
Cellular Effects of Cyclodextrins: Studies on HeLa Cells

 ,
,  ,
,  ,
, 
 ,
,  , ,
, , 
 and
and 
Abstract
:1. Introduction
2. Results
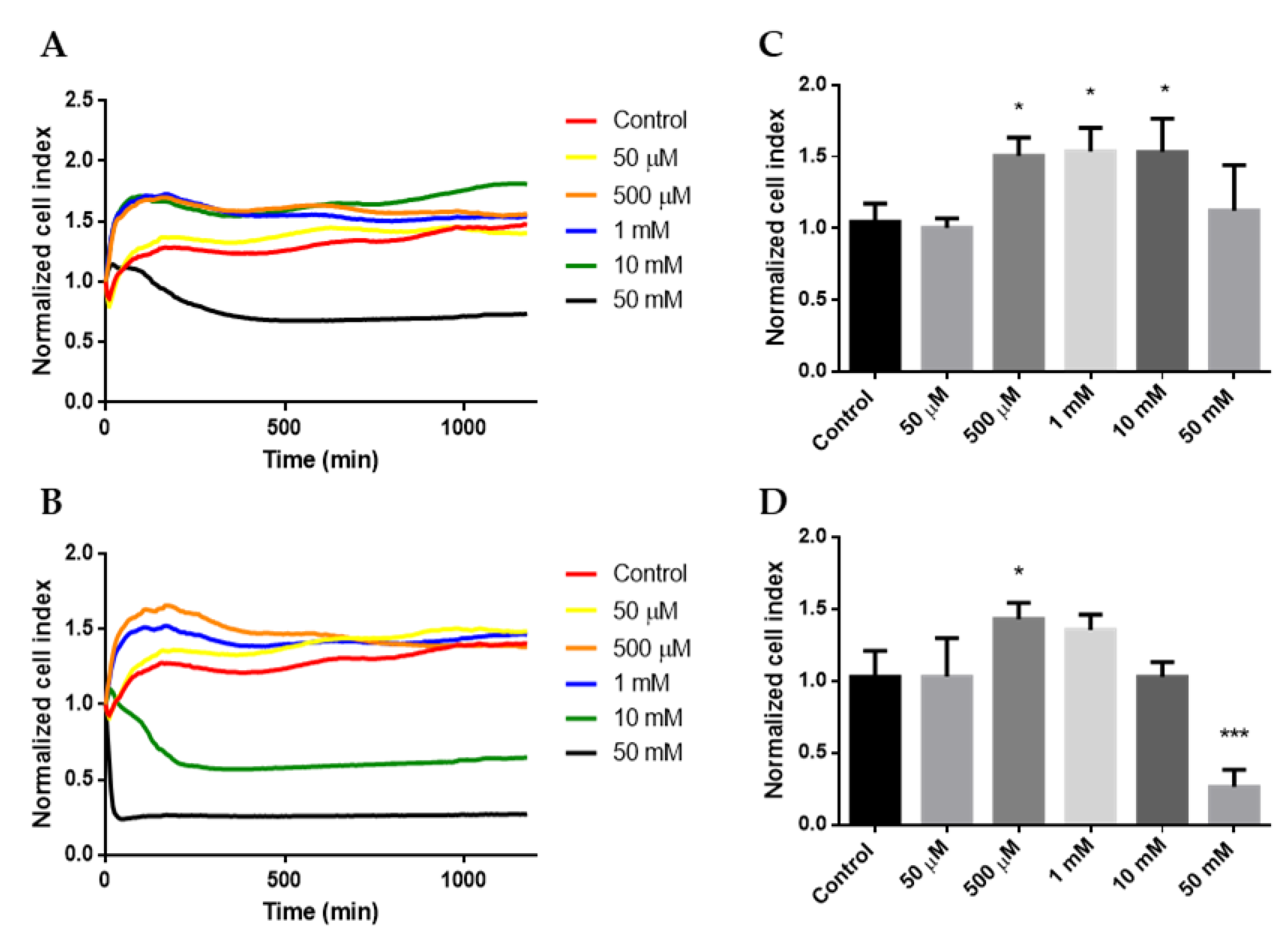
2.1. Cytotoxicity
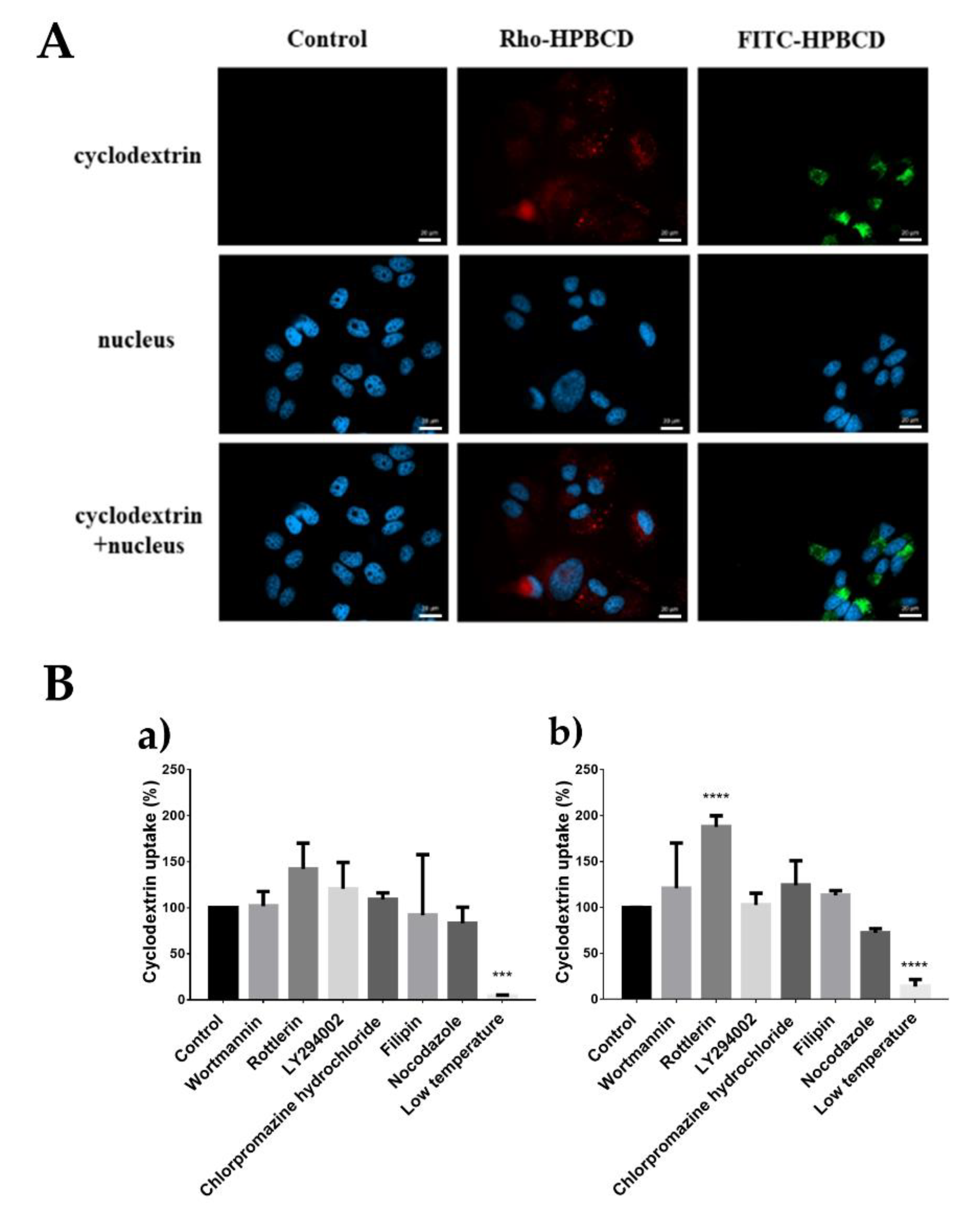
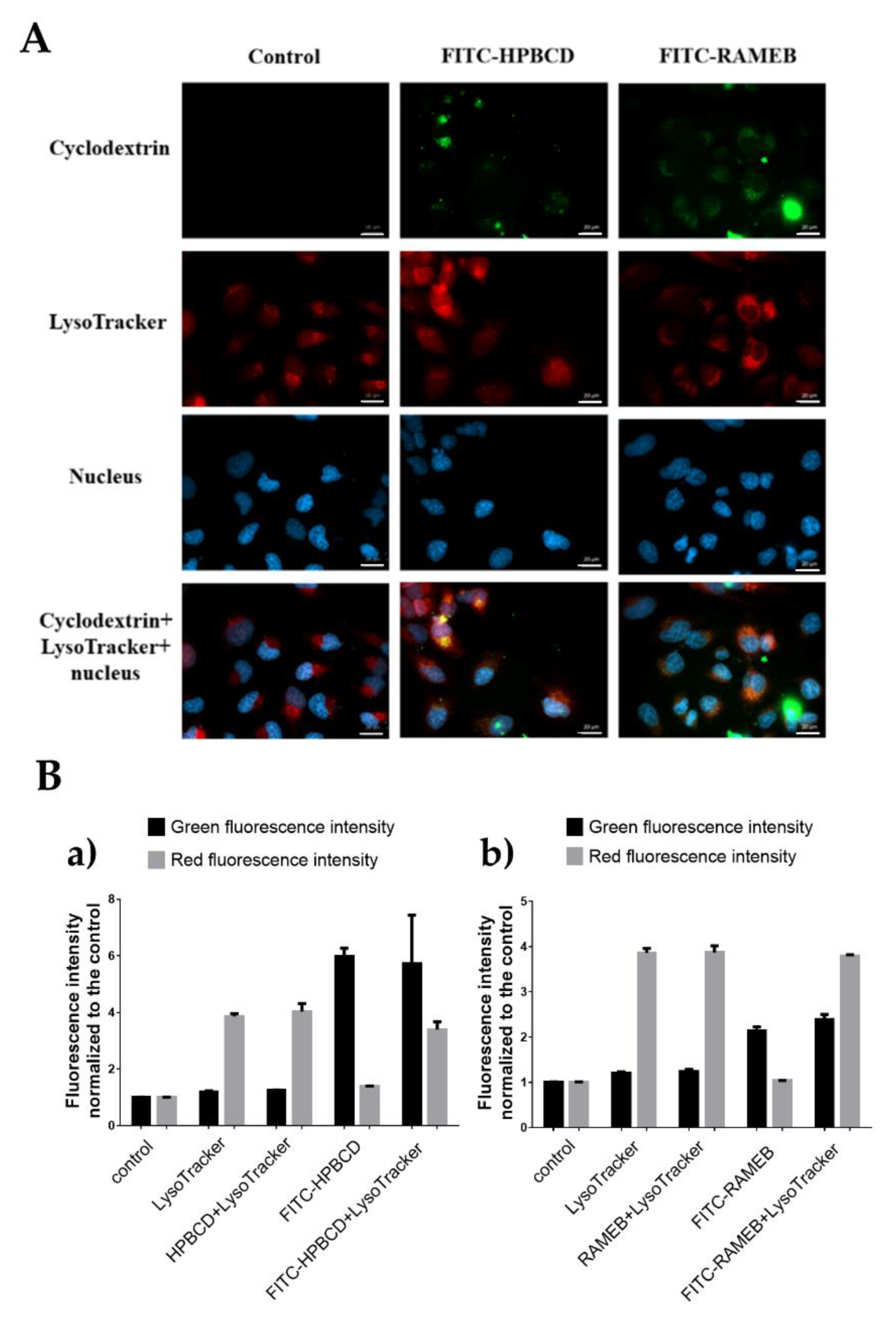
2.2. Investigation of the Cellular Uptake of Cyclodextrins
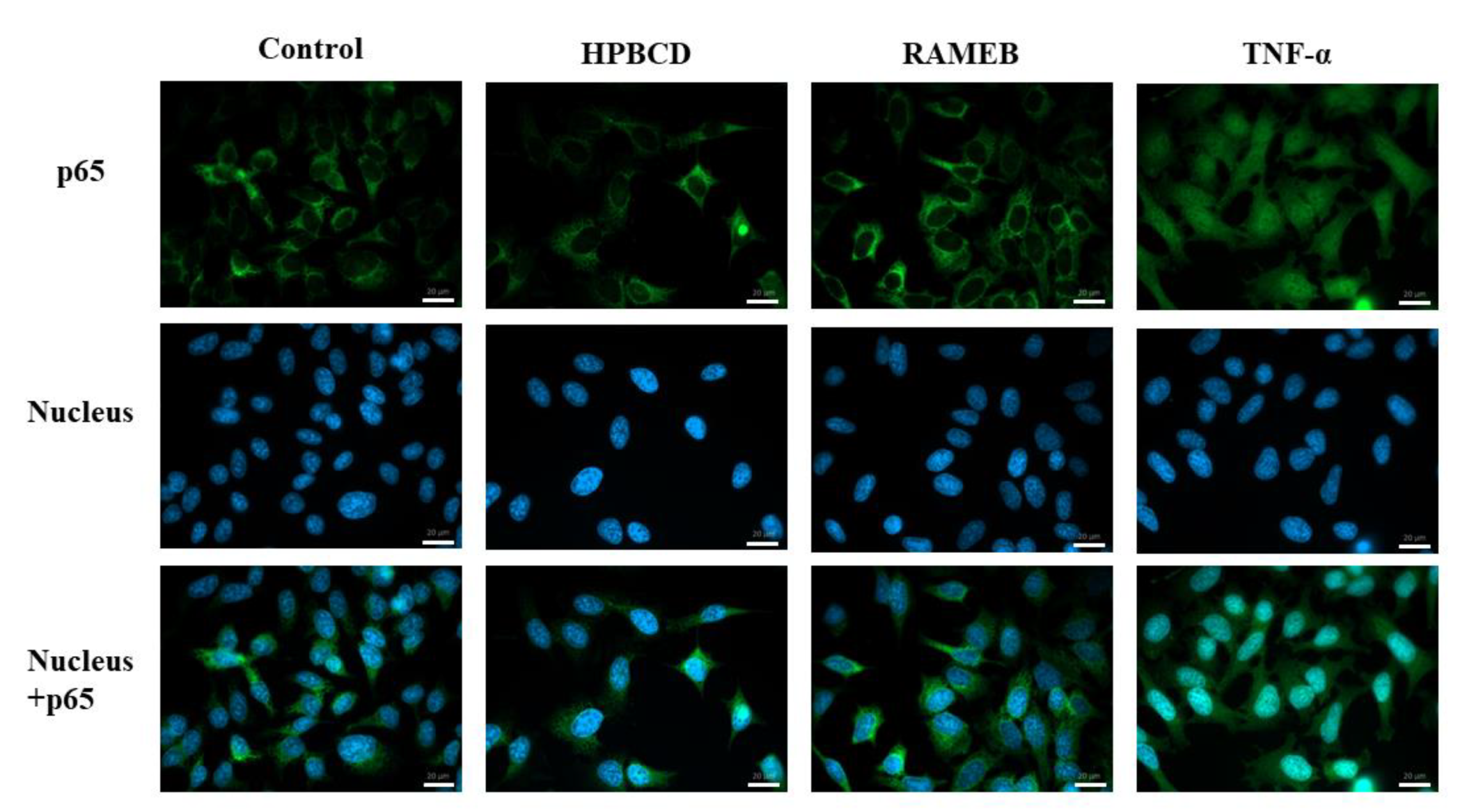
2.3. NF-κB Pathway Activation Study
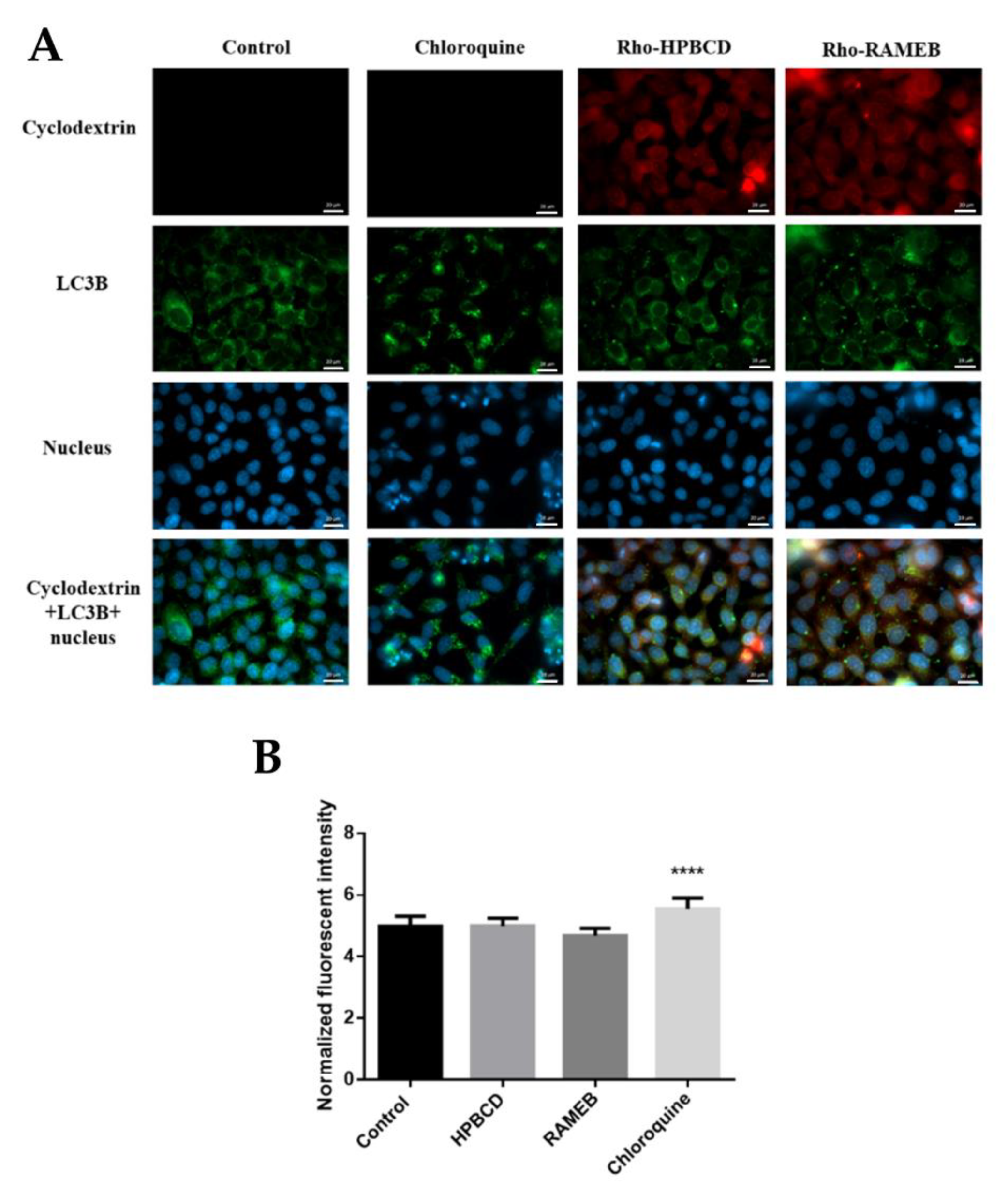
2.4. Autophagy Study
2.5. Investigation of Lysosomes
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cell Culture
4.3. Investigation of Cytotoxicity
4.4. Cellular Uptake of Fluorescently Labeled Cyclodextrin Derivatives on HeLa Cells
4.4.1. Fluorescence Microscopy
4.4.2. Flow Cytometry
4.5. Investigation of the NF-kB Pathway
4.6. Investigation of Autophagy
4.6.1. Fluorescence Microscopy
4.6.2. Microplate Reader
4.7. Investigation of the Lysosomes
4.7.1. Fluorescence Microscopy
4.7.2. Flow Cytometry
4.8. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Kurkov, S.V.; Loftsson, T. Cyclodextrins. Int. J. Pharm. 2013, 453, 167–180. [Google Scholar] [CrossRef] [PubMed]
- Szejtli, J. Past, present, and future of cyclodextrin research. Pure Appl. Chem. 2004, 76, 1825–1845. [Google Scholar] [CrossRef] [Green Version]
- Szejtli, J. Utilization of cyclodextrins in industrial products and processes. J. Mater. Chem. 1997, 7, 575–587. [Google Scholar] [CrossRef]
