
Influence of Soil Salinity on Selected Element Contents in Different Brassica Species

 ,
, 
Abstract
:1. Introduction
2. Results and Discussion
2.1. Plant Growth and Salinity Stress
2.2. Figures of Merit of the Analytical Procedure
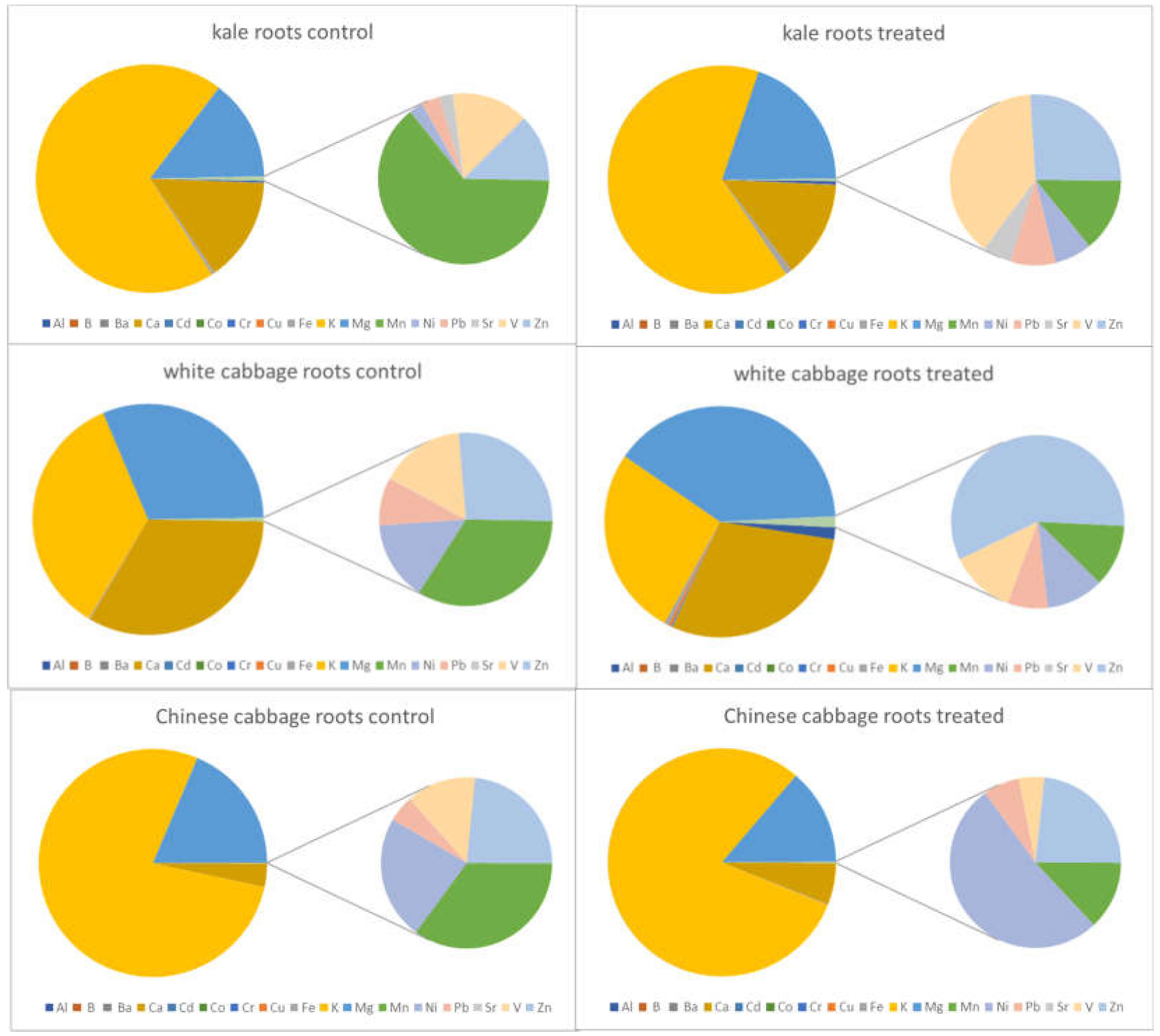
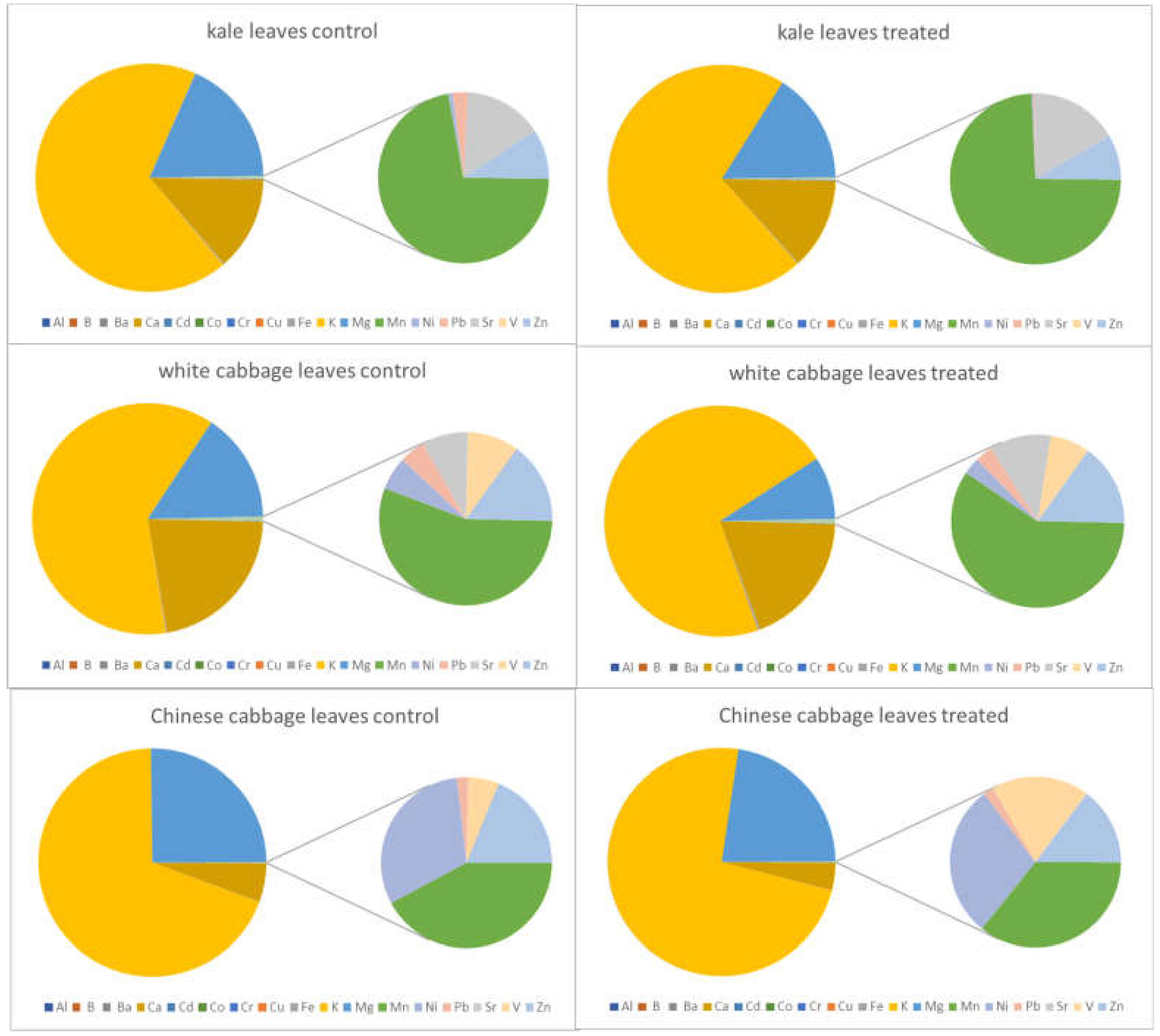
2.3. Elemental Contents in Salt-Treated Roots and Leaves Compared with the Corresponding Control
2.3.1. General Findings
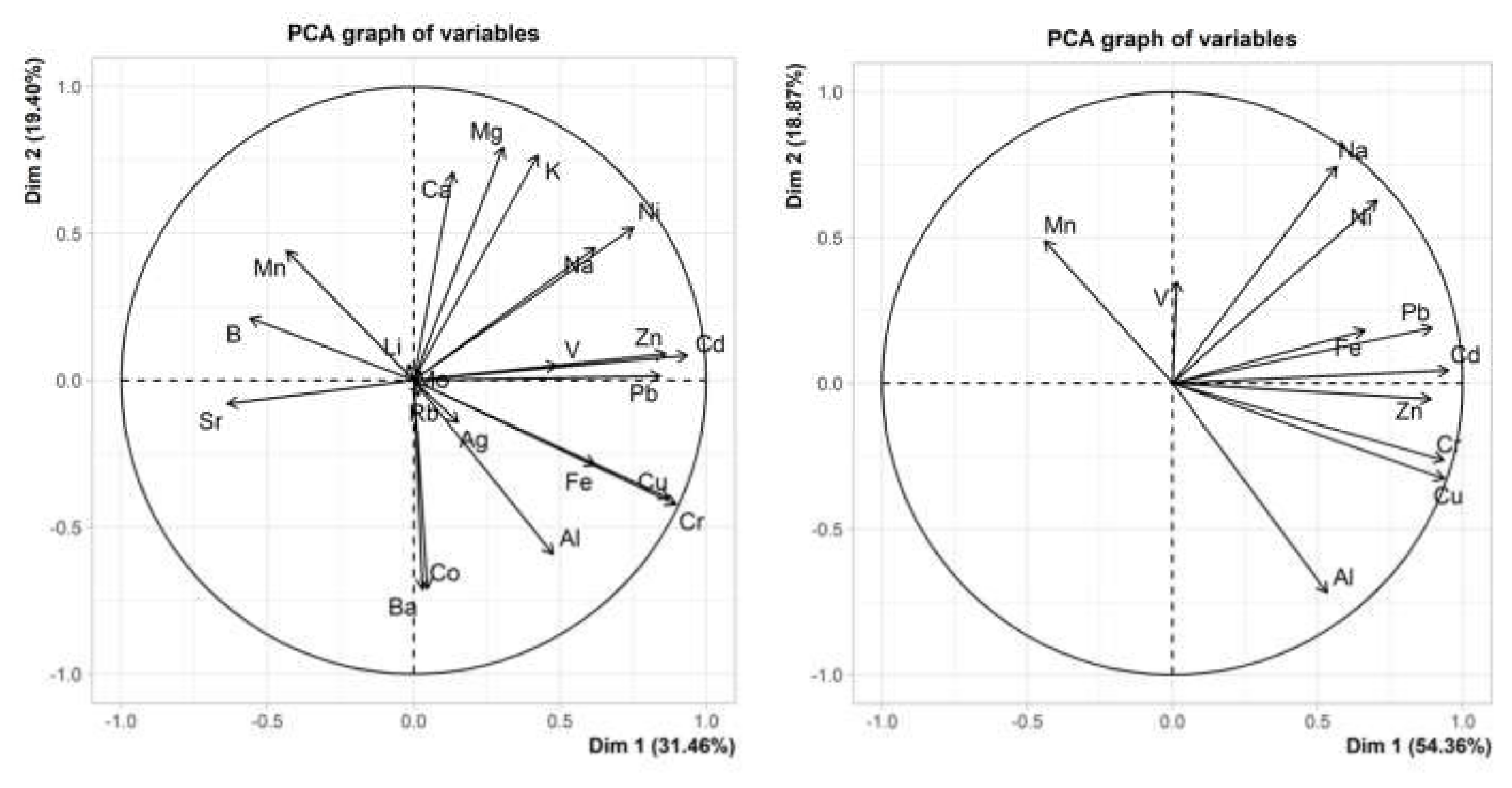
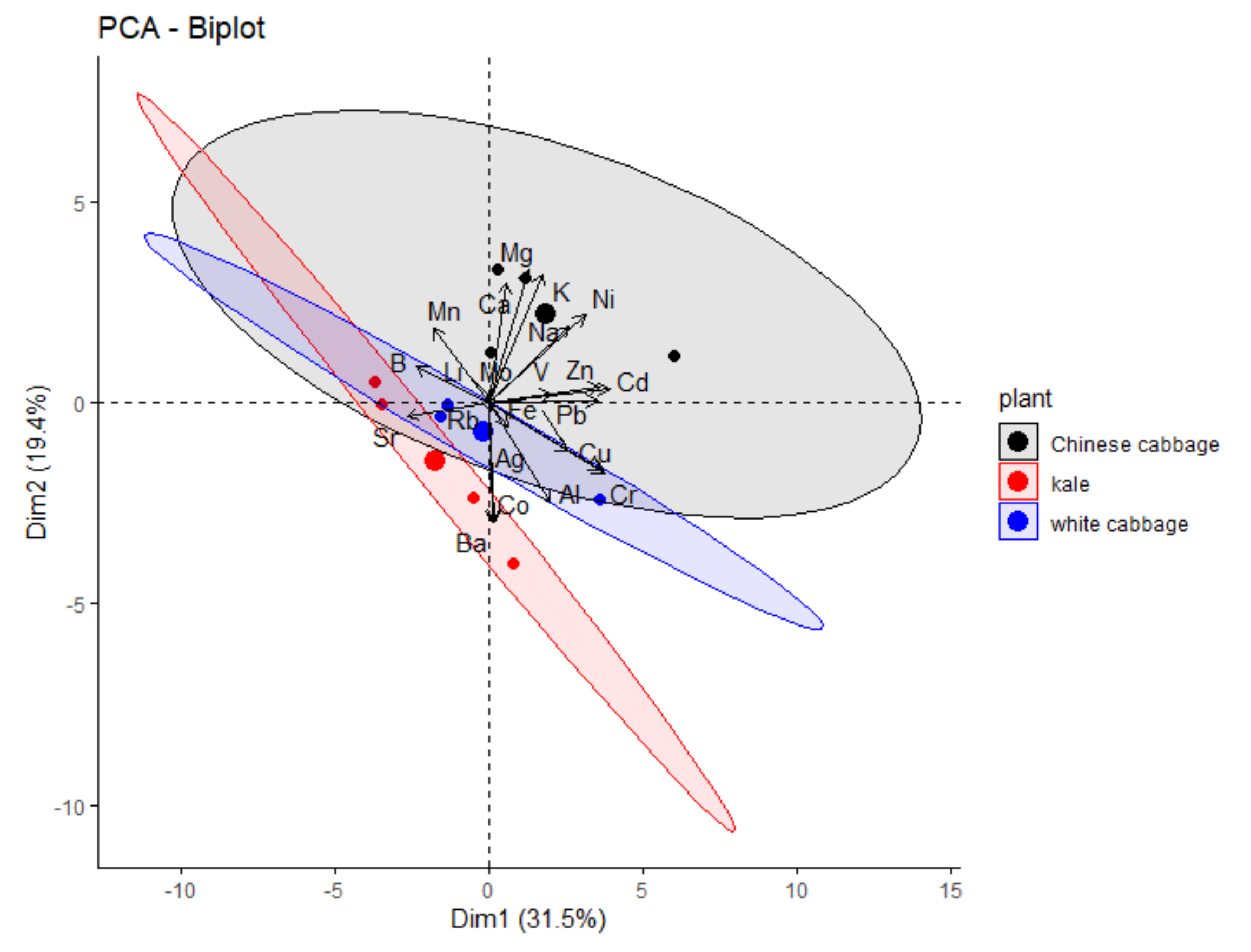
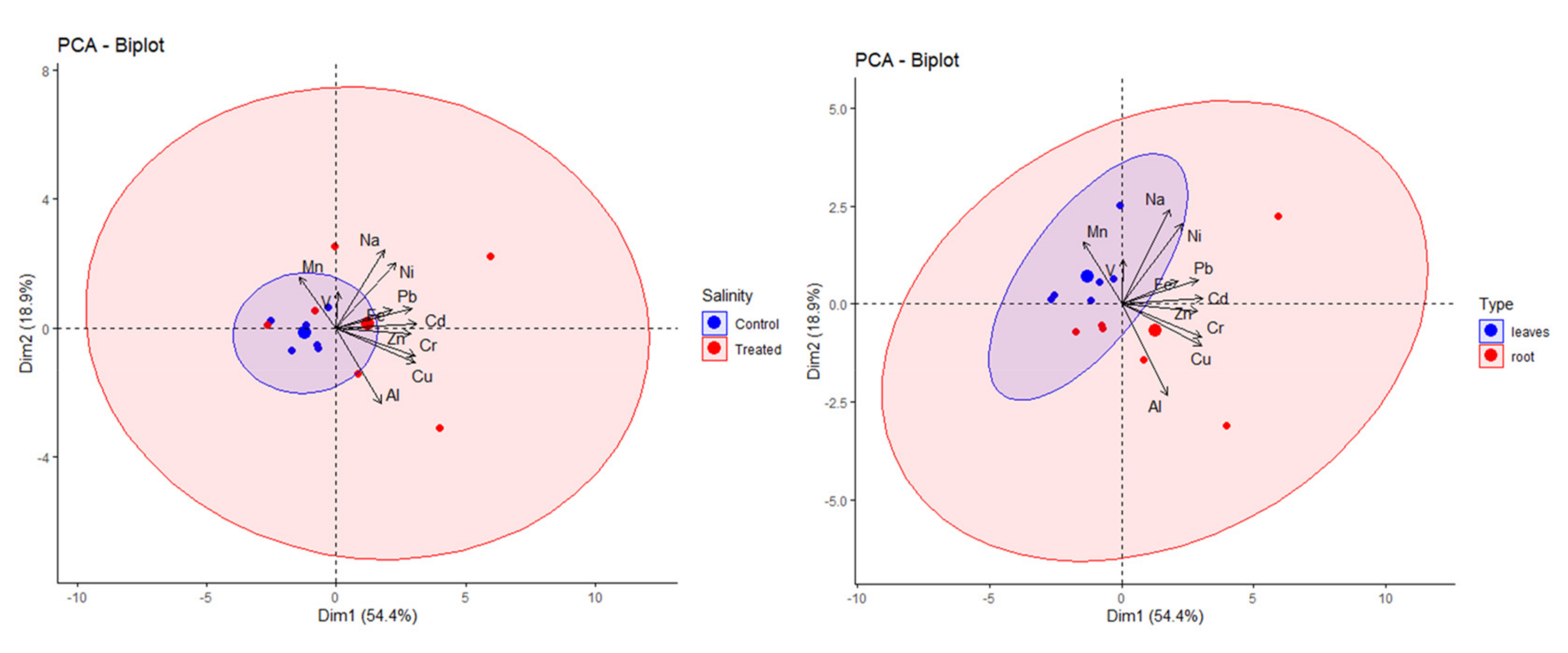
2.3.2. Principal Component Analysis of Obtained Data
3. Materials and Methods
3.1. Growing Plants and Sampling
3.2. Sample Preparation and Elemental Analysis
3.3. Data Evaluation and Statistical Tests
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Purty, R.S.; Kumar, G.; Singla-Pareek, S.L.; Pareek, A. Towards salinity tolerance in Brassica: An overview. Physiol. Mol. Biol. Plants 2008, 14, 39–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shrivastava, P.; Kumar, R. Soil salinity: A serious environmental issue and plant growth promoting bacteria as one of the tools for its alleviation. Saudi J. Biol. Sci. 2015, 22, 123–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jamil, A.; Riaz, S.; Ashraf, M.; Foolad, M.R. Gene expression profiling of plants under salt stress. Crit. Rev. Plant Sci. 2011, 30, 435–458. [Google Scholar] [CrossRef]
- Munns, R.; Gilliham, M. Salinity tolerance of crops—What is the cost? New Phytol. 2015, 208, 668–673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdul Qados, A.M.S. Effect of salt stress on plant growth and metabolism of bean plant Vicia faba (L.). J. Saudi Soc. Agric. Sci. 2011, 10, 7–15. [Google Scholar] [CrossRef] [Green Version]
