
The Antagonizing Role of Heme in the Antimalarial Function of Artemisinin: Elevating Intracellular Free Heme Negatively Impacts Artemisinin Activity in Plasmodium falciparum


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
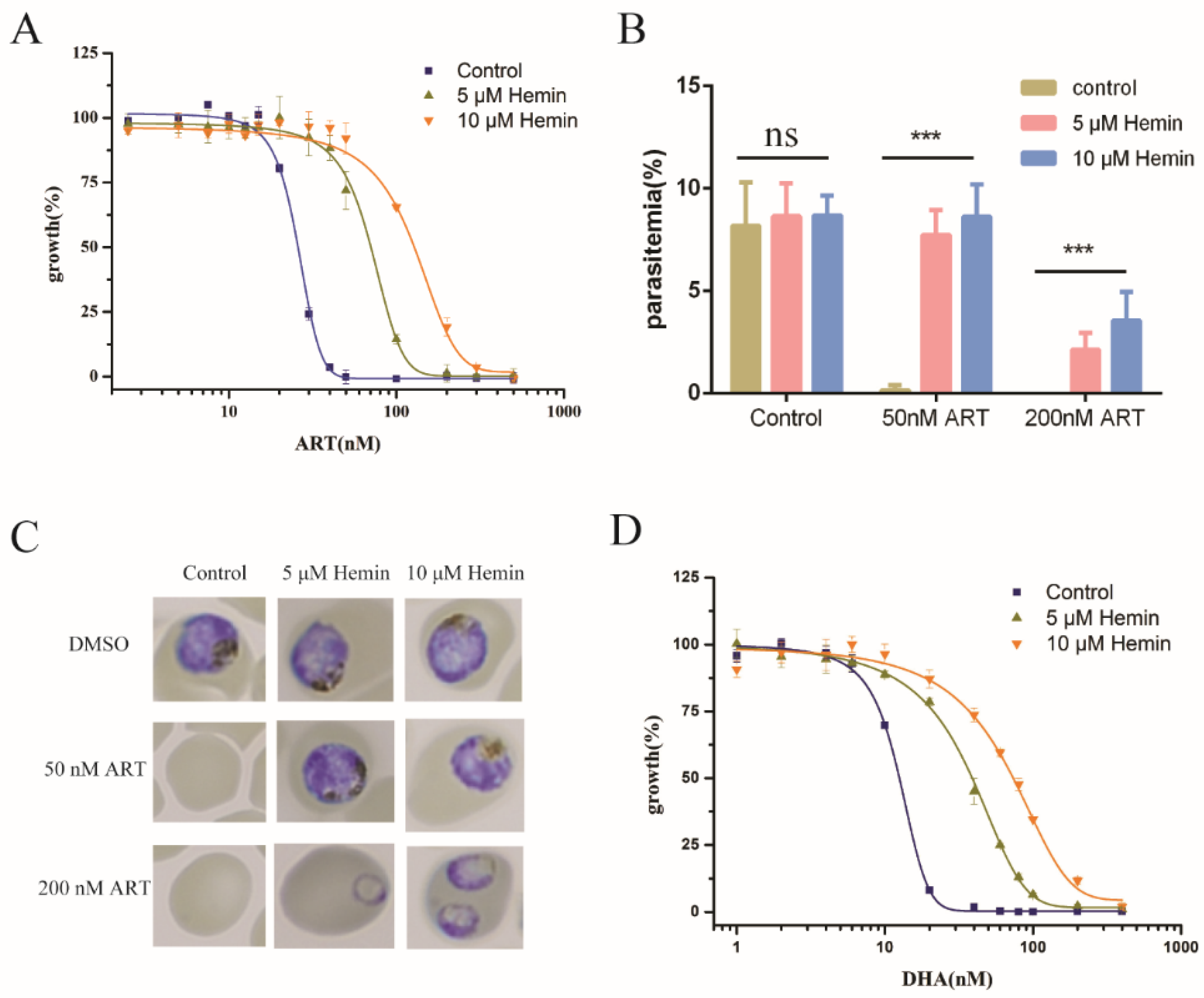
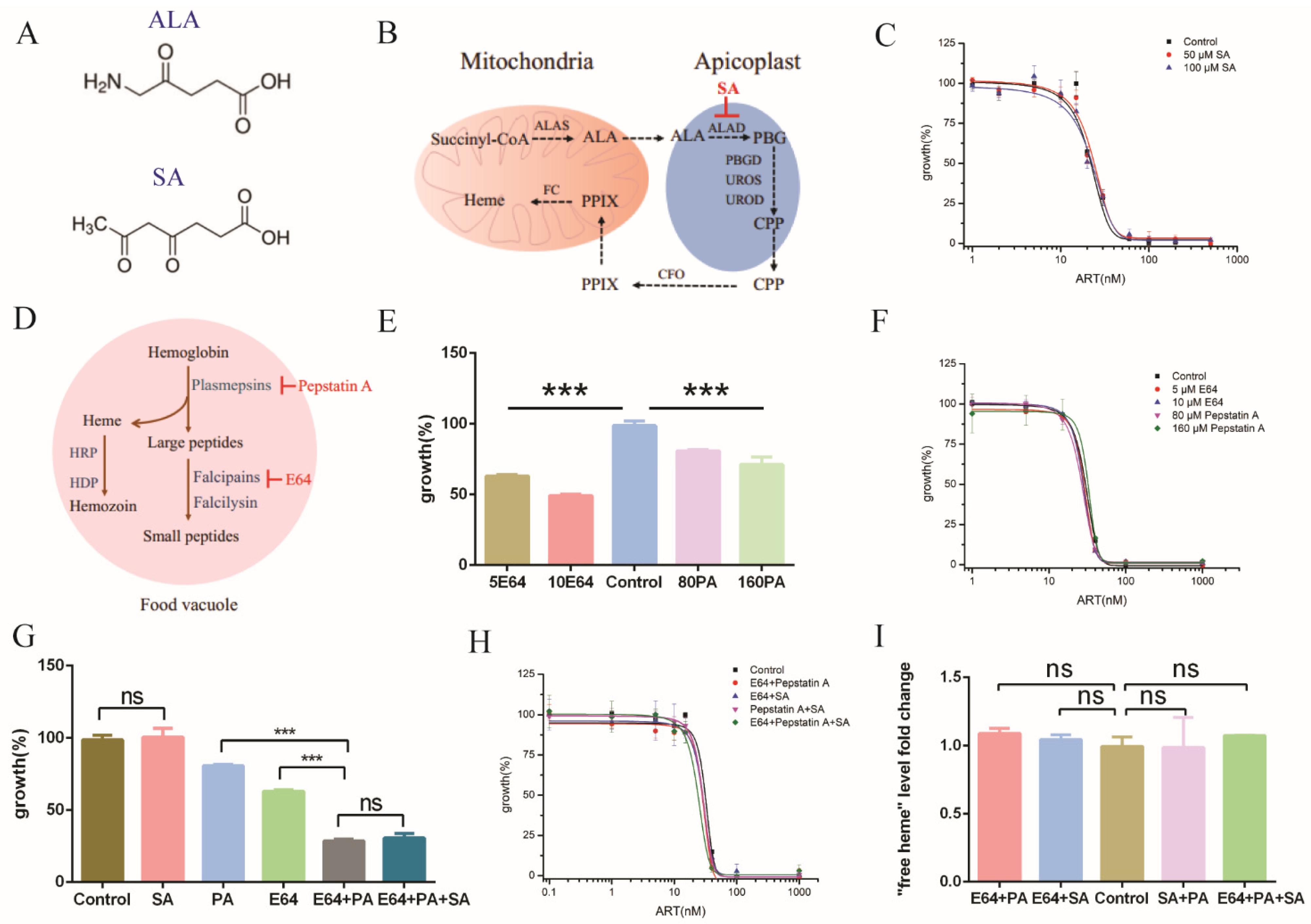
2.1. Extracellular Hemin Reduced the Antimalarial Activity of Artemisinin
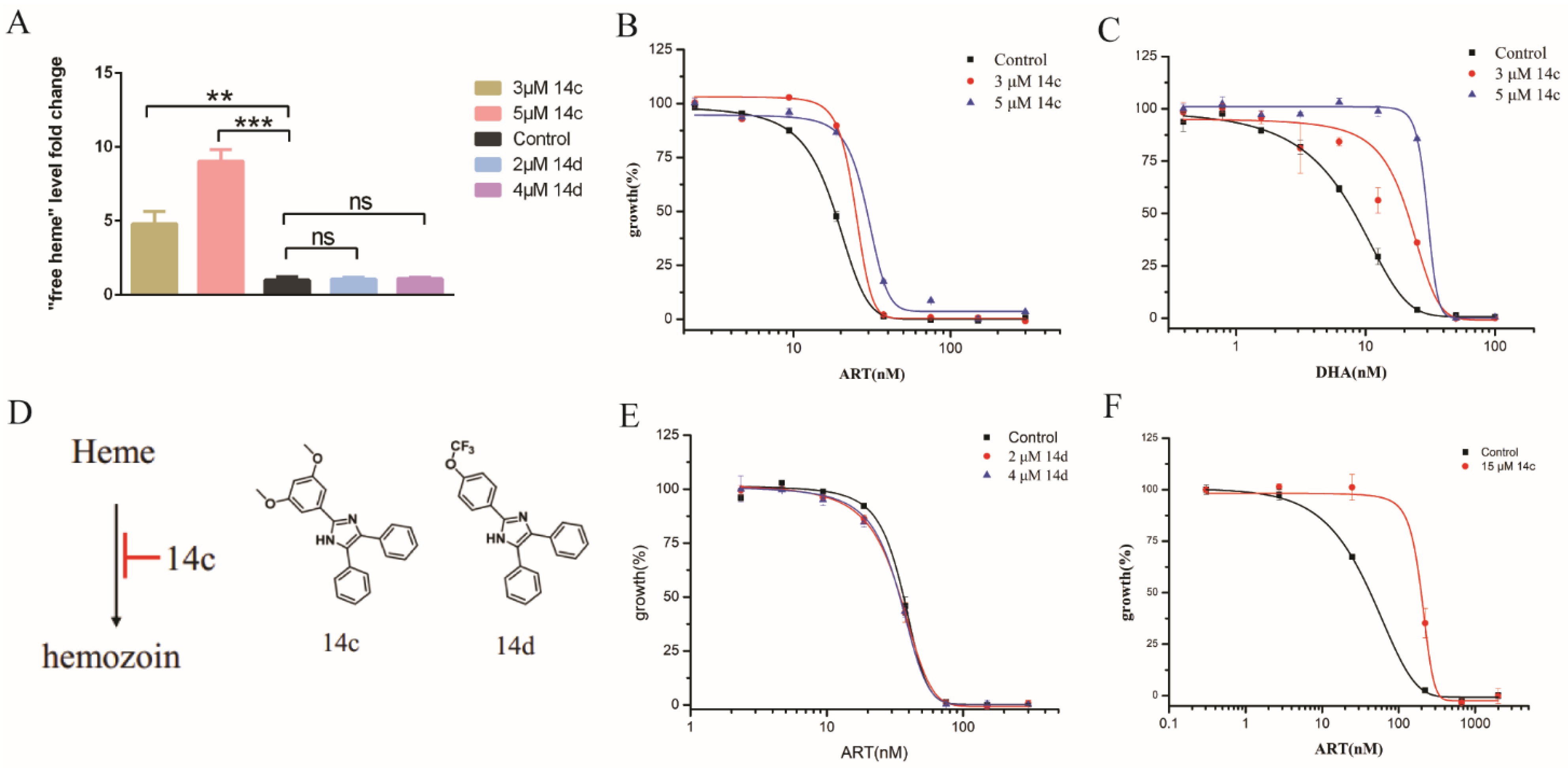
2.2. Inhibiting Hemozoin Formation Antagonized Artemisinin’s Action
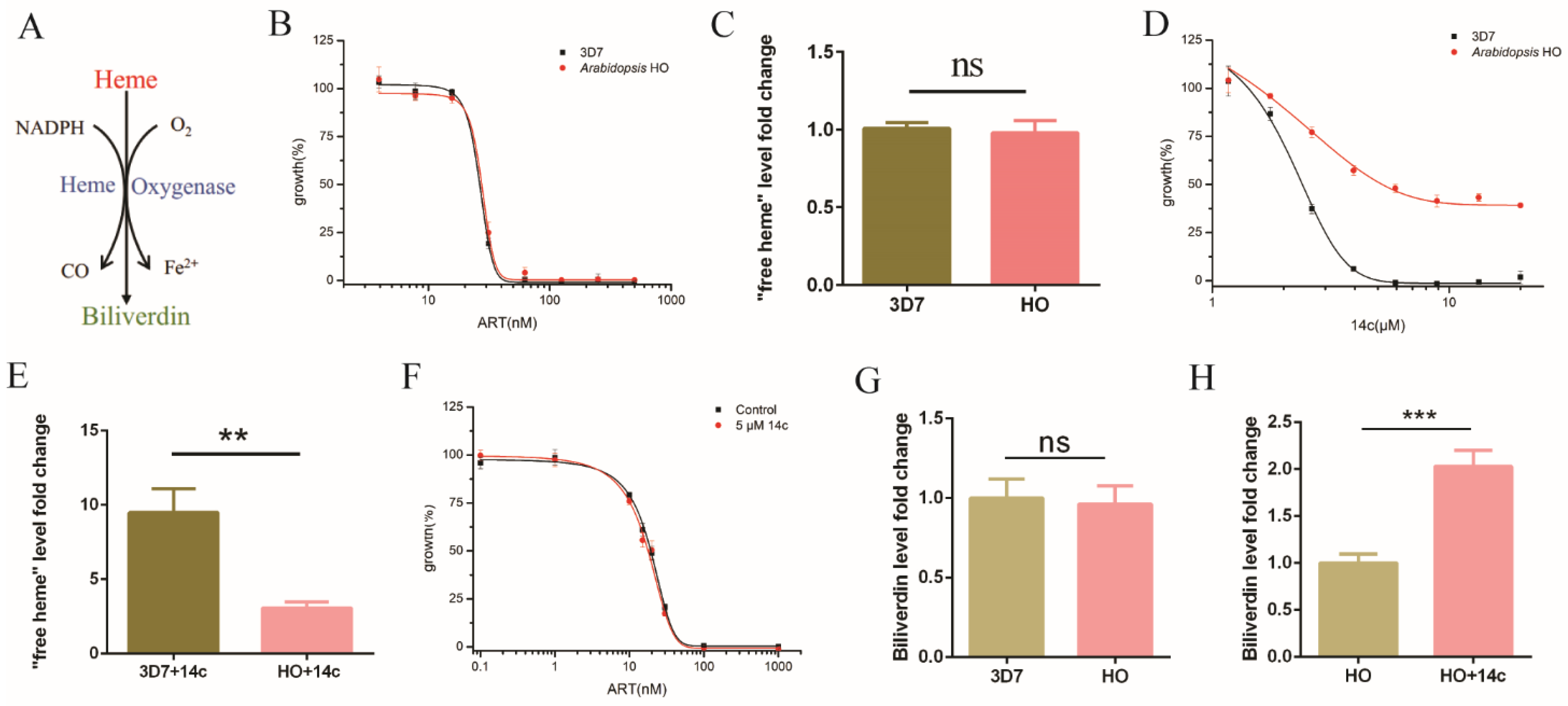
2.3. Heme Oxygenase Suppressed the Antagonizing Effect of 14c on Artemisinin’s Action
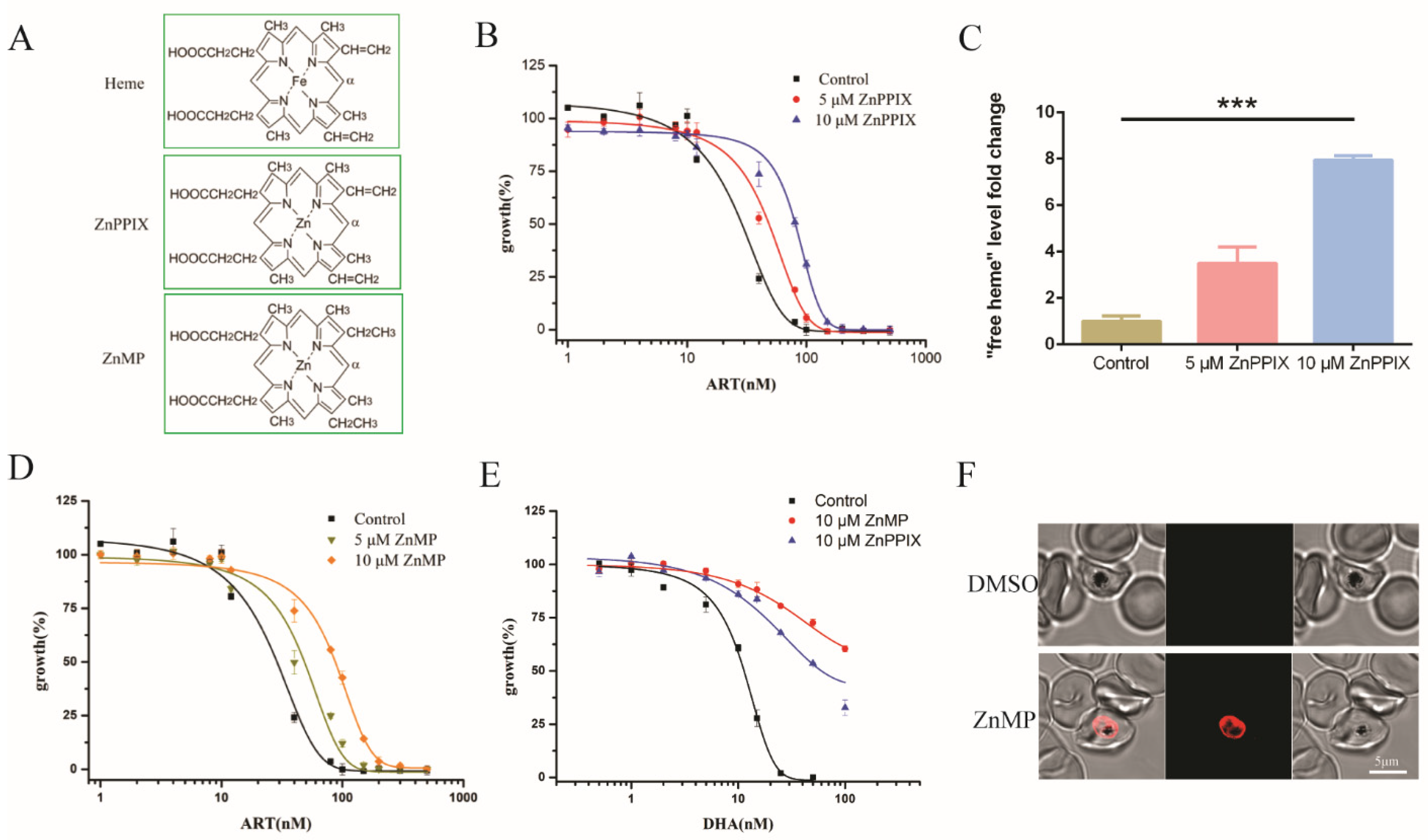
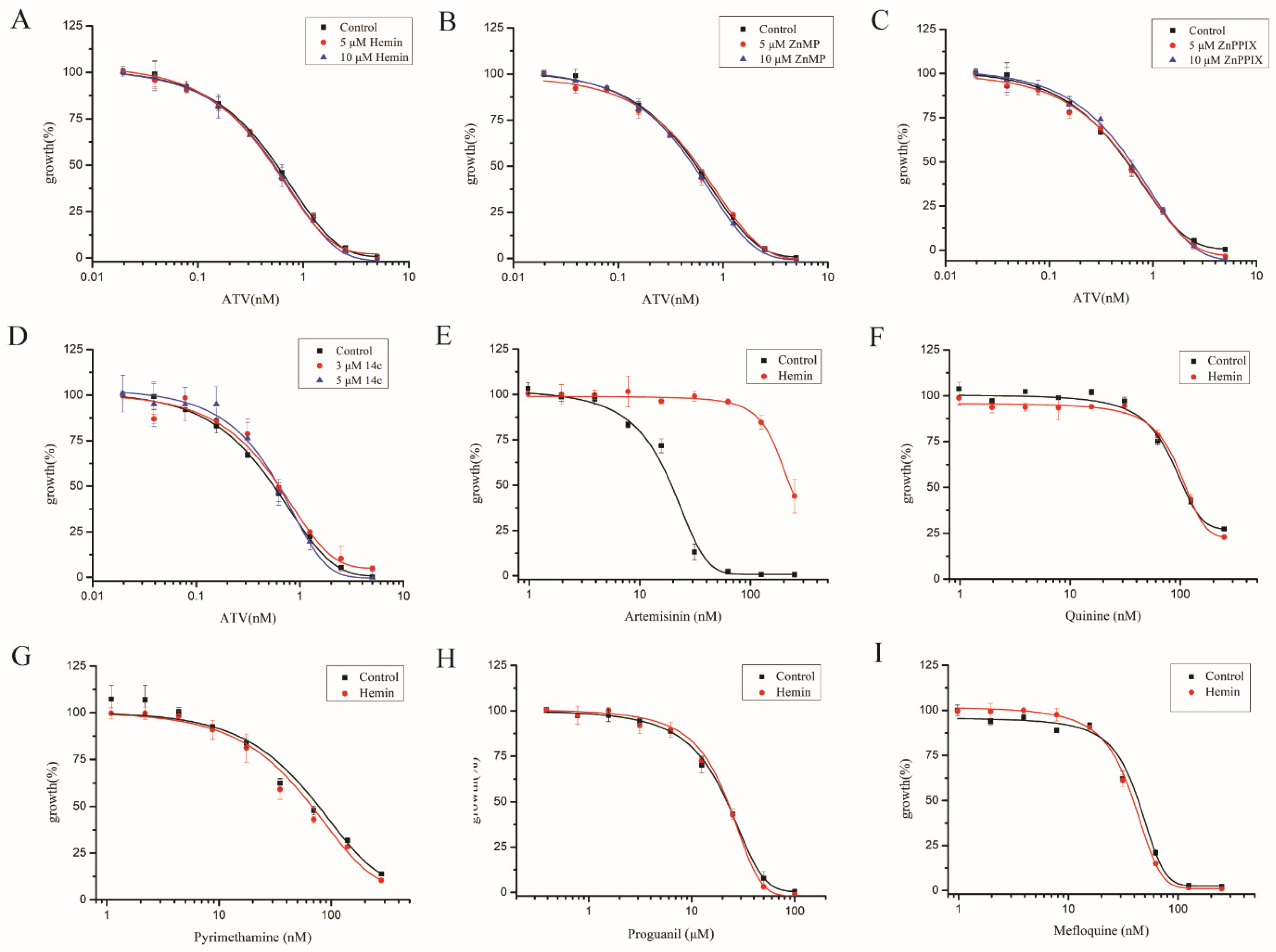
2.4. Enhancing Heme Levels by Extracellular Heme Analogue also Reduced the Antimalarial Activity of Artemisinin
2.5. Inhibiting de Novo Heme Biosynthesis and Hemoglobin Degradation Had Little Effect on Free Heme and the Action of Artemisinin
2.6. Elevating Heme Level Had Little Effect on Other Antimalarial Drugs
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Parasites Culture
4.3. Inhibition Assay
4.4. Heme Fractionation and Measurement
4.5. Confocal Microscopy
4.6. P. falciparum Transfection
4.7. LC-MS/MS Quantification of Biliverdin
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- WHO. World Malaria Report 2018; WHO: Geneva, Switzerland, 2018. [Google Scholar]
- Björkman, A.; Phillips-Howard, P.A. The epidemiology of drug-resistant malaria. Trans. R. Soc. Trop. Med. Hyg. 1990, 84, 177–180. [Google Scholar] [CrossRef] [Green Version]
- Wernsdorfer, W.H.; Payne, D. The Dynamics of Drug Resistance in Plasmodium Falciparum. Pharmacol. Ther. 1991, 50, 95–121. [Google Scholar] [CrossRef]
- Wongsrichanalai, C.; Pickard, A.L.; Wernsdorfer, W.H.; Meshnick, S.R. Epidemiology of drug-resistant malaria. Lancet Infect. Dis. 2002, 2, 209–218. [Google Scholar] [CrossRef]
