
Apigenin Isolated from Carduus crispus Protects against H2O2-Induced Oxidative Damage and Spermatogenic Expression Changes in GC-2spd Sperm Cells

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
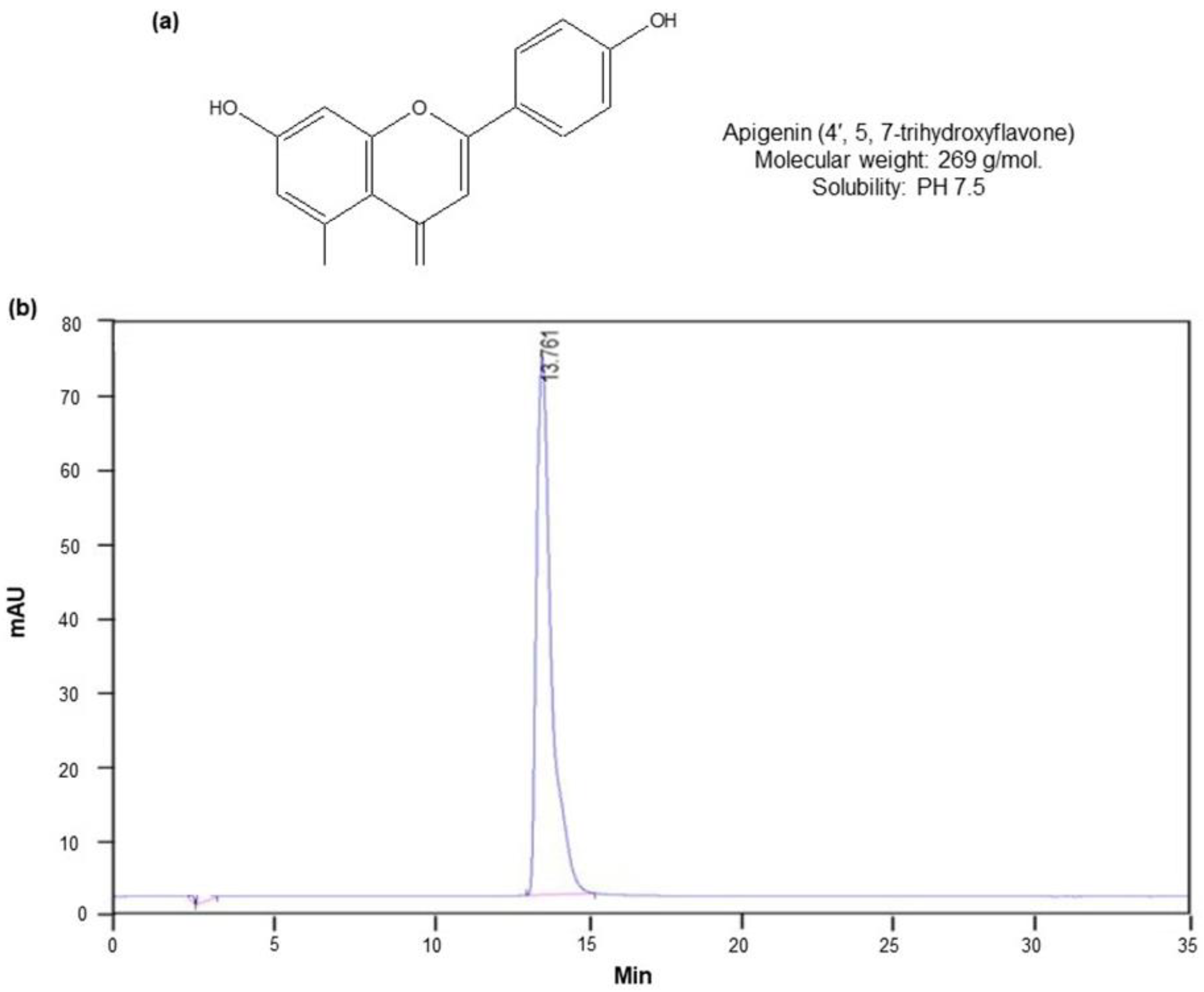
2.1. Isolation of Apigenin from C. crispus
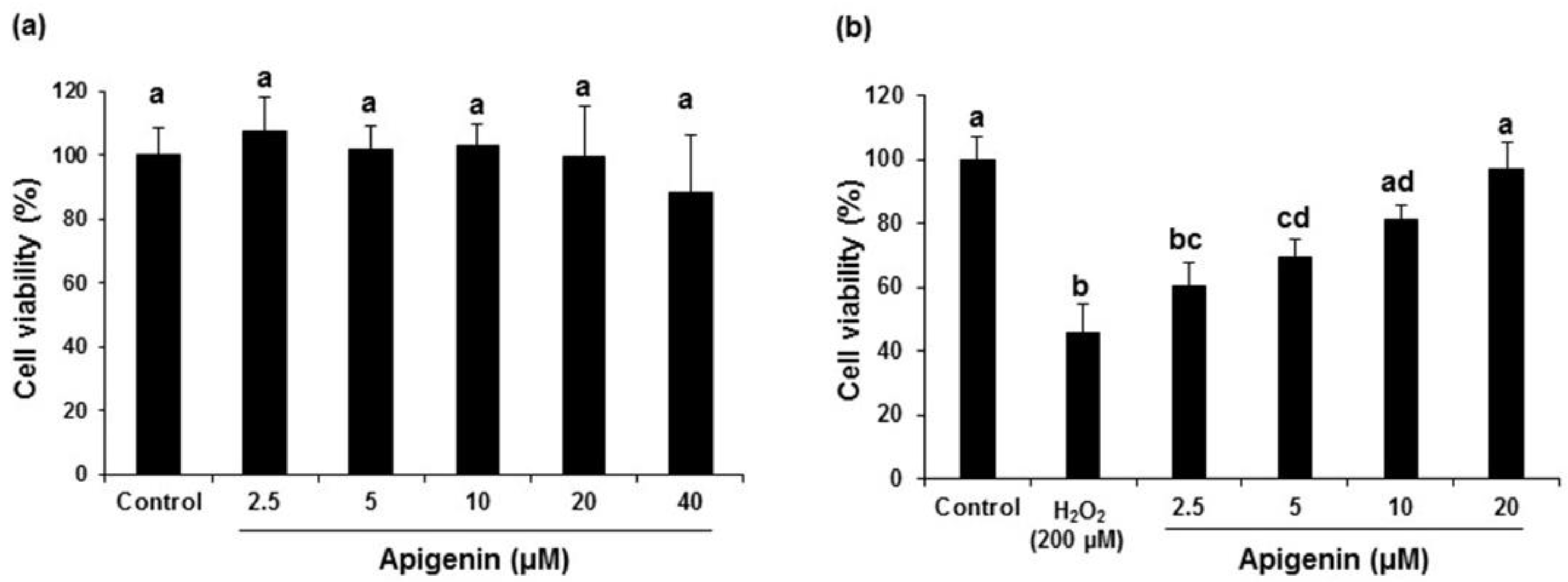
2.2. Effect of Apigenin on the Cell Viability in H2O2-Induced GC-2spd Cells
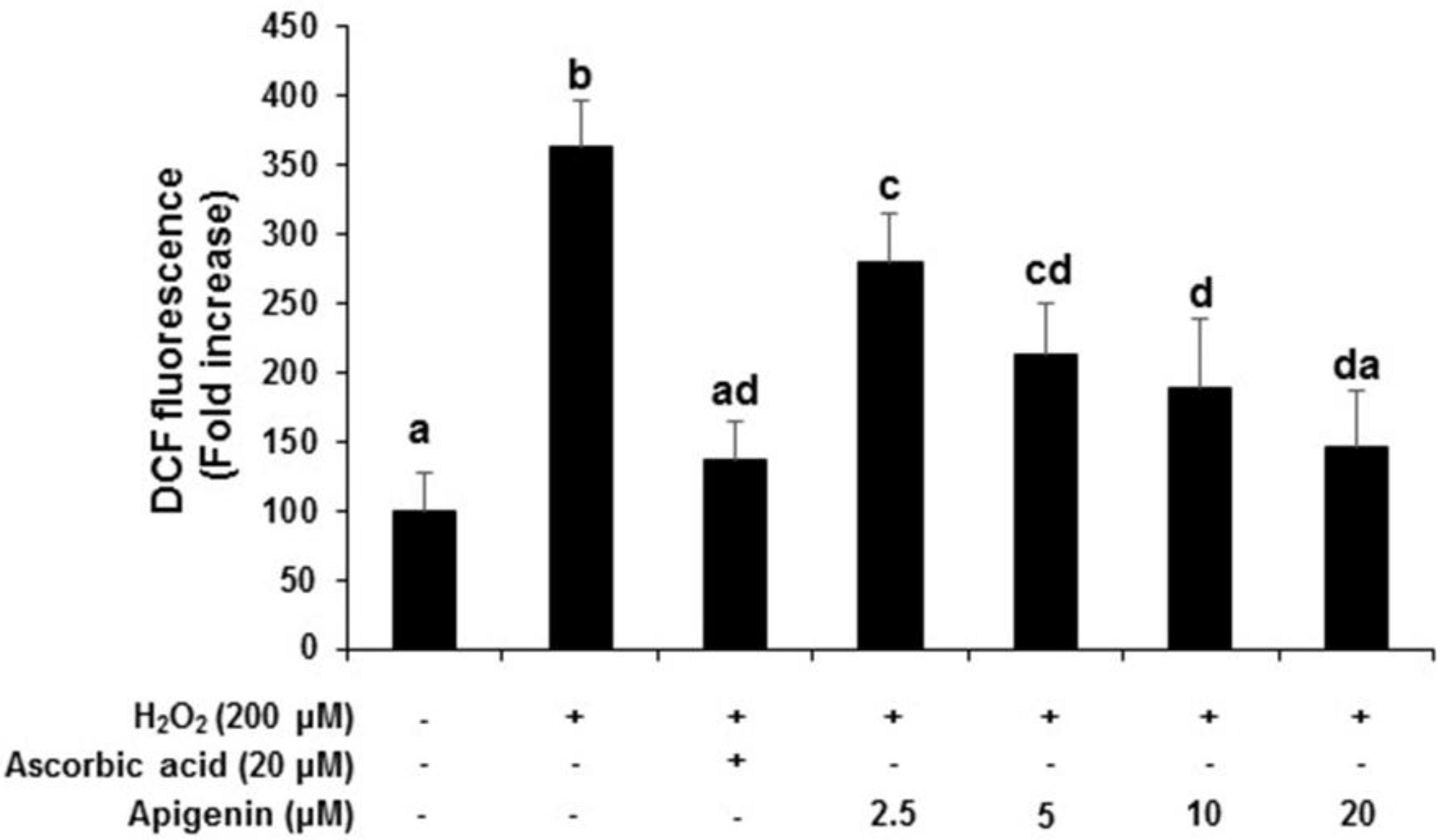
2.3. Effect of Apigenin on the Intracellular ROS Generation in H2O2-Induced GC-2spd Cells
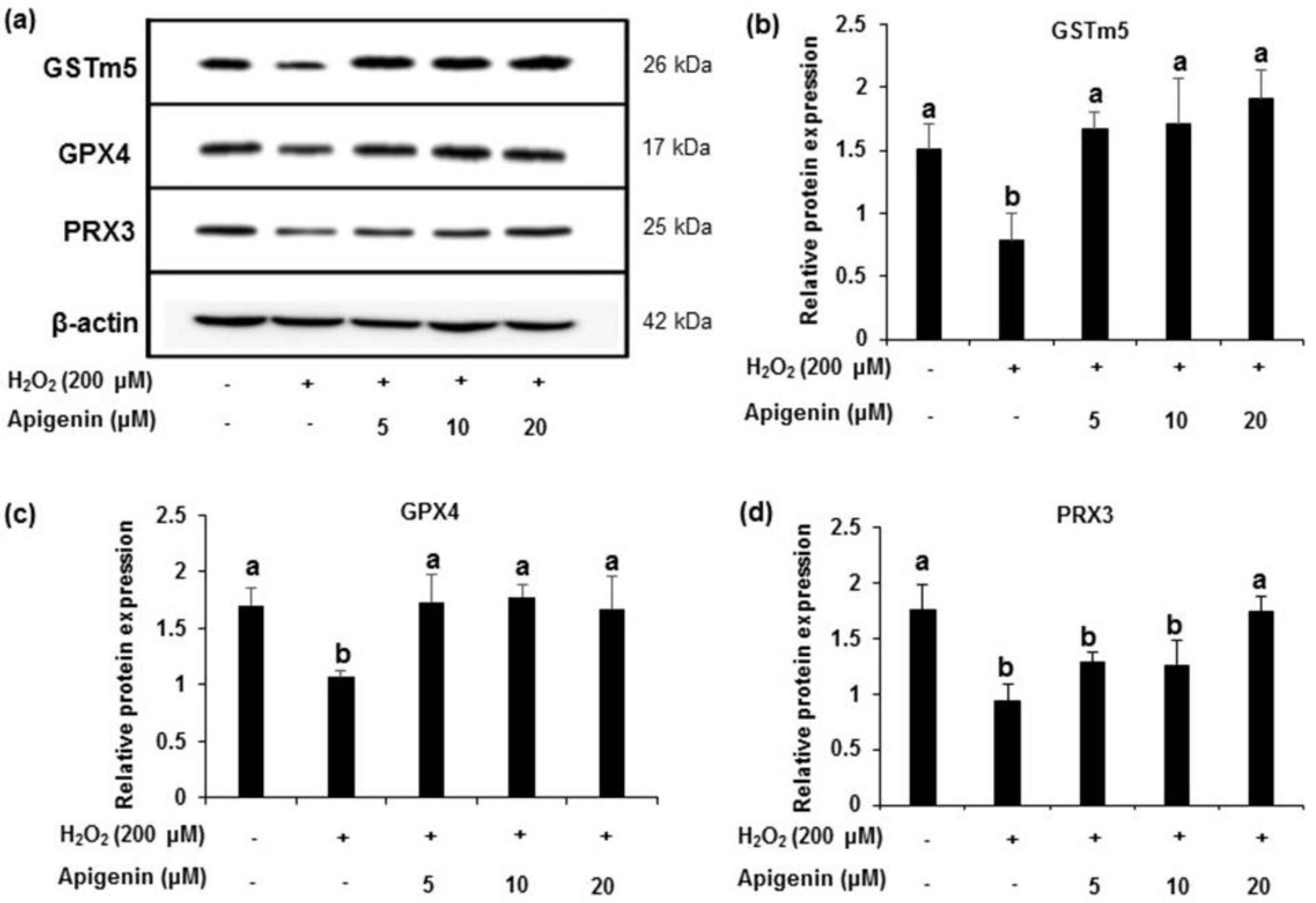
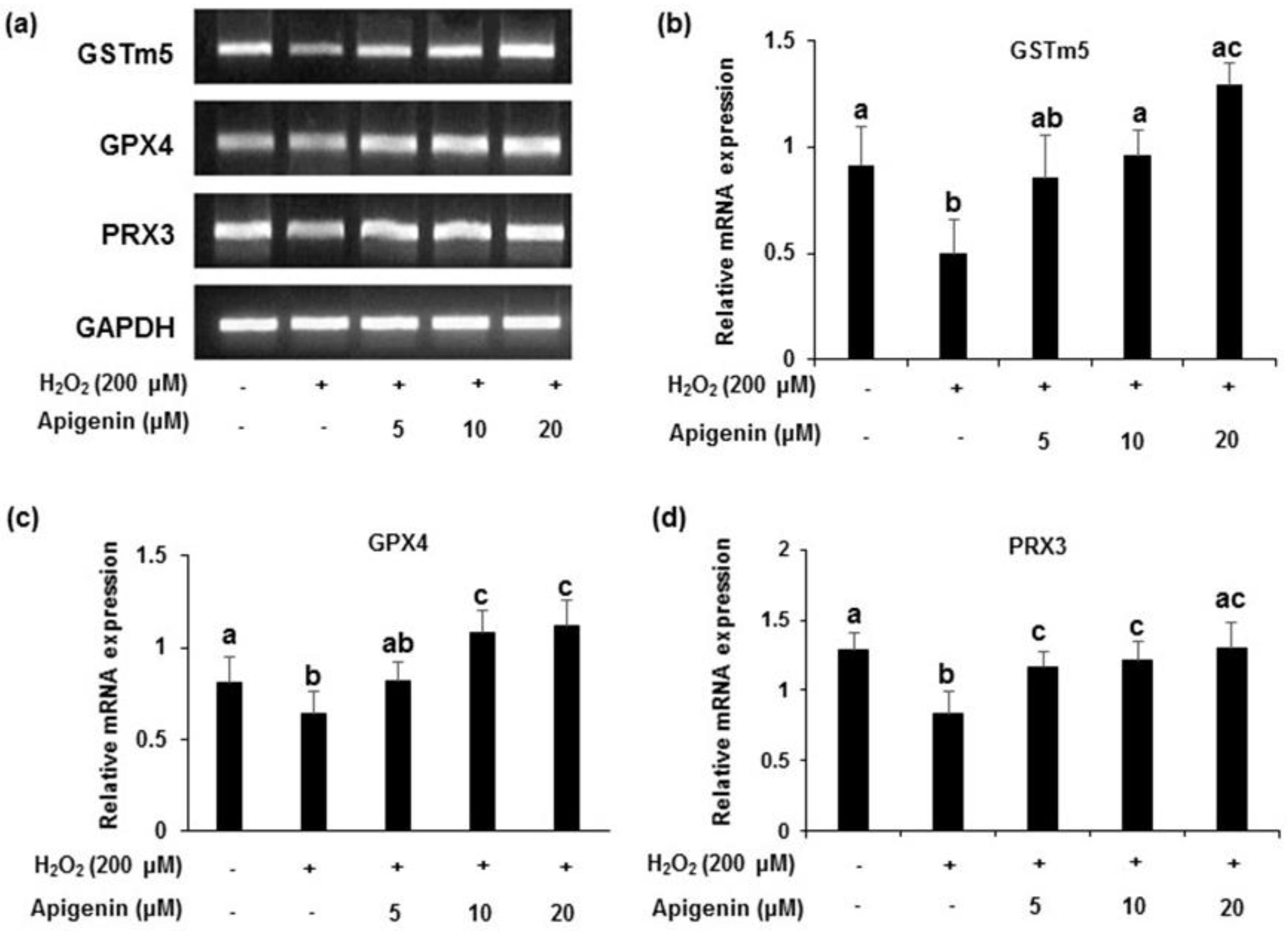
2.4. Effect of Apigenin on the Expression of Antioxidant Enzymes in H2O2-Induced GC-2spd Cells
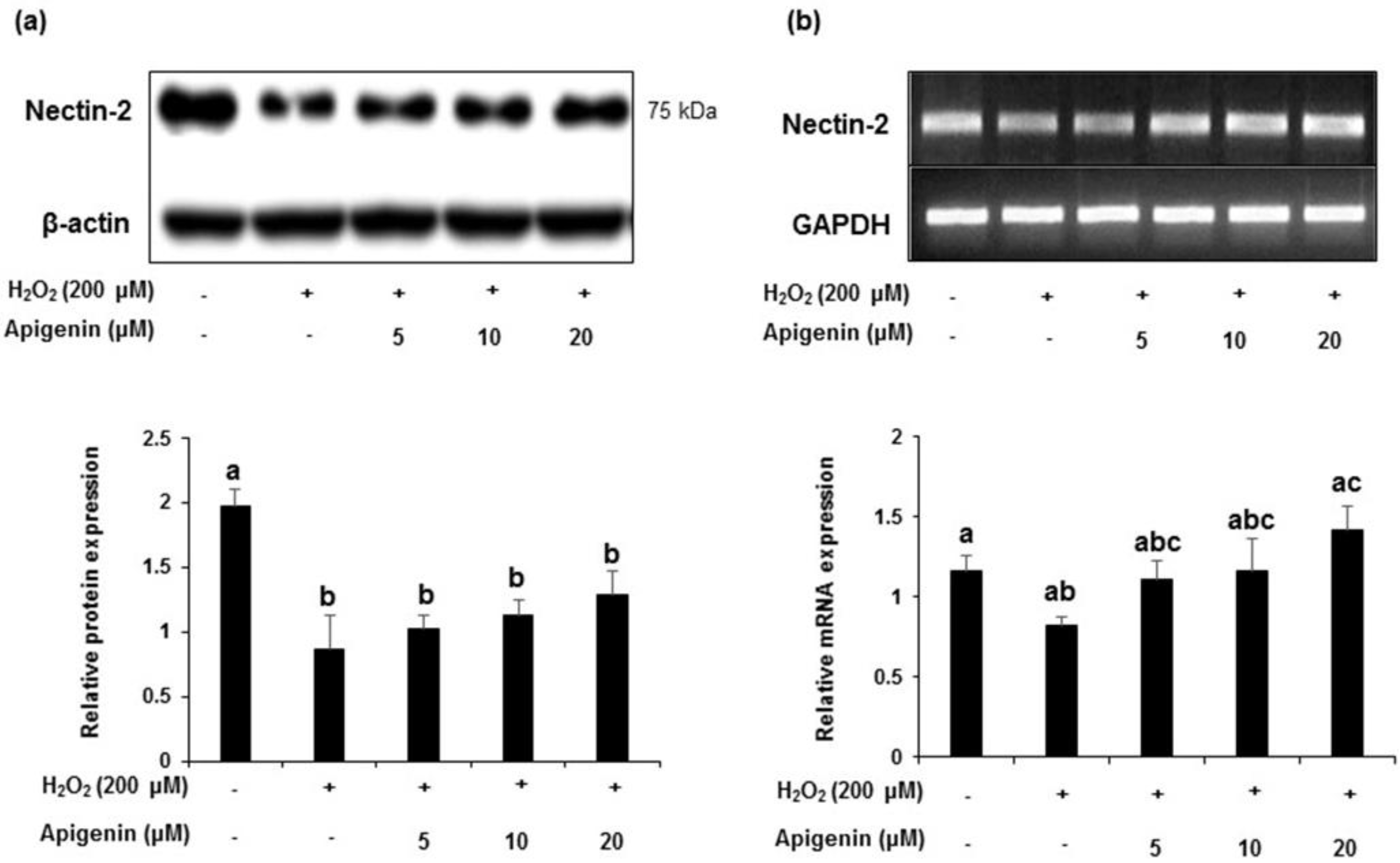
2.5. Effect of Apigenin on the Expression of Nectin-2 in H2O2-Exposed GC-2spd Cells
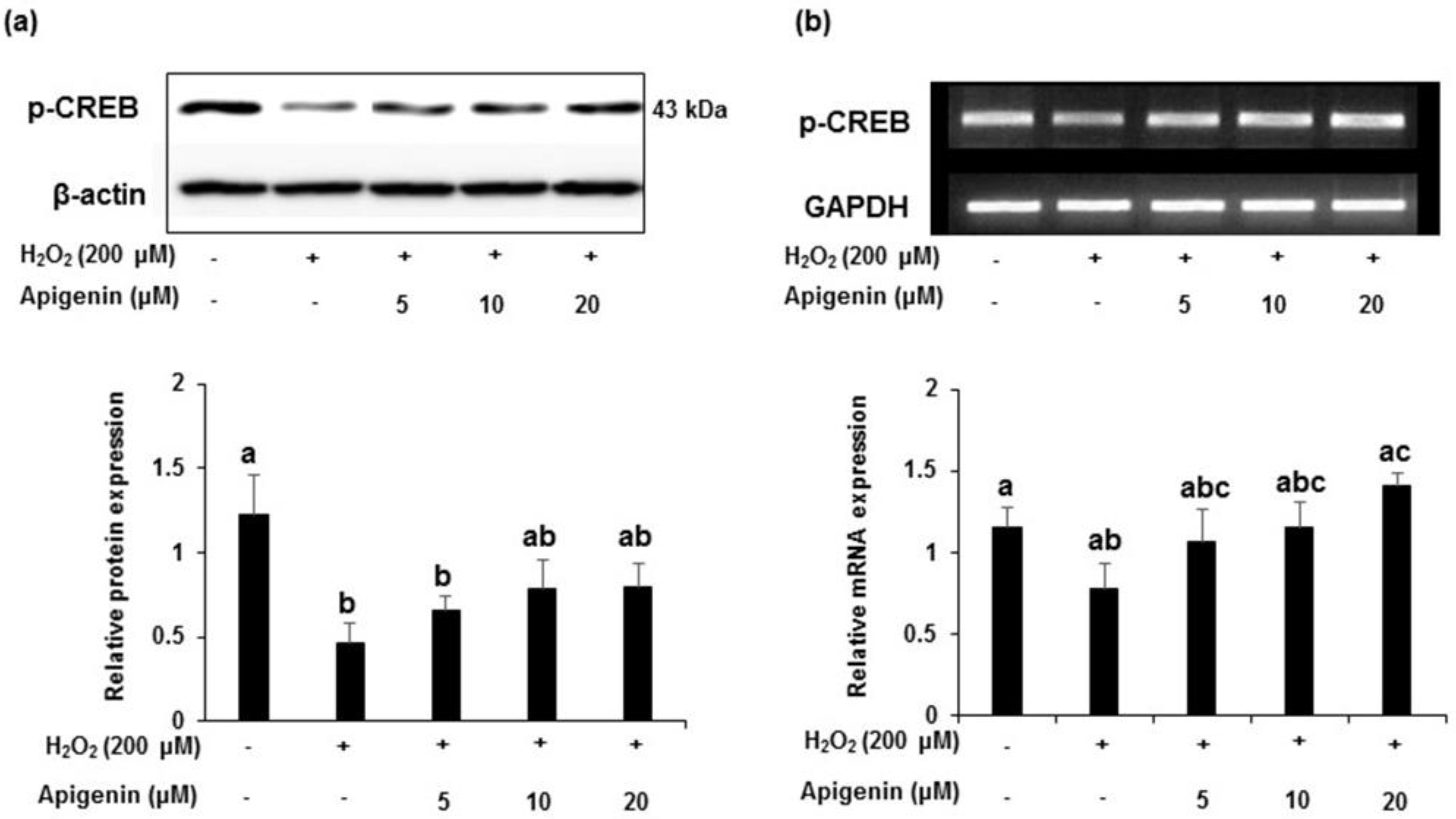
2.6. Effect of Apigenin on Expression of CREB in H2O2 Exposed GC-2spd Cells
3. Discussion
4. Materials and Methods
4.1. Chemicals and Medium for Cell Culture
4.2. Isolation and Purification of Apigenin from C. crispus
4.3. Cell Culture and Viability Assay
4.4. Intracellular Reactive Oxygen Species (ROS) Scavenging Assay
4.5. Reverse Transcription-Polymerase Chain Reaction (RT-PCR)
4.6. Western Blotting
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Vander Borght, M.; Wyns, C. Fertility and infertility: Definition and epidemiology. Clin. Biochem. 2018, 62, 2–10. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, A.; Baskaran, S.; Parekh, N.; Cho, C.-L.; Henkel, R.; Vij, S.; Arafa, M.; Panner Selvam, M.K.; Shah, R. Male infertility. Lancet 2021, 397, 319–333. [Google Scholar] [CrossRef]