- Crini, G.; Fourmentin, S.; Fenyvesi, É.; Torri, G.; Fourmentin, M.; Morin-Crini, N. Cyclodextrins, from molecules to applications. Environ. Chem. Lett. 2018, 16, 1361–1375. [Google Scholar] [CrossRef]
- Kilsdonk, E.P.C.; Yancey, P.G.; Stoudt, G.W.; Bangerter, F.W.; Johnson, W.J.; Phillips, M.C.; Rothblat, G.H. Cellular cholesterol efflux mediated by cyclodextrins. J. Biol. Chem. 1995, 270, 17250–17256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiss, T.; Fenyvesi, F.; Bácskay, I.; Váradi, J.; Fenyvesi, É.; Iványi, R.; Szente, L.; Tósaki, Á.; Vecsernyés, M. Evaluation of the cytotoxicity of β-cyclodextrin derivatives: Evidence for the role of cholesterol extraction. Eur. J. Pharm. Sci. 2010, 40, 376–380. [Google Scholar] [CrossRef]
- Fenyvesi, F.; Fenyvesi, É.; Szente, L.; Goda, K.; Bacsó, Z.; Bácskay, I.; Váradi, J.; Kiss, T.; Molnár, É.; Janáky, T.; et al. P-glycoprotein inhibition by membrane cholesterol modulation. Eur. J. Pharm. Sci. 2008, 34, 236–242. [Google Scholar] [CrossRef]
- Rosenbaum, A.I.; Zhang, G.; Warren, J.D.; Maxfield, F.R. Endocytosis of beta-cyclodextrins is responsible for cholesterol reduction in Niemann-Pick type C mutant cells. Proc. Natl. Acad. Sci. USA 2010, 107, 5477–5482. [Google Scholar] [CrossRef] [Green Version]
- Ory, D.S.; Ottinger, E.A.; Farhat, N.Y.; King, K.A.; Jiang, X.; Weissfeld, L.; Berry-Kravis, E.; Davidson, C.D.; Bianconi, S.; Keener, L.A.; et al. Intrathecal 2-hydroxypropyl-β-cyclodextrin decreases neurological disease progression in Niemann-Pick disease, type C1: A non-randomised, open-label, phase 1–2 trial. Lancet 2017, 390, 1758–1768. [Google Scholar] [CrossRef] [Green Version]
