid residues | [176] | ||
| Epitaondiol (43), epitaondiol monoacetate (44) and stypodiol (45) | Terpenophenolics | The highest activity of almost 100% inhibition of cell proliferation was observed against human neuroblastoma cells (SH-SY5Y) | [181] |
| Flabellinol 46 | Antiproliferative activity against mouse neuroblastoma cell line (Neuro-2a) and NCI-H460 cells (IC50 = 9 µM) | [183] | |
| Laurebiphenyl 47 | Antiproliferative activity against BGC-823, HeLa, A549, HCT-8 and Bel7402 cancer cells (IC50 = 1.22, 1.61, 1.68, 1.77 and 1.91 µM) | [184] | |
| 7-Hydroxylaurene 48 | Antiproliferative activity against MCF-7, PC3, A431, HeLa and K562, as well as CHO cancer cells (IC50 = 15.8, 18.1, 23.9, 40.5, 64.2 and 78.2 µM) | [186] | |
| (+)-curcuphenol 51 | Antiproliferative activity against P388 cancer cells (IC50 = 7 µg/mL) | [178] | |
| Antiproliferative activity against p53+/+, p53−/−, p21+/+ and p21−/− cancer cells (IC50 = 27, 33, 33 and 35 µg/mL) | [189] | ||
| Inhibition of CaCo-2 colon proliferation and DNA synthesis associated with induction of apoptosis via caspase-3 activation | [188] | ||
| Sargaquinoic acid (52), sargahydroquinoic acid (53) | Antiproliferative activity against P388 cancer cells (IC50 = 17 and 14 µM) | [190] | |
| Plastoquinones 54 and 55 | Antiproliferative activity against murine colon 26-L5 adenocarcinoma cell line (IC50 = 1.51 and 1.69 μg/mL) | [192] | |
| Moromycin B (56), saquayamycins B (57) and B1 (58) | Quinones and hydroquinones | Inhibition of proliferation of breast cancer cells (MCF-7, MDA-MB-231 and BT-474) in sub-micromolar range (0.16–0.67 μM) | [195] |
| Saquayamycin B (57) | Suppression of MDA-MB-231 cells invasion and migration at concentrations of 25 and 50 nM for 12 h | ||
| Buanmycin 59 | Antiproliferative activity against A549, HCT116, SNU638, SK-HEP1 and MDA-MB-231 cancer cells (IC50 = 1.7, 0.9, 0.8, 1.9 and 1.2 μM) | [196] | |
| Komodoquinone A 60 | Morphological changes and neuritogenic activity against neuroblastoma cell line (Neuro 2A) | [197,198] | |
| Induction of Neuro 2A cell cycle arrest at G1 phase | |||
| Fradimycin B 62 | Antiproliferative activity against HCT-15, SW620 and C6 cancer cells (IC50 = 0.13, 4.33 and 0.47 μM) | [199] | |
| Induction of cell cycle arrest at G0/G1 phase associated with an increase of apoptotic and necrotic cells | |||
| Makaluvamine J 66 | Antiproliferative activity against PANC-1 (IC50 = 0.054 μM) and OVCAR-5 cancer cells (IC50 = 120 nM) | [203] | |
| Makaluvamine F 68 | Antiproliferative activity against HCT-116 (IC50 = 0.17 μM) | [205] | |
| Role in hypoxia conditions via inhibition of HIF-1α and its downstream target, VEGF | [206] | ||
| Mayamycin 69 | Antiproliferative activity against eight cancer cell lines: HepG2, HT-29, GXF251L, LXF529L, MAXF401NL, MEXF462NL, PAXF1657L and RXF486L (IC50 = 0.2, 0.3, 0.2, 0.16, 0.29, 0.13, 0.15 and 0.33 μM) | [210] | |
| N-acetyl-N-demethylmayamycin 70 | Antiproliferative activity against U251, U87-MG, SHG-44 and C6 glioma cell lines (IC50 = 0.7, 1.4, 3.9 and 0.5 μM) | [211] | |
| Induction of apoptosis in U251 cells | |||
| Monacyclinone F 71 | Antiproliferative activity against rhabdomycosarcoma cancer cells (SJCRH30) (EC50 = 0.73 μM) | [214] | |
| Galvaquinone B 72 | Antiproliferative activity against Calu-3 and H2887 cancer cells (SJCRH30) (IC50 = 5.0 and 12.2 μM) | [216] | |
| Lupinacidin A 73 | Antiproliferative activity against Calu-3 and H2887 cancer cells (SJCRH30) (IC50 = 8.8 and 3.1 μM) | ||
