
Novel Chemical and Biological Insights of Inositol Derivatives in Mediterranean Plants



Abstract
:1. Introduction
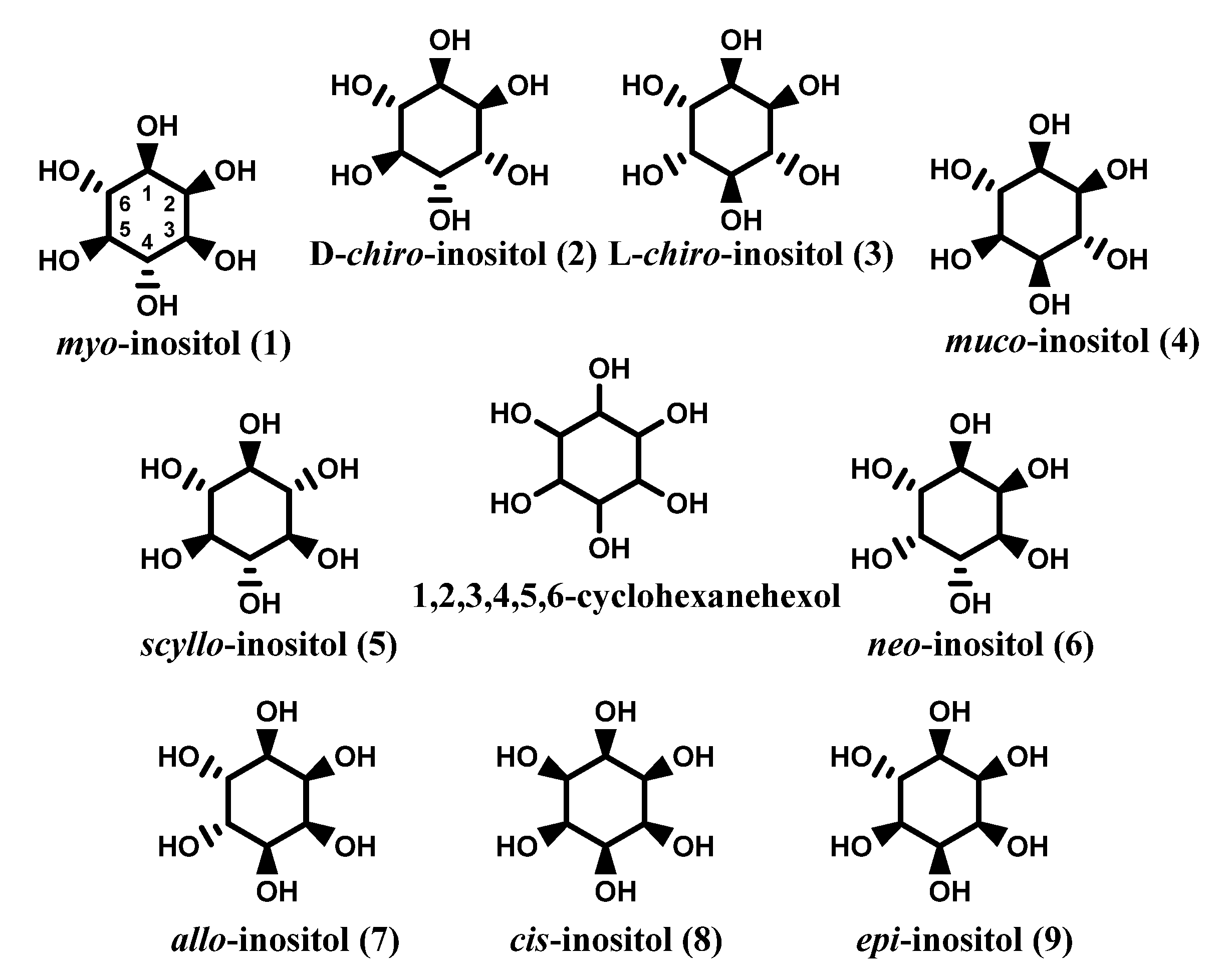
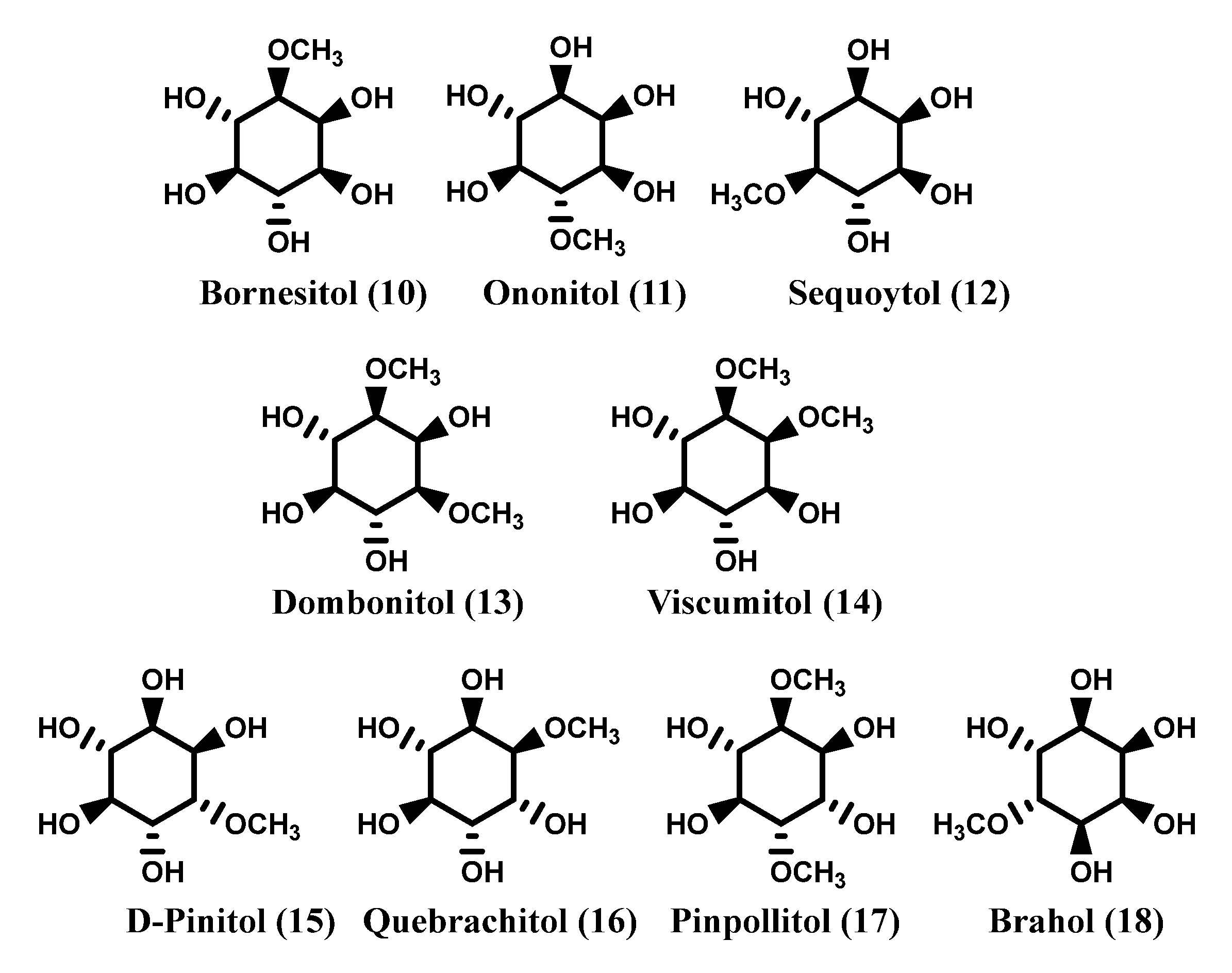
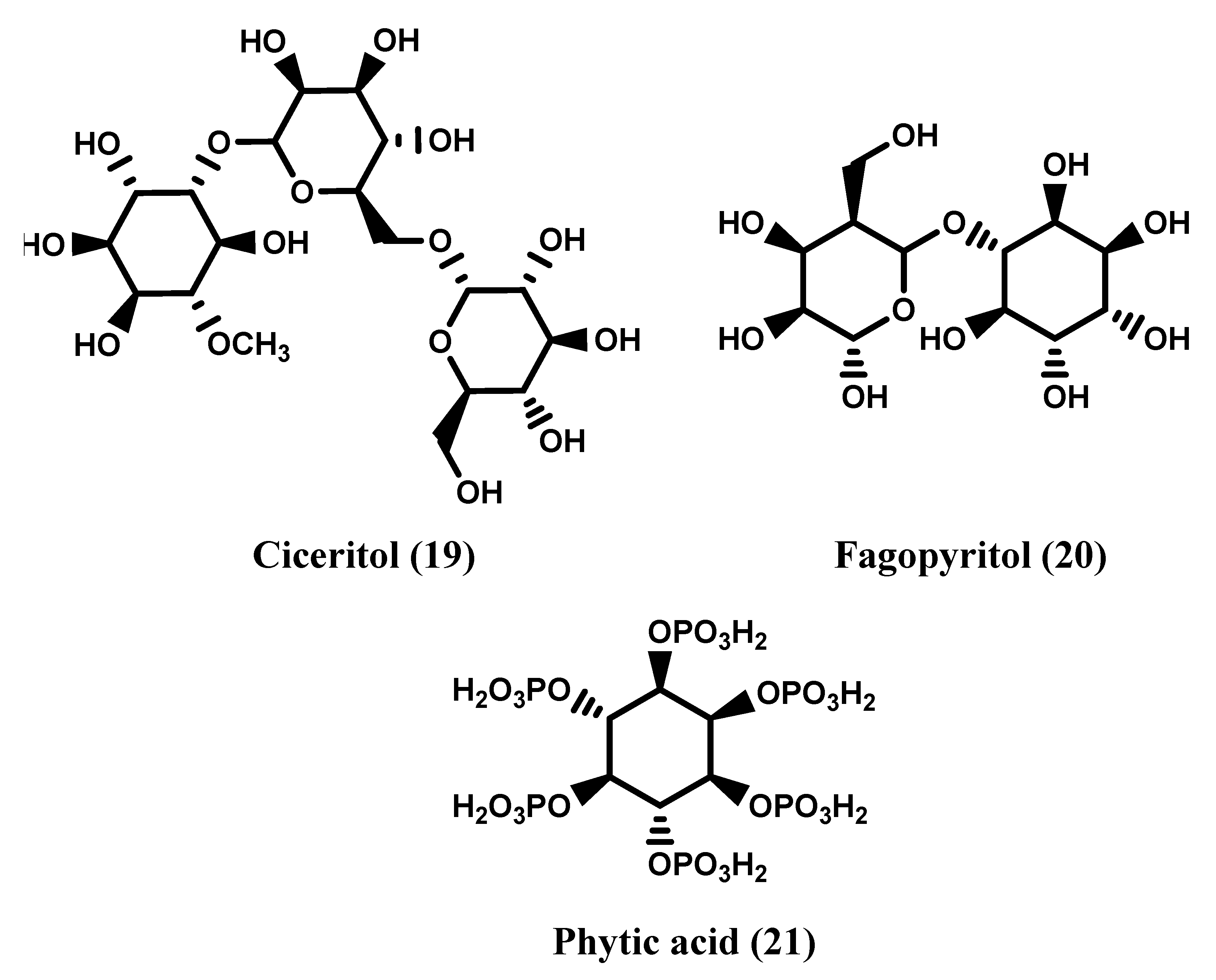
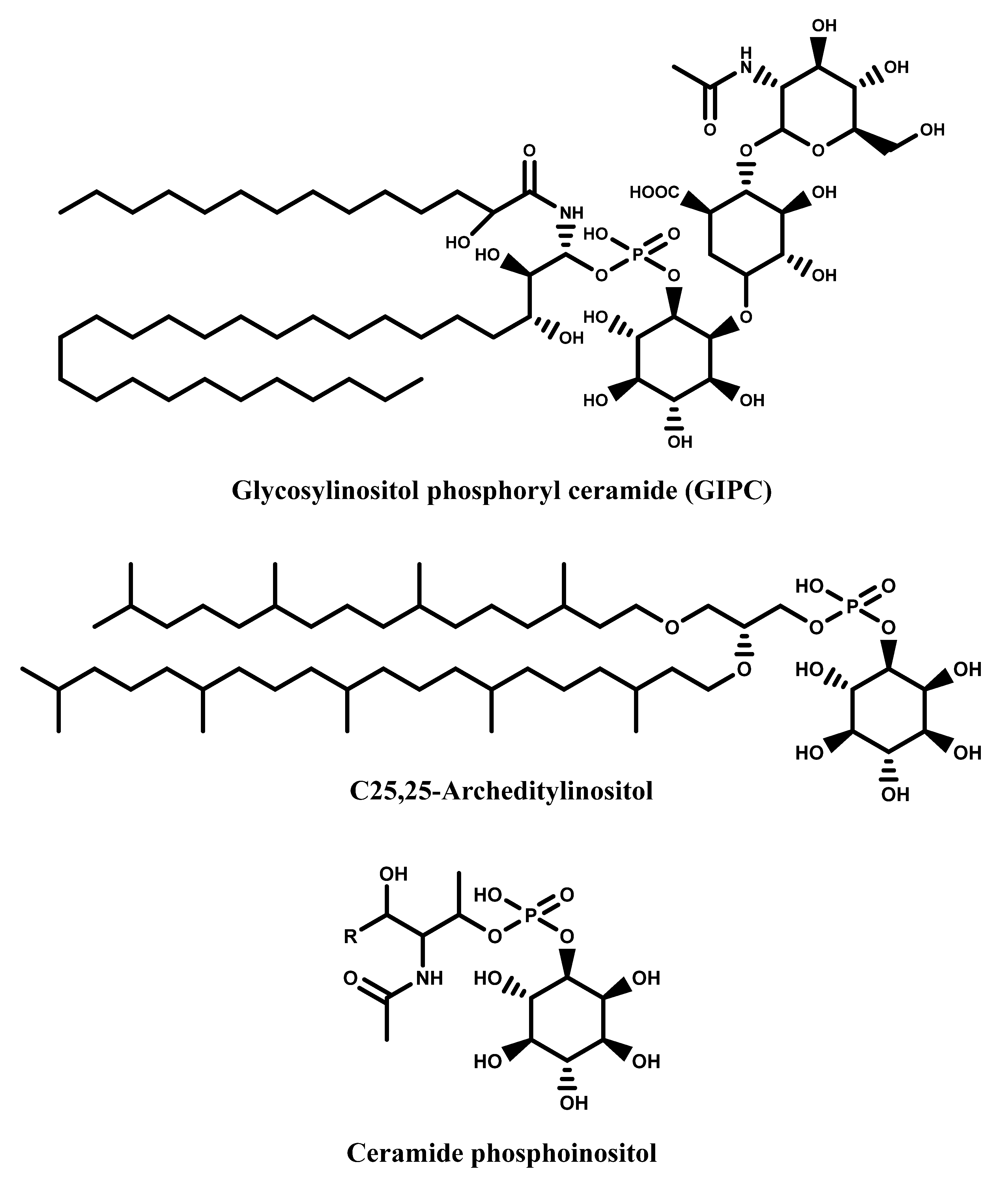
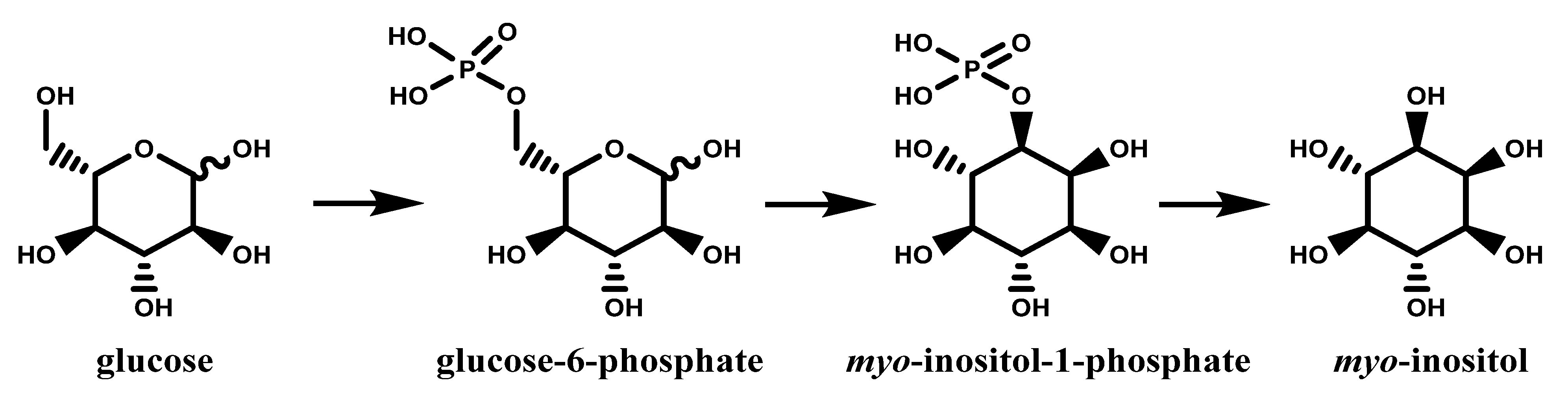
1.1. Inositols
1.2. The Dual Nature of Primary and Secondary Metabolites of Inositols
2. Natural Occurrence of Inositols
2.1. Natural Sources of Inositols: Some Examples
2.2. Accumulation of Inositols in Response to Biotic and Abiotic Stresses
3. Biological Activities of Inositols
3.1. Inositols and Polycystic Ovary Syndrome (PCOS)
3.2. Inositols in Metabolic Syndrome and Diabetes
3.3. Inositols in Neurodegenerative and Neurologic Disorders
3.4. Inositols and Sars-Cov-2
3.5. Inositols and Cancer
4. Concluding Remarks
Author Contributions
Funding
Informed Consent Statement
Conflicts of Interest
Sample Availability
References
- Michell, R.H. Inositol and its derivatives: Their evolution and functions. Adv. Enzym. Regul. 2011, 51, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Duchek, J.; Adams, D.R.; Hudlicky, T. Chemoenzymatic synthesis of inositols, conduritols, and cyclitol analogues. Chem. Rev. 2011, 111, 4223–4258. [Google Scholar] [CrossRef] [PubMed]
- Thomas, M.P.; Mills, S.J.; Potter, B.V.L. The “other” inositols and their phosphates: Synthesis, biology, and medicine (with recent advances in myo-inositol chemistry). Angew. Chem. 2016, 55, 1614–1650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ehlers, A.; Marakis, G.; Lamperi, A.; Hirsch-Ernst, K.I. Risk assessment of energy drinks with focus on cardiovascular parameters and energy drink consumption in Europe. Food Chem. Toxicol. 2019, 130, 109–121. [Google Scholar] [CrossRef] [PubMed]
- Pani, A.; Giossi, R.; Menichelli, D.; Fittipaldo, V.A.; Agnelli, F.; Inglese, E.; Roamndini, R.; Roncato, R.; Pintaudi, B.; Del Sole, F.; et al. Inositol and non-alcoholic fatty liver disease: A systematic review on deficiencies and supplementation. Nutrients 2020, 12, 3379. [Google Scholar] [CrossRef]
- Ruiz-Aceituno, L.; Rodriguez-Sanches, S.; Sanza, J.; Sanz, M.L.; Ramos, L. Optimization of pressurized liquid extraction of inositols from pine nuts (Pinus pinea L.). Food Chem. 2014, 153, 450–456. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Han, P.; Wang, J.; Shi, T.; You, C. Production of myo-inositol: Recent advances and prospective. Biotechnol. Appl. Biochem. 2021. [Google Scholar] [CrossRef]
- Jia, Q.; Kong, D.; Li, Q.; Sun, S.; Song, J.; Zhu, Y.; Liang, K.; Ke, Q.; Lin, W.; Huang, J. The function of inositol phosphatase in plant tolerance to abiotic stress. Int. J. Mol. Sci. 2019, 20, 3999. [Google Scholar] [CrossRef] [Green Version]
- Dinicola, S.; Minini, M.; Unfer, V.; Verna, R.; Cucina, A.; Bizzarri, M. Nutritional and acquired deficiencies in inositol bioavailability. Correlations with metabolic disorders. Int. J. Mol. Sci. 2017, 18, 2187. [Google Scholar] [CrossRef]
- Owczarczyk-Saczonek, A.; Lahuta, L.B.; Ligor, M.; Placek, W.; Gorecki, R.J.; Buszewski, B. The healing-promoting properties of selected cyclitols—A review. Nutrients 2018, 10, 1891. [Google Scholar] [CrossRef] [Green Version]
- Ford, C.W. Identification of inositols and their mono-O-methyl ethers by gas-liquid chromatography. J. Chromatogr. 1985, 333, 167–170. [Google Scholar] [CrossRef]
- Richter, A. Viscumitol, a dimethyl-ether of muco-inositol from Viscum album. Phytochemistry 1992, 31, 3925–3927. [Google Scholar] [CrossRef]
- Sanchez-Hidalgo, M.; Leon-Gonzalez, A.J.; Galvez-Peralia, M.; Gonzalez-Mauraz, N.H.; Martin-Corder, C. d-Pinitol: A cyclitol with versatile biological and pharmacological activities. Phytochem. Rev. 2021, 20, 211–224. [Google Scholar] [CrossRef]
- Ahmad, V.U.; Ali, Z.; Ali, M.S.; Zahid, M.; Tareen, R.B. Brahol: A new derivative of allo-inositol from Stocksia barhuica. Nat. Prod. Sci. 1998, 4, 170–173. [Google Scholar]
- Quemener, B.; Brillouet, J.-M. Ciceritol, a pinitol digalactoside from seeds of chickpea, lentil and white lupin. Phytochemistry 1983, 22, 1745–1751. [Google Scholar] [CrossRef]
- Horbowicz, M.; Brenac, P.; Obendorf, R.L. Fagopyritol B1, O-α-d-galactopyranosil-(1→2)-d-chiro-inositol, a galactosyl cyclitol in maturing buckwheat seeds associated with desiccation tolerance. Planta 1998, 205, 1–11. [Google Scholar] [CrossRef]
- Steadman, K.J.; Burgoon, M.S.; Shuster, R.L.; Lewis, B.A.; Edwardson, S.E.; Obendorf, R.L. Fagopyritols, d-chiro-inositol, and other soluble carbohydrates in buckwheat seed milling fractions. J. Agric. Food Chem. 2000, 48, 2843–2847. [Google Scholar] [CrossRef]