- Mittal, S.; Kumari, N.; Sharma, V. Differential response of salt stress on Brassica juncea: Photosynthetic performance, pigment, proline, D1 and antioxidant enzymes. Plant Physiol. Biochem. 2012, 54, 17–26. [Google Scholar] [CrossRef]
- Julkowska, M.M.; Testerink, C. Tuning plant signaling and growth to survive salt. Trends Plant Sci. 2015, 20, 586–594. [Google Scholar] [CrossRef] [Green Version]
- Šamec, D.; Salopek-Sondi, B. Cruciferous (Brassicaceae) vegetables. In Nonvitamin and Nonmineral Nutritional Supplements; Nabavi, S.M., Sanches Silva, T., Eds.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 195–202. [Google Scholar] [CrossRef]
- Fahey, J.W. Brassica: Characteristics and Properties. In Encyclopedia of Food and Health, Volume 1—Technology & Engineering; Caballero, B., Finglas, P.M., Toldra, F., Eds.; Academic Press: Cambridge, MA, USA; Elsevier: Oxford, UK, 2016; pp. 469–477. [Google Scholar]
- Hussain, S.; Hussain, S.; Ali, B.; Ren, X.; Chen, X.; Li, Q.; Saqib, M.; Ahmad, N. Recent progress in understanding salinity tolerance in plants: Story of Na+/K+ balance and beyond. Plant Physiol. Biochem. 2021, 160, 239–256. [Google Scholar] [CrossRef]
- Steinnes, E. Soil and human health. In Sustaining Soil Productivity in Response to Global Climate Change: Science, Policy, and Ethics, 1st ed.; Sauer, T.J., Norman, J.M., Sivakumar, M.V.K., Eds.; Chapter 6; Wiley-Blackwell: Oxford, UK, 2011; pp. 79–86. [Google Scholar]
- Soetan, K.O.; Olaiya, C.O.; Oyewole, O.E. The importance of mineral elements for humans, domestic animals and plants: A review. Afr. J. Food Sci. 2010, 4, 200–222. [Google Scholar]