- Vecsernyés, M.; Fenyvesi, F.; Bácskay, I.; Deli, M.A.; Szente, L.; Fenyvesi, É. Cyclodextrins, Blood–Brain Barrier, and Treatment of Neurological Diseases. Arch. Med. Res. 2014, 45, 711–729. [Google Scholar] [CrossRef]
- Zimmer, S.; Grebe, A.; Bakke, S.S.; Bode, N.; Halvorsen, B.; Ulas, T.; Skjelland, M.; De Nardo, D.; Labzin, L.I.; Kerksiek, A.; et al. Cyclodextrin promotes atherosclerosis regression via macrophage reprogramming. Sci. Transl. Med. 2016, 8, 333ra50. [Google Scholar] [CrossRef] [Green Version]
- Yokoo, M.; Kubota, Y.; Motoyama, K.; Higashi, T.; Taniyoshi, M.; Tokumaru, H.; Nishiyama, R.; Tabe, Y.; Mochinaga, S.; Sato, A.; et al. 2-Hydroxypropyl-β-cyclodextrin acts as a novel anticancer agent. PLoS ONE 2015, 10, 1–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Onodera, R.; Motoyama, K.; Tanaka, N.; Ohyama, A.; Okamatsu, A.; Higashi, T.; Kariya, R.; Okada, S.; Arima, H. Involvement of autophagy in antitumor activity of folate-appended methyl-β-cyclodextrin. Sci. Rep. 2014, 4, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rusznyák, Á.; Malanga, M.; Fenyvesi, É.; Szente, L.; Váradi, J.; Bácskay, I.; Vecsernyés, M.; Vasvári, G.; Haimhoffer, Á.; Fehér, P.; et al. Investigation of the Cellular Effects of Beta-Cyclodextrin Derivatives on Caco-2 Intestinal Epithelial Cells. Pharmaceutics 2021, 13, 157. [Google Scholar] [CrossRef] [PubMed]