- Wu, W.; Wang, L.; Qiu, J.; Li, Z. The analysis of fagopyritols from tartary buckwheat and their anti-diabetic effects in KK-Ay type 2 diabetic mice and HepG2 cells. J. Funct. Foods 2018, 50, 137–146. [Google Scholar] [CrossRef]
- Wilson, M.S.C.; Livermore, T.M.; Saiardi, A. Inositol pyrophosphate: Between signaling and metabolism. Biochem. J. 2013, 452, 369–379. [Google Scholar] [CrossRef] [Green Version]
- Brown, N.W.; Marmelstein, A.M.; Fiedler, D. Chemical tools, for interrofating pyrophosphate structure and function. Chem. Soc. Rev. 2016, 45, 6311–6326. [Google Scholar] [CrossRef]
- Marolt, G.; Kolar, M. Analytical methods for determination of phytic acid and other inositol phosphate: A review. Molecules 2021, 26, 174. [Google Scholar] [CrossRef]
- Shears, S.B.; Wang, H. Metabolism and functions of inositol pyrophosphates: Insights gained from the application of synthetic analogues. Molecules 2020, 25, 4515. [Google Scholar] [CrossRef]
- Lorenzo-Orts, L.; Couto, D.; Hothorn, M. Identity and functions of inorganic and isositol polyphosphates in plants. New Phytol. 2020, 225, 637–652. [Google Scholar] [CrossRef] [Green Version]
- Duong, Q.H.; Clark, K.D.; Lapsley, K.G.; Pegg, R.B. Quantification of inositol phosphates in almond meal and almond brown skins by HPLC/ESI/MS. Food Chem. 2017, 229, 84–92. [Google Scholar] [CrossRef]
- Feizollahi, E.; Mirmahdi, R.S.; Zoghi, A.; Zijlstra, R.T.; Roopesh, M.S.; Vasanthan, T. Review of the beneficial and anti-nutritional qualities of phytic acid, and procedures for removing it from food products. Food Res. Int. 2021, 143, 110284. [Google Scholar] [CrossRef]
- Silva, V.M.; Putti, F.F.; White, P.J.; dos Reis, A.R. Phytic acid accumulation in plants: Biosynthesis pathway regulation and role in human diet. Plant Physiol. Biochem. 2021, 164, 132–146. [Google Scholar] [CrossRef]
- Maga, J.A. Phytate: Its chemistry, occurrence, food interactions, nutritional significance, and methods of analysis. J. Agric. Food Chem. 1982, 30, 1–9. [Google Scholar] [CrossRef]
- Blaabjerg, K.; Hansen-Møller, J.; Poulsen, H.D. High-performance ion chromatography method for separation and quantification of inositol phosphates in diets and digests. J. Chromatogr. B 2010, 878, 347–354. [Google Scholar] [CrossRef] [PubMed]
- Tartaglio, V.; Rennie, E.A.; Cahoon, R.; Wang, G.; Baidoo, E.; Mortimer, J.C.; Cahoon, E.B.; Scheller, H.V. Glycosilation of inositol phosphorylceramide sphingolipids is required for normal growth and reproduction in Arabidopsis. Plant J. 2017, 89, 278–290. [Google Scholar] [CrossRef]
- Buré, C.; Cacas, J.-L.; Mongrand, S.; Schmitter, J.-M. Characterization of glycosyl inositol phosphoryl ceramides from plants and fungi by mass spectrometry. Anal. Bioanal. Chem. 2014, 406, 995–1010. [Google Scholar] [CrossRef]
- Gronnier, J.; Germain, V.; Gouguet, P.; Cacas, J.-L.; Mongrand, S. GIPC: Glycosyl inositol phospho ceramides, the major sphingolipids on earth. Plant Signal. Behav. 2016, 11, e1152438. [Google Scholar] [CrossRef] [Green Version]
- Hasi, R.Y.; Majima, D.; Morito, K.; Ali, H.; Kogure, K.; Nanjundan, M.; Hayashi, J.; Kawakami, R.; Kanemaru, K.; Tanaka, T.J. Isolation of glycosylinositol phosphoceramide and phytoceramide 1-phosphate in plants and their chemical stabilities. J. Chromatogr. B 2020, 1152, 122213. [Google Scholar] [CrossRef] [PubMed]
- Cassim, A.M.; Navon, Y.; Gao, Y.; Decossas, M.; Fouillen, L.; Grélard, A.; Nagano, M.; Lambert, O.; Behammou, D.; Ven Delft, P.; et al. Biophysical analysis of the plant-specific GIPC sphingolipids reveals multiple modes of membrane regulation. J. Biol. Chem. 2021, 296, 100602. [Google Scholar] [CrossRef] [PubMed]
- Gharwalowa, L.; Kulisova, M.; Vasyliuk, A.; Palyzova, A.; Nedbalova, L.; Kolouchova, I. Sphingolipids of plant pathogenic fungi. Plant Prot. Sci. 2021, 57, 134–139. [Google Scholar] [CrossRef]
- Moghimipour, E.; Kargar, M.; Ramezani, Z.; Handali, S. The potent in vitro skin of archaeosome made lipids extracted of Sulfolobus acidocaldarius. Archaea 2013, 2013, 782012. [Google Scholar]
- Genova, J.; Ulrih, N.P.; Kralj-Iglič, V.A.; Bivas, I. Bending elasticity modulus of giant vesicles composed of Aeropyrum pernix K1 archaeal lipid. Life 2012, 5, 1101–1110. [Google Scholar] [CrossRef] [Green Version]
- Drew, E.A. Physiology and metabolism of cyclitols. In Storage Carbohydrates in Vascular Plants: Distribution, Physiology and Metabolism; Cambridge University Press: Cambridge, UK, 1984; Chapter 6; pp. 133–156. ISBN 0-521-23698-3. [Google Scholar]