- Chen, L.; Shi, G.; Huang, D.; Li, Y.; Ma, C.; Shi, M.; Su, B.; Shi, G. Male sexual dysfunction: A review of literature on its pathological mechanisms, potential risk factors, and herbal drug intervention. Biomed. Pharmacother. 2019, 112, 108585. [Google Scholar] [CrossRef] [PubMed]
- Dutta, S.; Sengupta, P.; Slama, P.; Roychoudhury, S. Oxidative Stress, Testicular Inflammatory Pathways, and Male Reproduction. Int. J. Mol. Sci. 2021, 22, 10043. [Google Scholar] [CrossRef]
- Fatima, S. Role of Reactive Oxygen Species in Male Reproduction. In Novel Prospects in Oxidative and Nitrosative Stress; InTechOpen: London, UK, 2018. [Google Scholar]
- Asadi, N. The Impact of Oxidative Stress on Testicular Function and the Role of Antioxidants in Improving it: A Review. J. Clin. Diagn. Res. 2017, 11, IE01. [Google Scholar] [CrossRef]
- Ramesh, T.; Yoo, S.-K.; Kim, S.-W.; Hwang, S.-Y.; Sohn, S.-H.; Kim, I.-W.; Kim, S.-K. Cordycepin (3′-deoxyadenosine) attenuates age-related oxidative stress and ameliorates antioxidant capacity in rats. Exp. Gerontol. 2012, 47, 979–987. [Google Scholar] [CrossRef]
- Urnukhsaikhan, E.; Bold, B.-E.; Gunbileg, A.; Sukhbaatar, N.; Mishig-Ochir, T. Antibacterial activity and characteristics of silver nanoparticles biosynthesized from Carduus crispus. Sci. Rep. 2021, 11, 21047. [Google Scholar] [CrossRef]
- Jeong, D.M.; Jung, H.A.; Choi, J.S. Comparative antioxidant activity and HPLC profiles of some selected Korean thistles. Arch. Pharm. Res. 2008, 31, 28–33. [Google Scholar] [CrossRef]
- Ali, F.; Rahul; Naz, F.; Jyoti, S.; Siddique, Y.H. Health functionality of apigenin: A review. Int. J. Food Prop. 2017, 20, 1197–1238. [Google Scholar] [CrossRef]
- Kim, Y.J.; Cho, E.J.; Lee, A.Y.; Seo, W.T. Apigenin Ameliorates Oxidative Stress-induced Neuronal Apoptosis in SH-SY5Y Cells. Microbiol. Biotechnol. Lett. 2021, 49, 138–147. [Google Scholar] [CrossRef]