- Fenyvesi, F.; Réti-Nagy, K.; Bacsó, Z.; Gutay-Tóth, Z.; Malanga, M.; Fenyvesi, É.; Szente, L.; Váradi, J.; Ujhelyi, Z.; Fehér, P.; et al. Fluorescently labeled methyl-beta-cyclodextrin enters intestinal epithelial Caco-2 cells by fluid-phase endocytosis. PLoS ONE 2014, 9, e84856. [Google Scholar] [CrossRef] [Green Version]
- Réti-Nagy, K.; Malanga, M.; Fenyvesi, É.; Szente, L.; Vámosi, G.; Váradi, J.; Bácskay, I.; Fehér, P.; Ujhelyi, Z.; Róka, E.; et al. Endocytosis of fluorescent cyclodextrins by intestinal Caco-2 cells and its role in paclitaxel drug delivery. Int. J. Pharm. 2015, 496, 509–517. [Google Scholar] [CrossRef] [Green Version]
- Plazzo, A.P.; Höfer, C.T.; Jicsinszky, L.; Fenyvesi, É.; Szente, L.; Schiller, J.; Herrmann, A.; Müller, P. Uptake of a fluorescent methyl-β-cyclodextrin via clathrin-dependent endocytosis. Chem. Phys. Lipids 2012, 165, 505–511. [Google Scholar] [CrossRef]
- Lawrence, T. The nuclear factor NF-kappaB pathway in inflammation. Cold Spring Harb. Perspect. Biol. 2009, 1, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Ueberla, K.; Lu, Y.; Chung, E.; Haseltine, W.A. The NF-kappa B p65 promoter. J. Acquir. Immune Defic. Syndr. 1993, 6, 227–230. [Google Scholar]
- Feng, Y.; He, D.; Yao, Z.; Klionsky, D.J. The machinery of macroautophagy. Cell Res. 2014, 24, 24–41. [Google Scholar] [CrossRef] [Green Version]