- Loewus, F.A.; Pushpalata, P.N. Murthy, myo-Inositol metabolism in plants. Plant Sci. 2000, 150, 1–19. [Google Scholar] [CrossRef]
- Campbell, J.A.; Goheen, S.C.; Donald, P. Recent trends for enhancing the diversity and quality of soybean products. In Extraction and Analysis of Inositols and Other Carbohydrates from Soybean Plant Tissues; Krezhova, D., Ed.; IntechOpen: London, UK, 2011; pp. 281–304. [Google Scholar]
- Al-Suod, H.; Ligor, M.; Ratiu, I.-A.; Rafińska, K.; Gòrecki, R.; Buszewski, B. A window on cyclitols: Characterization and analytics of inositols. Phytochem. Lett. 2017, 20, 507–519. [Google Scholar] [CrossRef]
- Ratiu, I.-A.; Al-Suod, H.; Ligor, M.; Ligor, T.; Krakowska, A.; Górecki, R.; Buszewski, B. Simultaneous determination of cyclitols and sugars following a comprehensive investigation of 40 plants. Food Anal. Methods 2019, 12, 1466–1478. [Google Scholar] [CrossRef] [Green Version]
- Hernández-Hernández, O.; Ruiz-Aceituno, L.; Sanz, M.L.; Martínez-Castro, I. Determination of free inositols and other low molecular weight carbohydrates in vegetables. J. Agric. Food Chem. 2011, 59, 2451–2455. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-Aceituno, L.; Rodríguez-Sánchez, S.; Ruiz-Matute, A.I.; Ramos, L.; Soria, A.C.; Sanz, M.L. Optimisation of a biotechnological procedure for selective fractionation of bioactive inositols in edible legume extracts. J. Sci. Food Agric. 2013, 93, 2797–2803. [Google Scholar] [CrossRef]
- Zuluaga, A.M.; Mena-García, A.; Monzón, A.C.S.; Rada-Mendoza, M.; Chito, D.M.; Ruiz-Matute, A.I.; Sanz, M.L. Microwave assisted extraction of inositols for the valorization of legume by-products. LWT 2020, 133, 109971. [Google Scholar] [CrossRef]
- Giami, S.Y.; Akusu, M.O.; Emelike, J.N. Evaluation of selected food attributes of four advanced lines of ungerminated and germinated Nigerian cowpea (Vigna unguiculata (L.) Walp. Plant Foods Hum. Nutr. 2001, 56, 61–73. [Google Scholar] [CrossRef]
- Da Silva-Ribeiro, E.; da Cruz-Centeno, D.; de Cássia Figueiredo-Ribeiro, R.; Valevski Sales Fernandes, K.; Xavier-Filho, J.; Oliveira, A.E.A. Free cyclitol, soluble carbohydrate and protein contents in Vigna unguiculata and Phaseolus vulgaris bean sprouts. J. Agric. Food Chem. 2011, 59, 4273–4278. [Google Scholar] [CrossRef]
- Al-Suod, H.; Ratiu, I.-A.; Ligor, M.; Ligor, T.; Buszewski, B. Determination of sugars and cyclitols isolated from various morphological parts of Medicago sativa L. J. Sep. Sci. 2018, 41, 1118–1128. [Google Scholar] [CrossRef]
- Al-Suod, H.; Ratiu, I.-A.; Krakowska-Sieprawska, A.; Lahuta, L.; Górecki, R.; Buszewski, B. Supercritical fluid extraction in isolation of cyclitols and sugars from chamomile flowers. J. Sep. Sci. 2019, 42, 3243–3252. [Google Scholar] [CrossRef]
- Sarvin, B.A.; Seregin, A.P.; Shpigun, O.A.; Rodin, I.A.; Stavrianidi, A.N. A novel strategy for isolation and determination of sugars and sugar alcohols from conifers. J. Chromatogr. B 2018, 1092, 138–144. [Google Scholar] [CrossRef]
- Moreira, L.N.; Silva, G.C.; Câmara, D.V.; Pádua, R.M.; Lemos, V.S.; Braga, F.C.; Cortes, S.F. The cyclitol L-(-)-bornesitol as an active marker for the cardiovascular activity of the Brazilian medicinal lant Hancornia speciosa. Biol. Pharm. Bull. 2019, 42, 2076–2082. [Google Scholar] [CrossRef] [Green Version]
- Al-Suod, H.; Gadzała-Kopciuch, R.; Buszewski, B. Simultaneous HPLC-ELSD determination of sugars and cyclitols in different parts of Phacelia tanacetifolia Benth. Biochem. Syst. Ecol. 2018, 80, 32–38. [Google Scholar] [CrossRef]
- Siracusa, L.; Occhiuto, C.; Molonia, M.S.; Cimino, F.; Palumbo, M.; Saija, A.; Speciale, A.; Rocco, C.; Ruberto, G.; Cristani, M. A pinitol-rich Glycyrrhiza glabra L. leaf extract as functional supplement with potential in the prevention of endothelial dysfunction through improving insulin signaling. Arch. Phys. Biochem. 2022. [Google Scholar] [CrossRef]
- Molonia, M.S.; Occhiuto, C.; Muscarà, C.; Specale, A.; Ruberto, G.; Siracusa, L.; Cristani, M.; Sajia, A.; Cimino, F. Effects of a pinitol-rich Glycyrrhiza glabra L. leaf extract on insulin and inflammatory signaling pathways in palmitate-induced hypertrophic adipocytes. Nat. Prod. Res. 2022. [Google Scholar] [CrossRef] [PubMed]
- Streeter, J.G.; Lohnes, D.J.; Fioritto, R.J. Patterns of pinitol accumulation in soybean plants and relationships to drought tolerance. Plant Cell Environ. 2001, 24, 429–438. [Google Scholar] [CrossRef]