- Serbula, S.M.; Kalinovic, T.S.; Ilic, A.A.; Kalinovic, J.V.; Steharnik, M.M. Assessment of airborne heavy metal pollution using Pinus spp. and Tilia spp. Aerosol Air Qual. Res. 2013, 13, 563–573. [Google Scholar] [CrossRef] [Green Version]
- Jurić, D.; Puntarić, D.; Gvozdić, V.; Vidosavljević, D.; Lončarić, Z.; Puntarić, A.; Puntarić, E.; Puntarić, I.; Vidosavljević, M.; Begović, L.; et al. Cabbage (Brassica oleracea var. capitata) as possible indicator of wartime metal and metalloid contamination in eastern Croatia (ICP-MS method). Acta Agric. Scand. B Soil Plant Sci. 2017, 67, 270–277. [Google Scholar]
- Zeiner, M.; Juranović Cindrić, I.; Mihajlov Konanov, D.; Stingeder, G. Determination of selected toxic elements in leaves of White Hawthorn grown in a remote area. In Proceedings of the 16th International Conference on Heavy Metals in the Environment, Rome, Italy, 23–27 September 2012; pp. 34003–34006. Available online: https://www.e3s-conferences.org/articles/e3sconf/abs/2013/01/e3sconf_ichm13_34003/e3sconf_ichm13_34003.html (accessed on 9 March 2022).
- Pavlović, I.; Mlinarić, S.; Tarkowská, D.; Oklestková, J.; Novák, O.; Lepeduš, H.; Vujčić Bok, V.; Radić Brkanac, S.; Strnad, M.; Salopek-Sondi, B. Early Brassica crops responses to salinity stress: A comparative analysis between Chinese cabbage, white cabbage and kale. Front. Plant Sci. 2019, 10, 450. [Google Scholar] [CrossRef] [Green Version]
- Pavlović, I.; Pěnčík, A.; Novák, O.; Vujčić, V.; Radić Brkanac, S.; Lepeduš, H.; Strnad, M.; Salopek Sondi, B. Short-term salt stress in Brassica rapa seedlings causes alterations in auxin metabolism. Plant Physiol. Biochem. 2018, 125, 74–84. [Google Scholar] [CrossRef]