- Klayman, D.L. Qinghaosu (artemisinin): An antimalarial drug from China. Science 1985, 228, 1049–1055. [Google Scholar] [CrossRef] [Green Version]
- Tu, Y. The discovery of artemisinin (qinghaosu) and gifts from Chinese medicine. Nat. Med. 2011, 17, 1217–1220. [Google Scholar] [CrossRef]
- Li, J.; Zhou, B. Biological actions of artemisinin: Insights from medicinal chemistry studies. Molecules 2010, 15, 1378–1397. [Google Scholar] [CrossRef]
- Noedl, H.; Se, Y.; Schaecher, K.; Smith, B.L.; Socheat, D.; Fukuda, M.M. Evidence of Artemisinin-Resistant Malaria in Western Cambodia. N. Engl. J. Med. 2008, 359, 2619–2620. [Google Scholar] [CrossRef]
- Dondorp, A.M.; Nosten, F.; Yi, P.; Das, D.; Phyo, A.P.; Tarning, J.; Lwin, K.M.; Ariey, F.; Hanpithakpong, W.; Lee, S.J.; et al. Artemisinin Resistance in Plasmodium falciparum Malaria. N. Engl. J. Med. 2009, 361, 455–467. [Google Scholar] [CrossRef] [Green Version]
- Ariey, F.; Witkowski, B.; Amaratunga, C.; Beghain, J.; Langlois, A.-C.; Khim, N.; Kim, S.; Duru, V.; Bouchier, C.; Ma, L.; et al. A molecular marker of artemisinin-resistant Plasmodium falciparum malaria. Nature 2014, 505, 50–55. [Google Scholar] [CrossRef]
- Ashley, E.A.; Dhorda, M.; Fairhurst, R.M.; Amaratunga, C.; Lim, P.; Suon, S.; Sreng, S.; Anderson, J.M.; Mao, S.; Sam, B.; et al. Spread of artemisinin resistance in Plasmodium falciparum malaria. N. Engl. J. Med. 2014, 371, 411–423. [Google Scholar] [CrossRef] [Green Version]
- Witkowski, B.; Amaratunga, C.; Khim, N.; Sreng, S.; Chim, P.; Kim, S.; Lim, P.; Mao, S.; Sopha, C.; Sam, B.; et al. Novel phenotypic assays for the detection of artemisinin-resistant Plasmodium falciparum malaria in Cambodia: In-vitro and ex-vivo drug-response studies. Lancet Infect. Dis. 2013, 13, 1043–1049. [Google Scholar] [CrossRef] [Green Version]