- Skondras, I.; Lambropoulou, M.; Tsaroucha, A.; Gardikis, S.; Tripsianis, G.; Simopoulos, C.; Vaos, G. The role of Apigenin in testicular damage in experimental ischemia-reperfusion injury in rats. Hippokratia 2015, 19, 225–230. [Google Scholar] [PubMed]
- Ye, R.-J.; Yang, J.-M.; Hai, D.-M.; Liu, N.; Ma, L.; Lan, X.-B.; Niu, J.-G.; Zheng, P.; Yu, J.-Q. Interplay between male reproductive system dysfunction and the therapeutic effect of flavonoids. Fitoterapia 2020, 147, 104756. [Google Scholar] [CrossRef] [PubMed]
- Pei, Y.; Yang, L.; Wu, L.; He, H.; Geng, G.; Xu, D.; Chen, H.; Li, Q. Combined effect of apigenin and ferulic acid on frozen-thawed boar sperm quality. Anim. Sci. J. 2018, 89, 956–965. [Google Scholar] [CrossRef] [PubMed]
- Martin, L.J.; Touaibia, M. Improvement of Testicular Steroidogenesis Using Flavonoids and Isoflavonoids for Prevention of Late-Onset Male Hypogonadism. Antioxidants 2020, 9, 237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tabesh, M.; Azadbakht, L.; Faghihimani, E.; Tabesh, M.; Esmaillzadeh, A. Calcium-Vitamin D Cosupplementation Influences Circulating Inflammatory Biomarkers and Adipocytokines in Vitamin D-Insufficient Diabetics: A Randomized Controlled Clinical Trial. J. Clin. Endocrinol. Metab. 2014, 99, E2485–E2493. [Google Scholar] [CrossRef]
- Attimarad, M.; Mueen Ahmed, K.K.; Aldhubaib, B.E.; Harsha, S. High-performance thin layer chromatography: A powerful analytical technique in pharmaceutical drug discovery. Pharm. Methods 2011, 2, 71–75. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, N.Z.; Ibrahim, S.R.; Ahmed-Farid, O.A. Quercetin and Apigenin of Cymbopogon citratus mediate inhibition of HCT-116 and PC-3 cell cycle progression and ameliorate Doxorubicin-induced testicular dysfunction in male rats. Biomed. Res. Ther. 2018, 5, 2466–2479. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.Y.; Zhang, S.X.; Zhang, N.; Hao, C.F.; Zhuang, L.L.; Huang, X. Effects of apigenin on scrotal heat-induced damage in the mice testis. Int. J. Clin. Exp. Med. 2016, 9, 6342–6347. [Google Scholar]