- An, J.I.; Lee, S.; Oh, M.M. Air anions promote the growth and mineral accumulation of spinach (Spinacia oleracea) cultivated in greenhouses. Hortic. Sci. Technol. 2021, 39, 332–342. [Google Scholar]
- Lee, S.; Oh, M.M. Electric stimulation promotes growth, mineral uptake, and antioxidant accumulation in kale (Brassica oleracea var. acephala). Bioelectrochemistry 2021, 138, 107727. [Google Scholar] [CrossRef]
- Mohamed, A.E.; Rashed, M.N.; Mofty, A. Assessment of essential and toxic elements in some kinds of vegetables. Ecotoxicol. Environ. Saf. 2003, 55, 251–260. [Google Scholar] [CrossRef]
- Miller-Cebert, R.L.; Sistani, N.A.; Cebert, E. Comparative mineral composition among canola cultivars and other cruciferous leafy greens. J. Food Compos. Anal. 2009, 22, 112–116. [Google Scholar] [CrossRef]
- Thavarajah, D.; Thavarajah, P.; Abare, A.; Basnagala, S.; Lacher, C.; Smith, P.; Combs, G.F. Mineral micronutrient and prebiotic carbohydrate profiles of USA-grown kale (Brassica oleracea L. var. acephala). J. Food Compos. Anal. 2016, 52, 9–15. [Google Scholar] [CrossRef] [Green Version]
- Wubishet Gezahegn, W.; Srinivasulu, A.; Aruna, B.; Banerjee, S.; Sudarshan, M.; Lakshmi Narayana, P.V.; Rao, A.D.P. Study of heavy metals accumulation in leafy vegetables of Ethiopia. IOSR J. Environ. Sci. Toxicol. Food Technol. 2017, 11, 57–68. [Google Scholar] [CrossRef]
- Korus, A. Changes in the content of minerals, B-group vitamins and tocopherols in processed kale leaves. J. Food Compos. Anal. 2020, 89, 103464. [Google Scholar] [CrossRef]