- O’Neill, P.M.; Barton, V.E.; Ward, S.A. The molecular mechanism of action of artemisinin—The debate continues. Molecules 2010, 15, 1705–1721. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, P.M.; Posner, G.H. A Medicinal Chemistry Perspective on Artemisinin and Related Endoperoxides. J. Med. Chem. 2004, 47, 2945–2964. [Google Scholar] [CrossRef] [PubMed]
- Golenser, J.; Domb, A.; Leshem, B.; Kremsner, P.; Luty, A. Iron chelators as drugs against malaria pose a potential risk. Redox Rep. 2003, 8, 268–271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stocks, P.A.; Bray, P.G.; Barton, V.E.; Al-Helal, M.; Jones, M.; Araujo, N.C.; Gibbons, P.; Ward, S.A.; Hughes, R.H.; Biagini, G.A.; et al. Evidence for a common non-heme chelatable-iron-dependent activation mechanism for semisynthetic and synthetic endoperoxide antimalarial drugs. Angew. Chem. Int. Ed. Engl. 2007, 46, 6278–6283. [Google Scholar] [CrossRef]
- Sibmooh, N.; Udomsangpetch, R.; Kijjoa, A.; Chantharaksri, U.; Mankhetkorn, S. Redox Reaction of Artemisinin with Ferrous and Ferric Ions in Aqueous Buffer. Chem. Pharm. Bull. 2001, 49, 1541–1546. [Google Scholar] [CrossRef] [Green Version]
- Robert, A.; Coppel, Y.; Meunier, B. Alkylation of heme by the antimalarial drug artemisinin. Chem. Commun. 2002, 5, 414–415. [Google Scholar] [CrossRef]
- Zhang, S.; Gerhard, G.S. Heme activates artemisinin more efficiently than hemin, inorganic iron, or hemoglobin. Bioorg. Med. Chem. 2008, 16, 7853–7861. [Google Scholar] [CrossRef]
- Naimin, W.; Sadrzadeh, S.M.H. Enhancement of hemin-induced membrane damage by artemisinin. Biochem. Pharmacol. 1994, 48, 737–741. [Google Scholar] [CrossRef]
- Robert, A.; Benoit-Vical, F.; Claparols, C.; Meunier, B. The antimalarial drug artemisinin alkylates heme in infected mice. Proc. Natl. Acad. Sci. USA 2005, 102, 13676–13680. [Google Scholar] [CrossRef] [Green Version]