- Dang, Y.; Li, Z.; Luo, B.; Pan, L.; Wei, Q.; Zhang, Y. Protective effects of apigenin against acrylonitrile-induced subchronic sperm injury in rats. Food Chem. Toxicol. 2017, 109, 517–525. [Google Scholar] [CrossRef]
- Rambaldi, A.; Jacobs, B.P.; Iaquinto, G.; Gluud, C. Milk Thistle for Alcoholic and/or Hepatitis B or C Liver Diseases-A Systematic Cochrane Hepato-Biliary Group Review with Meta-Analyses of Randomized Clinical Trials. Am. J. Gastroenterol. 2005, 100, 2583–2591. [Google Scholar] [CrossRef]
- Zhang, Q.; Tu, G.; Zhao, Y.; Cheng, T. Novel bioactive isoquinoline alkaloids from Carduus crispus. Tetrahedron 2002, 58, 6795–6798. [Google Scholar] [CrossRef]
- Greenlee, H.; Abascal, K.; Yarnell, E.; Ladas, E. Clinical Applications of Silybum marianum in Oncology. Integr. Cancer Ther. 2007, 6, 158–165. [Google Scholar] [CrossRef] [PubMed]
- Auten, R.L.; Davis, J.M. Oxygen Toxicity and Reactive Oxygen Species: The Devil Is in the Details. Pediatr. Res. 2009, 66, 121–127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanocka, D.; Kurpisz, M. Reactive oxygen species and sperm cells. Reprod. Biol. Endocrinol. 2004, 2, 12. [Google Scholar] [CrossRef] [Green Version]
- Agarwal, A.; Virk, G.; Ong, C.; du Plessis, S.S. Effect of oxidative stress on male reproduction. World J. Mens. Health 2014, 32, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Won, Y.-J.; Kim, B.; Shin, Y.-K.; Jung, S.-H.; Yoo, S.-K.; Hwang, S.-Y.; Sung, J.-H.; Kim, S.-K. Pectinase-treated Panax ginseng extract (GINST) rescues testicular dysfunction in aged rats via redox-modulating proteins. Exp. Gerontol. 2014, 53, 57–66. [Google Scholar] [CrossRef]