| Hierrdin B 74 | Antiproliferative activity against HT-29 cancer cells (IC50 = 100 μM) | [218] | |
| Targeting mitochondrial activity by increasing mRNA expression of VDAC1 accompanied with inhibition of cell cycle progression | [219] | ||
| Norhierridin B 75 | Antiproliferative activity against MDA-MB-231, SKBR3, MDA-MB-468, A375, Huh-7 and HCT116 cancer cells (IC50 = 0.61, 0.77, 0.68, 2.0, 0.61 and 3.2 μM) | [220] | |
| Diorcinol L 76 | Miscellaneous | Antiproliferative activity against Huh-7, DU145, HeLa and HL60 cancer cells (IC50 = 5.7, 9.1, 7.1 and 9.6 µM) | [221] |
| Sulochrin 78 | Antiproliferative activity against L5178Y cancer cells (IC50 = 5.1 µM) | [223] | |
| Inhibition of MDA-MB-231 cell migration | |||
| Inhibition of CDK-2, TOP-2 and MMP-13 proteins | |||
| Penicitrinine A 79 | Antiproliferative activity against A-375 cancer cells (IC50 = 30.88, 12.78 and 7.06 µM, after treatment for 24 h, 48 h and 72 h, respectively) | [224] | |
| Induction of apoptosis of A-375 cells by modulation of Bcl-2 and Bax expression | |||
| Suppression of A-375 cell migration by downregulating MMP-9 and upregulating TIMP-1 levels | |||
| Psammaplin A 80 | Inhibition of aminopeptidase N (APN), mycothiol-S-conjugate amidase (MCA), topoisomerase II, farnesyl protein transferase, histone deacetylase (HDAC) and leucine aminopeptidase, involved in angiogenesis, DNA replication, apoptosis, proliferation and invasion | [225] | |
| Secalonic acid D 81 | Antiproliferative activity against PANC-1, S1, S1-MI-80, H460, H460/MX20, MCF-7 and MCF-7/ADR cancer cells (IC50 = 0.6, 6.8, 6.4, 5.3, 4.9, 5.1 and 4.9 µM) | [228,229] | |
| Induction of cell death through c-Jun/Src/STAT3 signalling by upregulation of c-Jun expression | [229] | ||
| Inhibition of eNOS and ERK phosphorylation together with MMP degradation via Akt/mTOR/p70S6K pathway | [230] | ||
| Secalonic acid F 82 | Antiproliferative activity against HL-60 cancer cells (IC50 = 4.1 µg/mL) | [232] | |
| Induction of apoptosis via caspase-3 activation and modulation of RhoGDI2 protein | |||
| Antiproliferative activity against HepG2 cancer cells (IC50 = 45.4, 8.7 and 7.7 µM, after treatment for 24 h, 48 h and 72 h, respectively) | |||
| Cell cycle arrest at the G1 phase and apoptosis induction activating caspases-3 and -9 | |||
| Suppression of HepG2 and Hep3B cells invasion via downregulation of MARCH1 | [234] | ||
| Toxicity towards leukemia and multiple myeloma cells mediated through apoptosis and necrosis as well as tubulin disassembly | [235] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matulja, D.; Vranješević, F.; Kolympadi Markovic, M.; Pavelić, S.K.; Marković, D. Anticancer Activities of Marine-Derived Phenolic Compounds and Their Derivatives. Molecules 2022, 27, 1449. https://doi.org/10.3390/molecules27041449
Matulja D, Vranješević F, Kolympadi Markovic M, Pavelić SK, Marković D. Anticancer Activities of Marine-Derived Phenolic Compounds and Their Derivatives. Molecules. 2022; 27(4):1449. https://doi.org/10.3390/molecules27041449
Chicago/Turabian StyleMatulja, Dario, Filip Vranješević, Maria Kolympadi Markovic, Sandra Kraljević Pavelić, and Dean Marković. 2022. "Anticancer Activities of Marine-Derived Phenolic Compounds and Their Derivatives" Molecules 27, no. 4: 1449. https://doi.org/10.3390/molecules27041449
APA StyleMatulja, D., Vranješević, F., Kolympadi Markovic, M., Pavelić, S. K., & Marković, D. (2022). Anticancer Activities of Marine-Derived Phenolic Compounds and Their Derivatives. Molecules, 27(4), 1449. https://doi.org/10.3390/molecules27041449