- Klonis, N.; Crespo-Ortiz, M.P.; Bottova, I.; Abu-Bakar, N.; Kenny, S.; Rosenthal, P.J.; Tilley, L. Artemisinin activity against Plasmodium falciparum requires hemoglobin uptake and digestion. Proc. Natl. Acad. Sci. USA 2011, 108, 11405–11410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, S.C.; Dogovski, C.; Hanssen, E.; Chiu, F.; Yang, T.; Crespo, M.P.; Stafford, C.; Batinovic, S.; Teguh, S.; Charman, S.; et al. Haemoglobin degradation underpins the sensitivity of early ring stage Plasmodium falciparum to artemisinins. J. Cell Sci. 2016, 129, 406–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, C.; Li, J.; Cao, Y.; Long, G.; Zhou, B. Two distinct and competitive pathways confer the cellcidal actions of artemisinins. Microb. Cell 2015, 2, 14–25. [Google Scholar] [CrossRef] [Green Version]
- Ponmee, N.; Chuchue, T.; Wilairat, P.; Yuthavong, Y.; Kamchonwongpaisan, S. Artemisinin effectiveness in erythrocytes is reduced by heme and heme-containing proteins. Biochem. Pharm. 2007, 74, 153–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sigala, P.A.; Crowley, J.R.; Henderson, J.P.; Goldberg, D.E. Deconvoluting heme biosynthesis to target blood-stage malaria parasites. Elife 2015, 4, e09143. [Google Scholar] [CrossRef] [PubMed]
- Wicht, K.J.; Combrinck, J.M.; Smith, P.J.; Hunter, R.; Egan, T.J. Identification and Mechanistic Evaluation of Hemozoin-Inhibiting Triarylimidazoles Active against Plasmodium falciparum. ACS Med. Chem. Lett. 2017, 8, 201–205. [Google Scholar] [CrossRef] [Green Version]
- Sigala, P.A.; Crowley, J.R.; Hsieh, S.; Henderson, J.P.; Goldberg, D.E. Direct tests of enzymatic heme degradation by the malaria parasite Plasmodium falciparum. J. Biol. Chem. 2012, 287, 37793–37807. [Google Scholar] [CrossRef] [Green Version]