- Amato-Lourenco, L.F.; Lopes Moreira, T.C.; de Oliveira Souza, V.C.; Barbosa, F.; Saiki, M.; Hilário, P.; Saldiva, N.; Mauad, T. The influence of atmospheric particles on the elemental content of vegetables in urban gardens of Sao Paulo. Brazil. Environ. Pollut. 2016, 216, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Broadley, M.; Bowen, H.; Hammond, J.; Hayden, R.; King, G.; Meacham, M.; Mead, A.; Teakle, G.; White, P. Natural genetic variation in the mineral nutrient composition of Brassica oleracea. Comp. Biochem. Physiol. Part A. Mol. Integr. Physiol. 2007, 146, S246–S247. [Google Scholar] [CrossRef]
- Chang, C.Y.; Yu, H.Y.; Chen, J.J.; Li, F.B.; Zhang, H.H.; Liu, C.P. Accumulation of heavy metals in leaf vegetables from agricultural soils and associated potential health risks in the Pearl River Delta, South China. Environ. Monit. Assess. 2014, 186, 1547–1560. [Google Scholar] [CrossRef] [Green Version]
- Agwaramgbo, L.; Thomas, C.; Grays, C.; Small, J.; Young, T. An evaluation of edible plant extracts for the phytoremediation of lead contaminated water. J. Environ. Prot. Sci. 2012, 3, 722–730. [Google Scholar] [CrossRef] [Green Version]
- Šamec, D.; Linić, I.; Salopek-Sondi, B. Salinity stress as an elicitor for phytochemicals and minerals accumulation in selected leafy vegetables of Brassicaceae. Agronomy 2021, 11, 361. [Google Scholar] [CrossRef]
- Zhou, M.; Engelmann, T.; Lutts, S. Salinity modifies heavy metals and arsenic absorption by the halophyte plant species Kosteletzkya pentacarpos and pollutant leaching from a polycontaminated substrate. Ecotoxicol. Environ. Saf. 2019, 182, 109460. [Google Scholar] [CrossRef] [PubMed]