- Glick, D.; Barth, S.; Macleod, K.F. Autophagy: Cellular and molecular mechanisms. J. Pathol. 2020, 221, 3–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mizushima, N. Autophagy: Process and function. Genes Dev. 2007, 21, 2861–2873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, S.; Dulcey, A.E.; Hu, X.; Wassif, C.A.; Porter, F.D.; Austin, C.P.; Ory, D.S.; Marugan, J.; Zheng, W. Methyl-β-cyclodextrin restores impaired autophagy flux in Niemann-Pick C1-deficient cells through activation of AMPK. Autophagy 2017, 13, 1435–1451. [Google Scholar] [CrossRef] [PubMed]
- Ren, B.; Jiang, B.; Hu, R.; Zhang, M.; Chen, H.; Ma, J.; Sun, Y.; Jia, L.; Zheng, J. HP-β-cyclodextrin as an inhibitor of amyloid-β aggregation and toxicity. Phys. Chem. Chem. Phys. 2016, 18, 20476–20485. [Google Scholar] [CrossRef]
- Kovacs, T.; Sohajda, T.; Szente, L.; Nagy, P.; Panyi, G.; Varga, Z.; Zakany, F. Cyclodextrins Exert a Ligand-like Current Inhibitory Effect on the KV1.3 Ion Channel Independent of Membrane Cholesterol Extraction. Front. Mol. Biosci. 2021, 8, 1–11. [Google Scholar] [CrossRef]
- Róka, E.; Ujhelyi, Z.; Deli, M.; Bocsik, A.; Fenyvesi, É.; Szente, L.; Fenyvesi, F.; Vecsernyés, M.; Váradi, J.; Fehér, P.; et al. Evaluation of the Cytotoxicity of α-Cyclodextrin Derivatives on the Caco-2 Cell Line and Human Erythrocytes. Molecules 2015, 20, 20269–20285. [Google Scholar] [CrossRef] [Green Version]
- Sarkar, K.; Kruhlak, M.J.; Erlandsen, S.L.; Shaw, S. Selective inhibition by rottlerin of macropinocytosis in monocyte-derived dendritic cells. Immunology 2005, 116, 513–524. [Google Scholar] [CrossRef]