- Iyer, J.K.; Shi, L.; Shankar, A.H.; Sullivan, D.J., Jr. Zinc protoporphyrin IX binds heme crystals to inhibit the process of crystallization in Plasmodium falciparum. Mol. Med. (Camb. Mass.) 2003, 9, 175–182. [Google Scholar] [CrossRef]
- Alves, E.; Iglesias, B.A.; Deda, D.K.; Budu, A.; Matias, T.A.; Bueno, V.B.; Maluf, F.V.; Guido, R.V.; Oliva, G.; Catalani, L.H.; et al. Encapsulation of metalloporphyrins improves their capacity to block the viability of the human malaria parasite Plasmodium falciparum. Nanomedicine 2015, 11, 351–358. [Google Scholar] [CrossRef]
- Ke, H.; Sigala, P.; Miura, K.; Morrisey, J.M.; Mather, M.; Crowley, J.R.; Henderson, J.P.; Goldberg, D.E.; Long, C.A.; Vaidya, A.B. The heme biosynthesis pathway is essential for Plasmodium falciparum development in mosquito stage but not in blood stages. J. Biol. Chem. 2014, 289, 34827–34837. [Google Scholar] [CrossRef] [Green Version]
- Chugh, M.; Sundararaman, V.; Kumar, S.; Reddy, V.S.; Siddiqui, W.A.; Stuart, K.D.; Malhotra, P. Protein complex directs hemoglobin-to-hemozoin formation in Plasmodium falciparum. Proc. Natl. Acad. Sci. USA 2013, 110, 5392–5397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, Y.; Zhang, Z.; Li, Q.; Li, W.; Xu, J.; Cao, H. Hyperbaric oxygen preconditioning protects lung against hyperoxic acute lung injury in rats via heme oxygenase-1 induction. Biochem. Biophys. Res. Commun. 2015, 456, 549–554. [Google Scholar] [CrossRef] [PubMed]
- Eckstein-Ludwig, U.; Webb, R.J.; Van Goethem, I.D.A.; East, J.M.; Lee, A.G.; Kimura, M.; O’Neill, P.M.; Bray, P.G.; Ward, S.; Krishna, S. Artemisinins target the SERCA of Plasmodium falciparum. Nature 2003, 424, 957–961. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Q.; Kyle, D.E.; Gatton, M.L. Artemisinin resistance in Plasmodium falciparum: A process linked to dormancy? Int. J. Parasitol. Drugs Drug. Resist. 2012, 2, 249–255. [Google Scholar] [CrossRef] [Green Version]