- Fritioff, Å.; Kautsky, L.; Greger, M. Influence of temperature and salinity on heavy metal uptake by submersed plants. Environ. Pollut. 2005, 133, 265–274. [Google Scholar] [CrossRef] [PubMed]
- Idaszkin, Y.L.; Márquez, F.; Mateos-Naranjo, E.; Pollicelli, M.; Saraví Cisneros, H. Multidimensional approach to evaluate Limonium brasiliense as source of early biomarkers for lead pollution monitoring under different saline conditions. Ecol. Indic. 2019, 104, 567–575. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Element | Control Leaves | Treated Leaves | Change * |
|---|---|---|---|
| Ag | <LOD | <LOD | |
| Al | 20.8 | 52.8 | ↑ |
| As | <LOD | <LOD | |
| B | 79.3 | 52.8 | ↓ |
| Ba | LOD < × < LOQ | LOD < × < LOQ | − |
| Be | <LOD | <LOD | |
| Bi | LOD < × < LOQ | LOD < × < LOQ | |
| Ca | 8111 | 8144 | − |
| Cd | 0.090 | 0.044 | ↓ |
| Co | 0.091 | 0.108 | − |
| Cr | 2.09 | 1.03 | ↓ |
| Cu | 5.97 | 5.78 | − |
| Fe | 101 | 101 | − |
| Ga | <LOD | <LOD | |
| K | 41,572 | 44,850 | − |
| Li | 0.227 | 0.079 | ↓ |
| Mg | 11,124 | 10,057 | − |
| Mn | 205 | 185 | ↓ |
| Mo | 25.2 | 16.1 | ↓ |
| Na | 1665 | 65,752 | ↑ |
| Ni | 2.28 | 0.773 | ↓ |
| Pb | 8.34 | 0.672 | ↓ |
| Rb | 7.97 | 8.81 | ↑ |
| Se | <LOD | <LOD | |
| Sr | 42.5 | 41.9 | − |
| Te | <LOD | <LOD | |
| Tl | <LOD | <LOD | |
| U | <LOD | <LOD | |
| V | 0.043 | 0.038 | − |
| Zn | 27.2 | 21.5 | − |
| Element | Kale | White Cabbage | Chinese Cabbage | |||
|---|---|---|---|---|---|---|
| Roots | Leaves | Roots | Leaves | Roots | Leaves | |
| Al | increase | increase | increase | no significant changes | decrease | no significant changes |
| B | no significant changes | decrease | increase | no significant changes | <LOD | <LOD |