- Song, J.; Zhou, Y.; Gong, Y.; Liu, H.; Tang, L. Rottlerin promotes autophagy and apoptosis in gastric cancer cell lines. Mol. Med. Rep. 2018, 18, 2905–2913. [Google Scholar] [CrossRef]
- Falcone, S.; Cocucci, E.; Podini, P.; Kirchhausen, T.; Clementi, E.; Meldolesi, J. Macropinocytosis: Regulated coordination of endocytic and exocytic membrane traffic events. J. Cell Sci. 2006, 119, 4758–4769. [Google Scholar] [CrossRef] [Green Version]
- O’Neill, M.J.; Guo, J.; Byrne, C.; Darcy, R.; O’Driscoll, C.M. Mechanistic studies on the uptake and intracellular trafficking of novel cyclodextrin transfection complexes by intestinal epithelial cells. Int. J. Pharm. 2011, 413, 174–183. [Google Scholar] [CrossRef] [Green Version]
- Ivanov, A.I. Pharmacological inhibition of endocytic pathways: Is it specific enough to be useful? Methods Mol. Biol. 2008, 440, 15–33. [Google Scholar] [CrossRef] [PubMed]
- Motoyama, K.; Arima, H.; Nishimoto, Y.; Miyake, K.; Hirayama, F.; Uekama, K. Involvement of CD14 in the inhibitory effects of dimethyl-α-cyclodextrin on lipopolysaccharide signaling in macrophages. FEBS Lett. 2005, 579, 1707–1714. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Caco-2 Cell Line | HeLa Cell Line | |
|---|---|---|
| Cytotoxicity of cyclodextrins | 50 mM HPBCD and 10 mM RAMEB were cytotoxic (RTCA method) | |