- Foti, C.; Kalampokis, I.F.; Aliferis, K.A.; Pavli, O.I. Metabolic Responses of two contrasting lentil genotypes to PEG-induced drought stress. Agronomy 2021, 11, 1190. [Google Scholar] [CrossRef]
- Liu, X.; Grieve, C. Accumulation of chiro-inositol and other non-structural carbohydrates in Limonium species in response to saline irrigation waters. J. Am. Soc. Hortic. Sci. 2009, 134, 329–336. [Google Scholar] [CrossRef] [Green Version]
- Sanz, M.L.; Sanz, J.; Martínez-Castro, I. Presence of some cyclitols in honey. Food Chem. 2004, 84, 133–135. [Google Scholar] [CrossRef]
- Ratiu, I.-A.; Al-Suod, H.; Bukowska, M.; Ligor, M.; Buszewski, B. Correlation study of honey regarding their physicochemical properties and sugars and cyclitols content. Molecules 2020, 25, 34. [Google Scholar] [CrossRef] [Green Version]
- Siracusa, L.; Ruberto, G. Plant polyphenol profiles as a tool for traceability and valuable support to biodiversity. In Polyphenols in Plants: Isolation, Purification and Extract Preparation; Elsevier: Amsterdam, The Netherlands, 2014; Chapter 2; pp. 15–33. ISBN 9780123979346. [Google Scholar]
- Dinicola, S.; Unfer, V.; Facchinetti, F.; Soulage, C.O.; Greene, N.D.; Bizzarri, M.; Laganà, A.S.; Chan, S.; Bevilacqua, A.; Pkhaladze, L.; et al. Inositols: From established knowledge to novel approach. Int. J. Mol. Sci. 2021, 22, 10575. [Google Scholar] [CrossRef]
- Gambioli, R.; Forte, G.; Aragona, C.; Bevilacqua, A.; Bizzarri, M.; Unfer, V. The use of d-chiro-inositol in clinical practice. Eur. Rev. Med. Pharm. Sci. 2021, 25, 438–446. [Google Scholar]
- Caputo, M.; Bona, E.; Leone, I.; Samà, M.T.; Nuzzo, A.; Ferrero, A.; Aimaretti, G.; Marzullo, P.; Prodam, F. Inositols and metabolic disorders: From farm to bedside. J. Tradit. Comp. Med. 2020, 10, 252–259. [Google Scholar] [CrossRef]
- Gambioli, R.; Oliva, M.M.; Nordio, M.; Chiefari, A.; Puliani, G.; Unfer, V. New insights into the activities of d-chiro-inositol: A narrative review. Biomedicine 2021, 9, 1378. [Google Scholar] [CrossRef]
- Lepore, E.; Lauretta, R.; Bianchini, M.; Mormando, M.; Di Lorenzo, C.; Unfer, V. Inositols depletion and resistance: Principal mechanisms and therapeutic strategies. Int. J. Mol. Sci. 2021, 22, 6796. [Google Scholar] [CrossRef]
- Franks, S. Polycystic ovary syndrome. Medicine 2017, 9, 527–531. [Google Scholar] [CrossRef]
- Laganà, A.S.; Garzon, S.; Casarin, J.; Franchi, M.; Ghezzi, F. Inositol in polycystic ovary syndrome: Restoring fertility through a pathophysiology based approach. Trends Endocrinol. Metab. 2018, 28, 768–780. [Google Scholar] [CrossRef]
- Facchinetti, F.; Unfer, V.; Dewailly, D.; Kamenov, Z.A.; Diamanti-Kandakis, E.; Laganà, A.S.; Nestler, J.E.; Soulage, C.O. Inositols in polycystic ovary syndrome: An overview on the advances. Trends Endocrinol. Metab. 2020, 31, 435–447. [Google Scholar] [CrossRef]
- Kamenov, Z.; Gateva, A. Inositols in PCOS. Molecules 2020, 25, 5566. [Google Scholar] [CrossRef]
- Monastra, G.; Vucenik, I.; Harrath, A.H.; Alwasel, S.H.; Kamenov, Z.A.; Laganà, A.S.; Monti, N.; Fedeli, V.; Bizzarri, M. PCOS and inositols: Controversial results and necessary clarifications. Basic differences between d-chiro and myo-inositol. Front. Endocrinol. 2021, 12, 660381. [Google Scholar] [CrossRef]
- Genazzani, A. Inositols. Reflection on how to choose the appropriate one for PCOS. Gynecol. Endocrinol. 2020, 36, 1045–1046. [Google Scholar] [CrossRef]
- Tabrizi, R.; Ostadmohammadi, V.; Lankarani, K.B.; Peymani, P.; Akbari, M.; Kolahdooz, F.; Asemi, Z. The effects of inositol supplementation on lipid profiles among patients with metabolic diseases: A systematic review and meta-analysis of randomized controlled trials. Lipids Health Dis. 2018, 17, 123. [Google Scholar] [CrossRef] [Green Version]
- Montt-Guevara, M.M.; Finiguerra, M.; Marzi, I.; Fidecicchi, T.; Ferrari, A.; Genazzani, A.D.; Simoncini, T. d-Chiro-inositol regulate insulin in human adipocytes. Front. Endocrinol. 2021, 12, 660815. [Google Scholar] [CrossRef]
- Antonowski, T.; Osowski, A.; Lahuta, L.; Gorecki, R.; Rynkiewicz, A.; Wojtkiewicz, J. Healt-promoting properties of selected cyclitols for metabolic syndrome and diabetes. Nutrients 2019, 11, 2312. [Google Scholar] [CrossRef] [Green Version]
- Tari, S.H.; Sohouli, M.H.; Lari, A.; Fatahi, S.; Rahideh, S.T. The effect of inositol supplementation on blood pressure: A systematic review and meta-analysis of randomized-controlled trials. Clin. Nutr. ESPEN 2021, 44, 78–84. [Google Scholar] [CrossRef] [PubMed]