- Kopalli, S.R.; Cha, K.-M.; Lee, S.-H.; Hwang, S.-Y.; Lee, Y.-J.; Koppula, S.; Kim, S.-K. Cordycepin, an Active Constituent of Nutrient Powerhouse and Potential Medicinal Mushroom Cordyceps militaris Linn., Ameliorates Age-Related Testicular Dysfunction in Rats. Nutrients 2019, 11, 906. [Google Scholar] [CrossRef] [Green Version]
- Mandai, K.; Rikitake, Y.; Mori, M.; Takai, Y. Nectins and Nectin-Like Molecules in Development and Disease. Curr. Top. Dev. Biol. 2015, 112, 197–231. [Google Scholar]
- Allocati, N.; Masulli, M.; Di Ilio, C.; Federici, L. Glutathione transferases: Substrates, inihibitors and pro-drugs in cancer and neurodegenerative diseases. Oncogenesis 2018, 7, 8. [Google Scholar] [CrossRef]
- Laborde, E. Glutathione transferases as mediators of signaling pathways involved in cell proliferation and cell death. Cell Death Differ. 2010, 17, 1373–1380. [Google Scholar] [CrossRef] [Green Version]
- Hayes, J.D.; Pulford, D.J. The Glut athione S-Transferase Supergene Family: Regulation of GST and the Contribution of the lsoenzymes to Cancer Chemoprotection and Drug Resistance Part I. Crit. Rev. Biochem. Mol. Biol. 1995, 30, 445–520. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.K.; Cha, K.-M.; Hwang, S.-Y.; Park, U.-K.; Seo, S.K.; Lee, S.-H.; Jeong, M.-S.; Cho, S.; Kopalli, S.R.; Kim, S.-K. Pectinase-treated Panax ginseng protects heat stress-induced testicular damage in rats. Reproduction 2017, 153, 737–747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, C.; Sun, S.; Johnson, T.; Qi, R.; Zhang, S.; Zhang, J.; Yang, K. The glutathione peroxidase Gpx4 prevents lipid peroxidation and ferroptosis to sustain Treg cell activation and suppression of antitumor immunity. Cell Rep. 2021, 35, 109235. [Google Scholar] [CrossRef] [PubMed]