- Straimer, J.; Gnädig, N.F.; Witkowski, B.; Amaratunga, C.; Duru, V.; Ramadani, A.P.; Dacheux, M.; Khim, N.; Zhang, L.; Lam, S.; et al. Drug resistance. K13-propeller mutations confer artemisinin resistance in Plasmodium falciparum clinical isolates. Science 2015, 347, 428–431. [Google Scholar] [CrossRef] [Green Version]
- Kannan, R.; Kumar, K.; Sahal, D.; Kukreti, S.; Chauhan, V.S. Reaction of artemisinin with haemoglobin: Implications for antimalarial activity. Biochem. J. 2005, 385 Pt 2, 409–418. [Google Scholar] [CrossRef] [Green Version]
- Abshire, J.R.; Rowlands, C.J.; Ganesan, S.M.; So, P.T.; Niles, J.C. Quantification of labile heme in live malaria parasites using a genetically encoded biosensor. Proc. Natl. Acad. Sci. USA 2017, 114, E2068–E2076. [Google Scholar] [CrossRef] [Green Version]
- Heller, L.E.; Roepe, P.D. Quantification of Free Ferriprotoporphyrin IX Heme and Hemozoin for Artemisinin Sensitive versus Delayed Clearance Phenotype Plasmodium falciparum Malarial Parasites. Biochemistry 2018, 57, 6927–6934. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, C.-J.; Ni Chia, W.; Loh, C.C.Y.; Li, Z.; Lee, Y.M.; He, Y.; Yuan, L.-X.; Lim, T.K.; Liu, M.; et al. Haem-activated promiscuous targeting of artemisinin in Plasmodium falciparum. Nat. Commun. 2015, 6, 10111. [Google Scholar] [CrossRef]
- Harding, C.R.; Sidik, S.M.; Petrova, B.; Gnädig, N.F.; Okombo, J.; Herneisen, A.L.; Ward, K.E.; Markus, B.M.; Boydston, E.A.; Fidock, D.A.; et al. Genetic screens reveal a central role for heme metabolism in artemisinin susceptibility. Nat. Commun. 2020, 11, 4813. [Google Scholar] [CrossRef]