- Foster, S.R.; Dilworth, L.L.; Thompson, R.K.; Alexander-Lindo, R.L.; Omoruyi, F.O. Effects of combined inositol hexakisphosphate and inositol supplement on antioxidant activity and metabolic enzymes in the liver of streptozotocin-induced type 2 diabetic rats. Chem.-Biol. Interact. 2017, 275, 108–115. [Google Scholar] [CrossRef] [PubMed]
- Pintaudi, B.; Di Vieste, G.; Bonomo, M. The effectiveness of myo-Inositol and D-chiro inositol treatment in type 2 diabetes. Int. J. Endocrinol. 2016, 9132052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minambres, I.; Cuixart, G.; Goncalves, A.; Corcoy, R. Effects of inositol on glucose homeostasis: Systematic review and meta-analysis of randomized controlled trials. Clin. Nutr. 2019, 38, 1146–1152. [Google Scholar] [CrossRef]
- Santamaria, A.; Alibrandi, A.; Di Benedetto, A.; Pintaudi, B.; Corrado, F.; Facchinetti, F.; D’Anna, R. Clinical and metabolic outcomes in pregnant women at risk for gestational diabetes mellitus supplemented with myo-inositol: A secondary analysis from 3 RCTs. Am. J. Obstet. Gynecol. 2018, 219, 300.e1–300.e6. [Google Scholar] [CrossRef] [PubMed]
- Kulshrestha, V.; Balani, S.; Kachhawa, G.; Vanamail, P.; Kumari, R.; Sharma, J.B.; Bhatla, N. Efficacy of myo-inositol in treatment of gestational diabetes mellitus in Asian Indian women: A pilot randomized clinical trial. Eur. J. Obstet. Gynecol. Reprod. Biol. 2021, 260, 42–47. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Guo, S.; Miao, Z.; Li, Z.; Zhang, H. Myo-inositol lowers the risk of developing gestational diabetic mellitus in pregnancies: A systematic review and meta-analysis of randomized controlled trials with trial sequential analysis. J. Diabetes Its Complicat. 2018, 32, 342–348. [Google Scholar] [CrossRef]
- D’Anna, R.; Corrado, F.; Loddo, S.; Gullo, G.; Giunta, L.; Di Benedetto, A. Myo-inositol plus α-lactalbumin supplementation, insulin resistance and birth outcomes in women with gestational diabetes mellitus: A randomized, controlled study. Sci. Rep. 2021, 11, 8866. [Google Scholar] [CrossRef]
- Hernandez-Marin, I.; Picconi, I.; Laganà, A.S.; Costabile, L.; Unfer, V. A multicenter clinical study with myo-inositol and alpha-lactalbumin in Mexican and Italian PCOS patients. Eur. Rev. Med. Pharm. Sci. 2021, 25, 3316–3324. [Google Scholar]
- Baldassarre, M.P.A.; Di Tomo, P.; Centorame, G.; Pandolfi, A.; Di Pietro, N.; Consoli, A.; Formoso, G. Myo-inositol reduces inflammation and oxidative stress in human endothelial cells exposed in vivo to chronic hyperglycemia. Nutrients 2021, 13, 2210. [Google Scholar] [CrossRef]
- Surguchov, A. Caveolin: A new link between diabetes and AD. Cell. Mol. Neurobiol. 2020, 40, 1059–1066. [Google Scholar] [CrossRef]
- Lopez-Gambero, A.J.; Sanjuan, C.; Serrano-Castro, P.J.; Suarez, J.; de Fonseca, F.R. The biomedical uses of inositols: A nutraceutical approach to metabolic dysfunction in aging and neurodegenerative diseases. Biomedicines 2020, 8, 295. [Google Scholar] [CrossRef]
- Ramp, P.; Lehnert, A.; Matamouros, S.; Wirtz, A.; Baumgart, M.; Bott, M. Metabolic engineering of Corynebacterium glutamicum for production of scyllo-inositol, a drug candidate against Alzheimer’s disease. Metab. Eng. 2021, 67, 173–185. [Google Scholar] [CrossRef]
- McLaurin, J.; Kierstead, M.E.; Brown, M.E.; Hawks, C.A.; Lambermon, M.H.L.; Phinney, A.L.; Darabie, A.A.; Cousins, J.E.; French, J.E.; Lan, M.F.; et al. Cyclohexanehexol inhibitors of Ab aggregation prevent and reverse Alzheimer phenotype in a mouse model. Nat. Med. 2006, 12, 801–808. [Google Scholar] [CrossRef]
- Rafii, M.S.; Skotko, B.G.; McDonough, M.E.; Pulsifier, M.; Evans, C.; Doran, E.; Muranevici, G.; Kesslak, P.; Abushakra, S.; Lott, I.T. A randomized, double-blind, placebo-controlled, phase 2 study of oral ELND005 (scyllo-Inositol) in young adults with down syndrome without dementia. J. Alzheimers Dis. 2017, 58, 401–411. [Google Scholar] [CrossRef] [Green Version]
- Cantelmi, T.; Lambiase, E.; Unfer, V.R.; Gambioli, R.; Unfer, V. Inositol treatment for psychological symptoms in polycystic ovary syndrome women. Eur. Rev. Med. Pharm. Sci. 2021, 25, 2383–2389. [Google Scholar]
- Urrilla, A.S.; Hakkarainen, A.; Castaneda, A.; Paunio, T.; Marttunen, M.; Lundbom, N. Frontal cortex myo-inositol is associated with sleep and depression in adolescents: A proton magnetic resonance spectroscopy study. Neuropsychobiology 2017, 75, 21–31. [Google Scholar] [CrossRef] [Green Version]
- Brunst, K.J.; Ryan, P.H.; Altaye, M.; Yolton, K.; Maloney, T.; Beckwith, T.; Lemasters, G.; Cecil, K.M. Myo-inositol mediates the effects of traffic-related air pollution on generalized anxiety symptoms at age 12 years. Environ. Res. 2019, 175, 71–78. [Google Scholar] [CrossRef]