| Ba | increase | no significant changes | <LOD | <LOD | <LOD | <LOD |
| Ca | no significant changes | no significant changes | no significant changes | no significant changes | no significant changes | no significant changes |
| Cd | increase | decrease | increase | no significant changes | increase | no significant changes |
| Co | increase | no significant changes | <LOD | <LOD | <LOD | <LOD |
| Cr | increase | decrease | increase | no significant changes | increase | no significant changes |
| Cu | increase | no significant changes | increase | no significant changes | increase | no significant changes |
| Fe | increase | no significant changes | increase | decrease | decrease | no significant changes |
| K | no significant changes | no significant changes | no significant changes | no significant changes | decrease | no significant changes |
| Mg | no significant changes | no significant changes | no significant changes | no significant changes | decrease | no significant changes |
| Mn | decrease | decrease | no significant changes | no significant changes | no significant changes | decrease |
| Na | increase | increase | increase | increase | increase | increase |
| Ni | increase | decrease | increase | no significant changes | increase | no significant changes |
| Pb | no significant changes | decrease | increase | no significant changes | increase | no significant changes |
| Sr | no significant changes | no significant changes | no significant changes | no significant changes | <LOD | <LOD |
| V | increase | no significant changes | increase | no significant changes | increase | no significant changes |
| Zn | no significant changes | no significant changes | increase | no significant changes | increase | decrease |
| Parameter | ICP-OES * | ICP-MS ** |
|---|---|---|
| Instrument | Prodigy High Dispersive ICP-OES (Teledyne Leeman, Hudson, NH, USA) | Agilent 7500cx ICP-MS (Agilent, Tokyo, Japan) |
| Output power | 1100 W | 1500 W |
| Argon flows | Coolant: 18 L/min | Coolant: 15 L/min |
| Auxiliary: 0.8 L/min | Auxiliary: 0.9 L/min | |
| Nebuliser: 1 L/min | Nebuliser: 0.2 L/min | |
| Sample flow | 1.0 mL/min | 0.3 mL/min |
| Nebuliser | Pneumatic (glass concentric) | MicroMist |
| Spray chamber | Glass cyclonic | Scott double pass |
| Collison cell | ------- | off |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zeiner, M.; Juranović Cindrić, I.; Nemet, I.; Franjković, K.; Salopek Sondi, B. Influence of Soil Salinity on Selected Element Contents in Different Brassica Species. Molecules 2022, 27, 1878. https://doi.org/10.3390/molecules27061878
Zeiner M, Juranović Cindrić I, Nemet I, Franjković K, Salopek Sondi B. Influence of Soil Salinity on Selected Element Contents in Different Brassica Species. Molecules. 2022; 27(6):1878. https://doi.org/10.3390/molecules27061878
Chicago/Turabian StyleZeiner, Michaela, Iva Juranović Cindrić, Ivan Nemet, Karla Franjković, and Branka Salopek Sondi. 2022. "Influence of Soil Salinity on Selected Element Contents in Different Brassica Species" Molecules 27, no. 6: 1878. https://doi.org/10.3390/molecules27061878
APA StyleZeiner, M., Juranović Cindrić, I., Nemet, I., Franjković, K., & Salopek Sondi, B. (2022). Influence of Soil Salinity on Selected Element Contents in Different Brassica Species. Molecules, 27(6), 1878. https://doi.org/10.3390/molecules27061878