- Maiorino, M.; Conrad, M.; Ursini, F. GPx4, Lipid Peroxidation, and Cell Death: Discoveries, Rediscoveries, and Open Issues. Antioxid. Redox Signal. 2018, 29, 61–74. [Google Scholar] [CrossRef]
- Fujii, J.; Imai, H. Redox reactions in mammalian spermatogenesis and the potential targets of reactive oxygen species under oxidative stress. Spermatogenesis 2014, 4, e979108. [Google Scholar] [CrossRef] [Green Version]
- Imai, H.; Hakkaku, N.; Iwamoto, R.; Suzuki, J.; Suzuki, T.; Tajima, Y.; Konishi, K.; Minami, S.; Ichinose, S.; Ishizaka, K.; et al. Depletion of Selenoprotein GPx4 in Spermatocytes Causes Male Infertility in Mice. J. Biol. Chem. 2009, 284, 32522–32532. [Google Scholar] [CrossRef] [Green Version]
- Wood, Z.A.; Poole, L.B.; Karplus, P.A. Peroxiredoxin Evolution and the Regulation of Hydrogen Peroxide Signaling. Science 2003, 300, 650–653. [Google Scholar] [CrossRef]
- Duan, T.; Fan, K.; Chen, S.; Yao, Q.; Zeng, R.; Hong, Z.; Peng, L.; Shao, Y.; Yao, B. Role of peroxiredoxin 2 in H2O2-induced oxidative stress of primary Leydig cells. Mol. Med. Rep. 2016, 13, 4807–4813. [Google Scholar] [CrossRef] [Green Version]
- Dyson, M.T.; Kowalewski, M.P.; Manna, P.R.; Stocco, D.M. The differential regulation of steroidogenic acute regulatory protein-mediated steroidogenesis by type I and type II PKA in MA-10 cells. Mol. Cell. Endocrinol. 2009, 300, 94–103. [Google Scholar] [CrossRef] [Green Version]
- Reyland, M.E.; Evans, R.M.; White, E.K. Lipoproteins Regulate Expression of the Steroidogenic Acute Regulatory Protein (StAR) in Mouse Adrenocortical Cells. J. Biol. Chem. 2000, 275, 36637–36644. [Google Scholar] [CrossRef] [Green Version]