- Klonis, N.; Xie, S.; McCaw, J.; Crespo-Ortiz, M.P.; Zaloumis, S.; Simpson, J.A.; Tilley, L. Altered temporal response of malaria parasites determines differential sensitivity to artemisinin. Proc. Natl. Acad. Sci. USA 2013, 110, 5157–5162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atamna, H.; Brahmbhatt, M.; Atamna, W.; Shanower, G.A.; Dhahbi, J.M. ApoHRP-based assay to measure intracellular regulatory heme. Metallomics 2015, 7, 309–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanna, D.; Harvey, R.M.; Martinez-Guzman, O.; Yuan, X.; Chandrasekharan, B.; Raju, G.; Outten, F.; Hamza, I.; Reddi, A.R. Heme dynamics and trafficking factors revealed by genetically encoded fluorescent heme sensors. Proc. Natl. Acad. Sci. USA 2016, 113, 7539–7544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mercer, A.E.; Copple, I.M.; Maggs, J.L.; O’Neill, P.M.; Park, B.K. The role of heme and the mitochondrion in the chemical and molecular mechanisms of mammalian cell death induced by the artemisinin antimalarials. J. Biol. Chem. 2011, 286, 987–996. [Google Scholar] [CrossRef] [Green Version]
- Geroldinger, G.; Tonner, M.; Quirgst, J.; Walter, M.; De Sarkar, S.; Machín, L.; Monzote, L.; Stolze, K.; Duvigneau, J.C.; Staniek, K.; et al. Activation of artemisinin and heme degradation in Leishmania tarentolae promastigotes: A possible link. Biochem. Pharm. 2020, 173, 113737. [Google Scholar] [CrossRef]
- Sun, C.; Zhou, B. The molecular and cellular action properties of artemisinins: What has yeast told us? Microb. Cell 2016, 3, 196–205. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Huang, L.; Li, J.; Fan, Q.; Long, Y.; Li, Y.; Zhou, B. Artemisinin directly targets malarial mitochondria through its specific mitochondrial activation. PLoS ONE 2010, 5, e9582. [Google Scholar] [CrossRef] [Green Version]