- Shirayama, Y.; Takahashi, M.; Osone, F.; Hara, A.; Okubo, T. Myo-inositol, glutamate, and glutamine in the prefrontal cortex, hippocampus, and amygdala in major depression. Biol. Psychiatry Cogn. Neurosci. Neuroimaging 2017, 2, 196–204. [Google Scholar] [CrossRef]
- Alonso-Castro, A.J.; Alba-Betancourt, C.; Rocha-Gonzalez, E.; Ruiz-Arredondo, A.; Zapata-Morales, J.R.; Gasca-Martinez, D.; Perez-Gutierrez, S. Neuropharmacological effects of d-pinitol and its possible mechanisms of action. J. Food Biochem. 2019, 43, e13070. [Google Scholar] [CrossRef]
- Gamkrelidze, G.N.; Nanobashvili, Z.I.; Bilanishvili, I.G.; Lordkipanidze, T.; Kandashvili, M.; Kokaia, M.; Solomonia, R.O. Concentration- and time-dependent effects of myo-inositol on evoked epileptic after discharge in the hippocampus in vivo. NeuroReport 2019, 30, 1129–1134. [Google Scholar] [CrossRef] [PubMed]
- Laganà, A.S.; Unfer, V.; Garzon, S.; Bizzarri, M. Role of inositol to improve surfactant functions and reduce IL-6 levels: A potential adjuvant strategy for SARS-CoV-2 pneumonia? Med. Hypotheses 2020, 144, 110262. [Google Scholar] [CrossRef] [PubMed]
- Bizzarri, M.; Laganà, A.S.; D’Aragona, D.; Unfer, V. Inositol and pulmonary function. Could inositol treatment down-regulate inflammation and cytokine release syndrome in SARS-CoV-2? Eur. Rev. Med. Pharm. Sci. 2020, 24, 3426–3432. [Google Scholar]
- Espinola, M.S.B.; Bertelli, M.; Bizzarri, M.; Unfer, V.; Laganà, A.S.; Visconti, B.; Aragona, C. Inositol and vitamin D may naturally protect human reproduction and women undergoing assisted reproduction from Covid-19 risk. J. Reprod. Immun. 2021, 144, 103271. [Google Scholar] [CrossRef] [PubMed]
- Wisniewski, K.; Jozwik, M.; Wojtkiewicz, J. Cancer prevention by natural products introduced into the diet—Selected cyclitols. Int. J. Mol. Sci. 2020, 21, 8988. [Google Scholar] [CrossRef] [PubMed]
- Vucenik, I.; Druzijanic, A.; Druzijanic, N. Inositol hexaphosphate (IP6) and colon cancer: From concepts and first experiments to clinical application. Molecules 2020, 25, 5931. [Google Scholar] [CrossRef]
- Weinberg, S.E.; Sun, L.Y.; Yang, A.L.; Liao, J.; Yang, G.Y. Overview of inositol and inositol phosphates on chemoprevention of colitis-induced carcinogenesis. Molecules 2021, 26, 31. [Google Scholar] [CrossRef]
- Bizzarri, M.; Dinicola, S.; Bevilacqua, A.; Cucina, A. Broad spectrum anticancer activity of myo-inositol and inositol hexakisphospate. Int. J. Endocrinol. 2016, 5616807. [Google Scholar]
- Unver, N.; Delgado, O.; Zeleke, K.; Cumpian, A.; Tang, X.; Caetano, M.S.; Wang, H.; Katayama, H.; Yu, H.; Szabo, E.; et al. Reduced IL-6 levels and tumor-associated phospho-STAT3 are associated with reduced tumor development in a mouse model of lung cancer chemoprevention with myo-inositol. Int. J. Cancer 2018, 142, 1405–1417. [Google Scholar] [CrossRef] [Green Version]
- El Hafni-Rahbi, B.; Brodaczewska, K.; Collet, G.; Majewska, A.; Klimkiewicz, K.; Delalande, A.; Grillon, C.; Kieda, C. Tumour angiogenesis normalized by myo-inositol trispyrophosphate alleviates hypoxia in the microenvironment and promotes antitumor immune response. J. Cell. Mol. Med. 2021, 25, 3284–3299. [Google Scholar] [CrossRef]
- Grgic, I.; Tschanz, F.; Borgeaud, N.; Gupta, A.; Clavien, P.A.; Guckenberger, M.; Graf, R.; Pruschy, M. Tumor oxygenation by myo-inositol trispyrophosphate enhances radiation response. Int. J. Radiat. Oncol. Biol. Phys. 2021, 110, 1222–1233. [Google Scholar] [CrossRef] [PubMed]
- Amabile, M.I.; De Luca, A.; Tripodi, D.; D’Alberti, E.; Melcarne, R.; Imbimbo, G.; Picconi, O.; D’Andrea, V.; Vergine, M.; Sorrenti, S.; et al. Effects of inositol hexaphosphate and myo-inositol administration in breast cancer patients during adjuvant chemotherapy. J. Pers. Med. 2021, 11, 756. [Google Scholar] [CrossRef] [PubMed]
- Proietti, S.; Pasta, V.; Cucina, A.; Aragona, C.; Palombi, E.; Vucenik, I.; Bizzarri, M. Inositol hexaphosphate (InsP6) as an effective topical treatment for patients receiving adjuvant chemotherapy after breast surgery. Eur. Rev. Med. Pharm. Sci. 2017, 21, 43–50. [Google Scholar]
- Pasta, V.; Dinicola, S.; Giuliani, A.; Harrat, A.H.; Alwasel, S.H.; Tartaglia, F.; Cucina, A.; Bizzarri, M. A randomized pilot study of inositol in association with betaine and boswellia in the management of mastalgia and benign breast lump in premenopausal women. Breast Cancer Basic Clin. Res. 2016, 10, 37–43. [Google Scholar] [CrossRef] [Green Version]