- Sandhoff, T.W.; Hales, D.B.; Hales, K.H.; McLean, M.P. Transcriptional Regulation of the Rat Steroidogenic Acute Regulatory Protein Gene by Steroidogenic Factor 1**This work was supported by NIH Grant R01-HD35163 (to M.P.M.). Endocrinology 1998, 139, 4820–4831. [Google Scholar] [CrossRef] [PubMed]
- De Cesare, D.; Sassone-Corsi, P. Transcriptional regulation by cyclic AMP-responsive factors. Prog. Nucleic Acid. Res Mol. Biol. 2000, 64, 343–369. [Google Scholar] [PubMed]
- Shaywitz, A.J.; Greenberg, M.E. CREB: A Stimulus-Induced Transcription Factor Activated by A Diverse Array of Extracellular Signals. Annu. Rev. Biochem. 1999, 68, 821–861. [Google Scholar] [CrossRef] [PubMed]
- Naqvi, S.; Martin, K.J.; Arthur, J.S.C. CREB phosphorylation at Ser133 regulates transcription via distinct mechanisms downstream of cAMP and MAPK signalling. Biochem. J. 2014, 458, 469–479. [Google Scholar] [CrossRef]
- Zhang, X.; Lui, W.-Y. Dysregulation of nectin-2 in the testicular cells: An explanation of cadmium-induced male infertility. Biochim. Biophys. Acta-Gene Regul. Mech. 2014, 1839, 873–884. [Google Scholar] [CrossRef]
- Mueller, S. Loss of Nectin-2 at Sertoli-Spermatid Junctions Leads to Male Infertility and Correlates with Severe Spermatozoan Head and Midpiece Malformation, Impaired Binding to the Zona Pellucida, and Oocyte Penetration. Biol. Reprod. 2003, 69, 1330–1340. [Google Scholar] [CrossRef]
- Lui, W.-Y.; Sze, K.-L.; Lee, W.M. Nectin-2 expression in testicular cells is controlled via the functional cooperation between transcription factors of the Sp1, CREB, and AP-1 families. J. Cell. Physiol. 2006, 207, 144–157. [Google Scholar] [CrossRef] [Green Version]
- Ryu, H.; Lee, J.; Impey, S.; Ratan, R.R.; Ferrante, R.J. Antioxidants modulate mitochondrial PKA and increase CREB binding to D-loop DNA of the mitochondrial genome in neurons. Proc. Natl. Acad. Sci. USA 2005, 102, 13915–13920. [Google Scholar] [CrossRef] [Green Version]