- Antoine, T.; Fisher, N.; Amewu, R.; O’Neill, P.M.; Ward, S.; Biagini, G.A. Rapid kill of malaria parasites by artemisinin and semi-synthetic endoperoxides involves ROS-dependent depolarization of the membrane potential. J. Antimicrob. Chemother. 2014, 69, 1005–1016. [Google Scholar] [CrossRef]
- Combrinck, J.M.; Fong, K.Y.; Gibhard, L.; Smith, P.J.; Wright, D.W.; Egan, T.J. Optimization of a multi-well colorimetric assay to determine haem species in Plasmodium falciparum in the presence of anti-malarials. Malar. J. 2015, 14, 253. [Google Scholar] [CrossRef] [Green Version]
- Combrinck, J.M.; Mabotha, T.E.; Ncokazi, K.K.; Ambele, M.A.; Taylor, D.; Smith, P.J.; Hoppe, H.; Egan, T.J. Insights into the role of heme in the mechanism of action of antimalarials. ACS Chem. Biol. 2013, 8, 133–137. [Google Scholar] [CrossRef] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, P.; Zhou, B. The Antagonizing Role of Heme in the Antimalarial Function of Artemisinin: Elevating Intracellular Free Heme Negatively Impacts Artemisinin Activity in Plasmodium falciparum. Molecules 2022, 27, 1755. https://doi.org/10.3390/molecules27061755
Zhu P, Zhou B. The Antagonizing Role of Heme in the Antimalarial Function of Artemisinin: Elevating Intracellular Free Heme Negatively Impacts Artemisinin Activity in Plasmodium falciparum. Molecules. 2022; 27(6):1755. https://doi.org/10.3390/molecules27061755
Chicago/Turabian StyleZhu, Pan, and Bing Zhou. 2022. "The Antagonizing Role of Heme in the Antimalarial Function of Artemisinin: Elevating Intracellular Free Heme Negatively Impacts Artemisinin Activity in Plasmodium falciparum" Molecules 27, no. 6: 1755. https://doi.org/10.3390/molecules27061755
APA StyleZhu, P., & Zhou, B. (2022). The Antagonizing Role of Heme in the Antimalarial Function of Artemisinin: Elevating Intracellular Free Heme Negatively Impacts Artemisinin Activity in Plasmodium falciparum. Molecules, 27(6), 1755. https://doi.org/10.3390/molecules27061755