- Badodi, S.; Pomella, N.; Zhang, X.; Rosser, G.; Whittingham, J.; Niklison-Chirou, M.V.; Lim, Y.M.; Bradner, S.; Morrison, G.; Pollard, S.M.; et al. Inositol treatment inhibits medulloblastoma through suppression of epigenetic-driven metabolic adaptation. Nat. Commun. 2021, 12, 2148. [Google Scholar] [CrossRef]
- Ren, L.; Hong, E.S.; Mendoza, A.; Issaq, S.; Hoang, C.T.; Lizardo, M.; Le Blanc, A.; Khanna, C. Metabolomics uncovers a link between inositol metabolism and osteosarcoma metastasis. Oncotarget 2017, 8, 38541–38553. [Google Scholar] [CrossRef] [Green Version]
- Ferlay, J.; Soerjomataram, I.; Dikshit, R.; Eser, S.; Mathers, C.; Rebelo, M.; Parkin, D.M.; Forman, D.; Bray, F. Cancer incidence and mortality worldwide: Sources, methods and major patterns in GLOBOCAN 2012. Int. J. Cancer 2015, 136, E359–E386. [Google Scholar] [CrossRef]
- Kapral, M.; Wawszczyk, J.; Jesse, K.; Paul-Samojedny, M.; Kusmierz, D.; Weglarz, L. Inositol hexaphosphate inhibits proliferation and induces apoptosis of colon cancer cells by suppressing the AKT/mTOR signaling pathway. Molecules 2017, 22, 1657. [Google Scholar] [CrossRef] [Green Version]
- Yu, W.; Liu, C.; Li, X.; Yang, F.; Cheng, L.; Liu, C.; Song, Y. Inositol hexaphosphate suppresses colorectal cancer cell proliferation via the Akt/GSK-3β/β-catenin signaling cascade in a 1,2-dimethylhydrazine induced rat model. Eur. J. Pharm. 2017, 805, 67–74. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family | Species | Inositols | Refs. |
|---|---|---|---|
| Adoxaceae | Sambucus nigra | d-pinitol, allo-inositol, d-chiro-inositol, ononitol, bornesitol, scyllo-inositol, myo-inositol | [41] |
| Agaricaceae | Agaricus bisporus | d-pinitol, allo-inositol, d-chiro-inositol, bornesitol, scyllo-inositol, myo-inositol | [41] |
| Amarantaceae | Spinacia olearia | myo-inositol | [42] |
| Beta vulgaris | d-chiro-inositol, bornesitol, myo-inositol | [41,42] | |
| Amarylidaceae | Allium cepa | d-chiro-inositol, bornesitol, myo-inositol | [41,42] |
| Allium sativum | allo-inositol, ononitol, myo-inositol | [41] | |
| Allium ursinum | d-pinitol, d-chiro-inositol, ononitol, scyllo-inositol, myo-inositol | [41] | |
| Apiaceae | Anethum graveolens | ononitol, bornesitol, scyllo-inositol, myo-inositol | [41] |
| Apiaceae | Carum carvi | d-pinitol, d-chiro-inositol, bornesitol, scyllo-inosito, myo-inositol | [41] |
| Daucus carota | d-chiro-inositol, bornesitol, scyllo-inositol, myo-inositol | [41] | |
| Petroselinum crispum | d-pinitol, allo-inositol, d-chiro-inositol, bornesitol, scyllo-inositol, myo-inositol | [41] | |
| Apocynaceae | Hancornia speciosa | Bornesitol | [50] |
| Calendula anthodium | d-pinitol, d-chiro-inositol, bornesitol, scyllo-inositol, myo-inositol | [41] | |
| Asteraceae | Cichorium intybus | d-chiro-inositol, scyllo-inositol, myo-inositol | [42] |
| Cichorium endivia | d-chiro-inositol, scyllo-inositol, myo-inositol | [42] | |
| Cichorium endivia var. latifolia | d-chiro-inositol, myo-inositol | [42] | |
| Cynara cardunculus | d-chiro-inositol, scyllo-inositol, myo-inositol | [42] | |
| Lactuca sativa | d-chiro-inositol, myo-inositol | [42] | |
| Lactuca sativa var. crispa | d-chiro-inositol, myo-inositol | [42] | |
| Lactuca sativa ‘Lollo Rosso’ | d-chiro-inositol, myo-inositol | [42] | |
| Lactuca sativa var. longifolia | d-chiro-inositol, myo-inositol | [42] | |
| Lactuca sativa ‘Red Batavian’ | d-chiro-inositol, myo-inositol | [42] | |
| Matricaria chamomila | d-pinitol, chiro-inositol, neo-inositol, bornesitol, scyllo-inositol, myo-inositol | [48] | |
| Solidago virgaurea | d-pinitol, d-chiro-inositol, ononitol, bornesitol, scyllo-inositol, myo-inositol | [41] | |
| Tanacetum officinale | myo-inositol | [41] | |
| Taraxacum officinale | d-pinitol, d-chiro-inositol, bornesitol, myo-inositol | [41] | |
| Boraginaceae | Phacelia tanacetifolia | allo-inositol, scyllo-inositol, myo-inositol | [51] |
| Brassicaceae | Brassica oleracea | d-pinitol, d-chiro-inositol, bornesitol, myo-inositol | [41,42] |
| Brassica oleracea var. sabellica | d-pinitol, d-chiro-inositol, ononitol, myo-inositol | [41] | |
| Camelina sativa | d-pinitol, d-chiro-inositol, bornesitol, myo-inositol | [41] | |
| Raphanus raphanistrum subsp. sativus | myo-inositol | [42] | |
| Convolvulaceae | Ipomoea batatas | d-pinitol, d-chiro-inositol, bornesitol, scyllo-inositol, myo-inositol | [41] |
| Cupressaceae | Juniperus communis | d-pinitol, sequoytol | [49] |
| Dioscoreaceae | Dioscorea alata | scyllo-inositol, myo-inositol | [42] |