- Kumashiro, N.; Tamura, Y.; Uchida, T.; Ogihara, T.; Fujitani, Y.; Hirose, T.; Mochizuki, H.; Kawamori, R.; Watada, H. Impact of Oxidative Stress and Peroxisome Proliferator-Activated Receptor Coactivator-1 in Hepatic Insulin Resistance. Diabetes 2008, 57, 2083–2091. [Google Scholar] [CrossRef] [Green Version]
- Kielkopf, C.L.; Bauer, W.; Urbatsch, I.L. Bradford Assay for Determining Protein Concentration. Cold Spring Harb. Protoc. 2020, 4, 102269. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kopalli, S.R.; Yoo, S.-K.; Kim, B.; Kim, S.-K.; Koppula, S. Apigenin Isolated from Carduus crispus Protects against H2O2-Induced Oxidative Damage and Spermatogenic Expression Changes in GC-2spd Sperm Cells. Molecules 2022, 27, 1777. https://doi.org/10.3390/molecules27061777
Kopalli SR, Yoo S-K, Kim B, Kim S-K, Koppula S. Apigenin Isolated from Carduus crispus Protects against H2O2-Induced Oxidative Damage and Spermatogenic Expression Changes in GC-2spd Sperm Cells. Molecules. 2022; 27(6):1777. https://doi.org/10.3390/molecules27061777
Chicago/Turabian StyleKopalli, Spandana Rajendra, Sung-Kwang Yoo, Bokyung Kim, Si-Kwan Kim, and Sushruta Koppula. 2022. "Apigenin Isolated from Carduus crispus Protects against H2O2-Induced Oxidative Damage and Spermatogenic Expression Changes in GC-2spd Sperm Cells" Molecules 27, no. 6: 1777. https://doi.org/10.3390/molecules27061777
APA StyleKopalli, S. R., Yoo, S. -K., Kim, B., Kim, S. -K., & Koppula, S. (2022). Apigenin Isolated from Carduus crispus Protects against H2O2-Induced Oxidative Damage and Spermatogenic Expression Changes in GC-2spd Sperm Cells. Molecules, 27(6), 1777. https://doi.org/10.3390/molecules27061777