| Cellular internalization of fluorescently labeled cyclodextrins | Both fluorescein and rhodamine labeled HPBCD and RAMEB were localized in vesicles in the cytoplasm. (FM) | |
| Normalized fluorescence intensity (FC, 50 µM): FITC-HPBCD: 35.63 ± 2.72 FITC-RAMEB: 4.96 ± 1.23 | Normalized fluorescence intensity (FC, 50 µM): FITC-HPBCD: 8.55 ± 2.26 FITC-RAMEB: 2.13 ± 0.33 | |
| The internalization of cyclodextrins was significantly inhibited at 0 °C. (FC) | ||
| Rottlerin significantly reduced the endocytosis both of FITC-HPBCD and FITC-RAMEB. (FC) | Rottlerin significantly increased the internalization of FITC-RAMEB. (FC) | |
| Chlorpromazine significantly increased the internalization of the cyclodextrins. (FC) | Chlorpromazine had no significant effect on the cellular uptake of cyclodextrins. (FC) | |
| Type of cyclodextrin endocytosis: fluid phase endocytosis predominates. | Type of endocytosis: its precise determination requires further experiments, presumably several simultaneous processes including clathrin-dependent endocytosis | |
| Investigation of lysosomes | Cyclodextrin tested: Rho-HPBCD, Rho-RAMEB | Cyclodextrins tested: FITC-HPBCD, FITC-RAMEB |
| Cyclodextrins entered the lysosomes, did not show more intense staining compared to the control and did not induce the formation of lysosomes. (FM, FC) | ||
| Investigation of the NF-κB pathway | p65 translocation was not detected to the nucleus and induction of the NF-κB inflammatory pathway. (FM) | |