| Ericaceae | Vaccinium myrtillus | allo-inositol, d-chiro-inositol, ononitol, bornesitol, myo-inositol | [41] |
| Fabaceae | Arachis hypogea | d-pinitol, d-chiro-inositol, myo-inositol | [41] |
| Ceratonia silique | d-pinitol, allo-inositol, d-chiro-inositol, bornesitol, scyllo-inositol, myo-inositol | [41,43] | |
| Cicer arietinum | d-pinitol, d-chiro-inositol, myo-inositol, galactosyl-inositol, galactosyl-pinitol, ciceritol | [43,44] | |
| Erythrina edulis | d-chiro-inositol, myo-inositol | [44] | |
| Fagopyrum esculentum | d-chiro-inositol, myo-inositol, galactosyl-inositol, fagopyritol | [43] | |
| Glycine max | d-pinitol, d-chiro-inositol, myo-inositol, fagopyritol, galactosyl-pinitol | [43,44,54] | |
| Glycyrrhiza glabra | d-pinitol | [52] | |
| Latirus sativus | bornesitol, myo-inositol, galactosyl-inositol | [43] | |
| Lens culinaris | d-pinitol, d-chiro-inositol, bornesitol, myo-inositol, galactosyl-inositol, galactosyl-pinitol, ciceritol | [43,55] | |
| Lupinus perennis | d-pinitol, allo-inositol, d-chiro-inositol, bornesitol, myo-inositol | [41] | |
| Medicago sativa | d-pinitol, d-chiro-inositol, ononitol, scyllo-inositol, myo-inositol | [41,47] | |
| Phaseolus polyanthus | myo-inositol | [44] | |
| Phaseolus vilgaris | myo-inositol | [44,46] | |
| Pisum sativum | myo-inositol | [44] | |
| Trigonella foenum-graecum | d-pinitol, d-chiro-inositol, bornesitol, myo-inositol | [41] | |
| Vicia faba | myo-inositol | [44] | |
| Vicia sativa | myo-inostol | [44] | |
| Vigna unaniculata | ononitol, myo-inositol, galactosyl-inositol, galactosyl-ononitol | [43,44,45] | |
| Hydrophyllaceae | Phacella tanacetifolia | d-pinitol, allo-inositol, d-chiro-inositol, bornesitol, scyllo-inositol, myo-inositol | [41] |
| Hypericaceae | Hypericum perforatum | d-chiro-inositol, bornesitol, scyllo-inositol, myo-inositol | [41] |
| Lamiaceae | Mentha piperita | d-pinitol, d-chiro-inositol, ononitol, bornesitol, myo-inositol | [41] |
| Salvia officinalis | d-pinitol, d-chiro-inositol, bornesitol, scyllo-inositol, myo-inositol | [41] | |
| Lauraceae | Cinnamomum verum | allo-inositol, d-chiro-inositol, scyllo-inositol, myo-inositol | [41] |
| Laurus nobilis | d-pinitol, bornesitol, scyllo-inositol, myo-inositol | [41] | |
| Myristicaceae | Myristica fragrans | d-chiro-inositol, myo-inositol | [41] |
| Myrtaceae | Eugenia caryophyllus | allo-inositol, d-chiro-inositol, ononitol, bornesitol, scyllo-inositol, myo-inositol | [41] |
| Pinaceae | Abies sibirica | d-pinitol, sequoytol | [49] |
| Larix gmelinii | d-pinitol, sequoytol | [49] | |
| Picea abies | d-pinitol, sequoytol | [49] | |
| Pinus sibirica | d-pinitol, sequoytol | [49] | |
| Plumbaginaceae | Limonium perezii | d-chiro-inositol, myo-inositol | [56] |
| Limonium sinvatum | d-chiro-inositol, myo-inositol | [56] | |
| Poaceae | Oryza sativa | d-chiro-inositol, myo-inositol | [41] |
| Rosaceae | Rosa canina | d-pinitol, d-chiro-inositol, bornesitol, scyllo-inositol, myo-inositol | [41] |
| Sorbus aucuparia | d-pinitol, allo-inositol, d-chiro-inositol, bornesitol, scyllo-inositol, myo-inositol | [41] | |
| Solanaceae | Capsicum annuum | allo-inositol, d-chiro-inositol, bornesitol, scyllo-inositol, myo-inositol | [41] |
| Solanum melongena | scyllo-inositol, myo-inositol | [42] | |
| Solanum tuberosum | d-pinitol, d-chiro-inositol, bornesitol, myo-inositol | [41] | |
| Zingiberaceae | Curcuma longa | d-pinitol, d-chiro-inositol, bornesitol, myo-inositol | [41] |
| Elettaria cardamomum | d-pinitol, allo-inositol, d-chiro-inositol, ononitol, scyllo-inositol, myo-inositol | [41] | |
| Zingiber officinalis | d-pinitol, d-chiro-inositol, bornesitol, myo-inositol | [41] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Siracusa, L.; Napoli, E.; Ruberto, G. Novel Chemical and Biological Insights of Inositol Derivatives in Mediterranean Plants. Molecules 2022, 27, 1525. https://doi.org/10.3390/molecules27051525
Siracusa L, Napoli E, Ruberto G. Novel Chemical and Biological Insights of Inositol Derivatives in Mediterranean Plants. Molecules. 2022; 27(5):1525. https://doi.org/10.3390/molecules27051525
Chicago/Turabian StyleSiracusa, Laura, Edoardo Napoli, and Giuseppe Ruberto. 2022. "Novel Chemical and Biological Insights of Inositol Derivatives in Mediterranean Plants" Molecules 27, no. 5: 1525. https://doi.org/10.3390/molecules27051525
APA StyleSiracusa, L., Napoli, E., & Ruberto, G. (2022). Novel Chemical and Biological Insights of Inositol Derivatives in Mediterranean Plants. Molecules, 27(5), 1525. https://doi.org/10.3390/molecules27051525