| Investigation of the autophagy | Based on qualitative and quantitative studies, neither Rho-HPBCD nor Rho-RAMEB induced the formation of autophagosomes and autophagy. The effect of chloroquine (used as a positive control) was significantly different from that of the control sample. (FM, MR) | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rusznyák, Á.; Palicskó, M.; Malanga, M.; Fenyvesi, É.; Szente, L.; Váradi, J.; Bácskay, I.; Vecsernyés, M.; Réti-Nagy, K.S.; Vasvári, G.; et al. Cellular Effects of Cyclodextrins: Studies on HeLa Cells. Molecules 2022, 27, 1589. https://doi.org/10.3390/molecules27051589
Rusznyák Á, Palicskó M, Malanga M, Fenyvesi É, Szente L, Váradi J, Bácskay I, Vecsernyés M, Réti-Nagy KS, Vasvári G, et al. Cellular Effects of Cyclodextrins: Studies on HeLa Cells. Molecules. 2022; 27(5):1589. https://doi.org/10.3390/molecules27051589
Chicago/Turabian StyleRusznyák, Ágnes, Mercédesz Palicskó, Milo Malanga, Éva Fenyvesi, Lajos Szente, Judit Váradi, Ildikó Bácskay, Miklós Vecsernyés, Katalin Szászné Réti-Nagy, Gábor Vasvári, and et al. 2022. "Cellular Effects of Cyclodextrins: Studies on HeLa Cells" Molecules 27, no. 5: 1589. https://doi.org/10.3390/molecules27051589
APA StyleRusznyák, Á., Palicskó, M., Malanga, M., Fenyvesi, É., Szente, L., Váradi, J., Bácskay, I., Vecsernyés, M., Réti-Nagy, K. S., Vasvári, G., Haimhoffer, Á., & Fenyvesi, F. (2022). Cellular Effects of Cyclodextrins: Studies on HeLa Cells. Molecules, 27(5), 1589. https://doi.org/10.3390/molecules27051589