
The Use of Flavylium Salts as Dynamic Inhibitor Moieties for Human Cb5R

 ,
,  ,
,  and
and 
Abstract
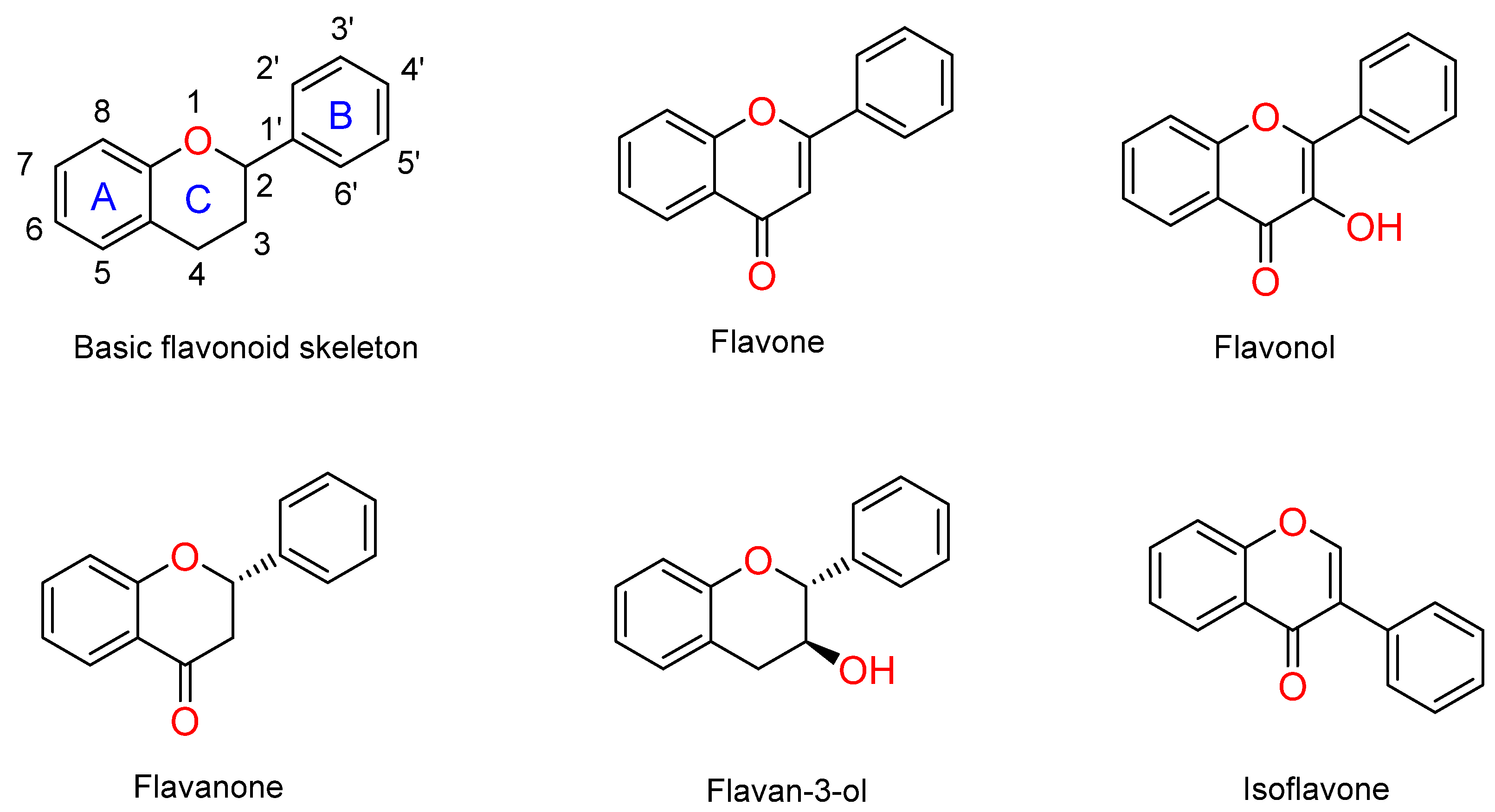
:1. Introduction
2. Results
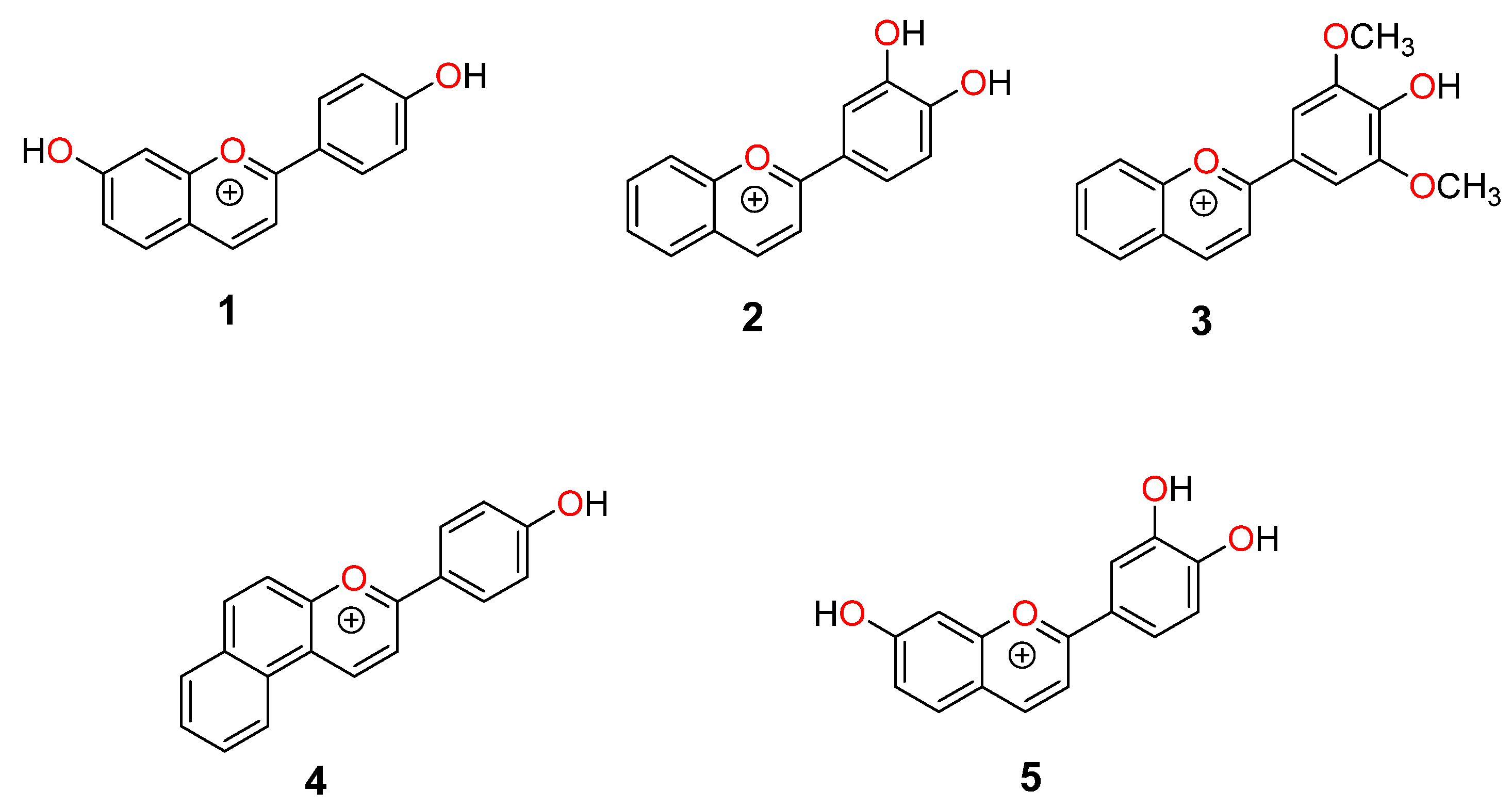
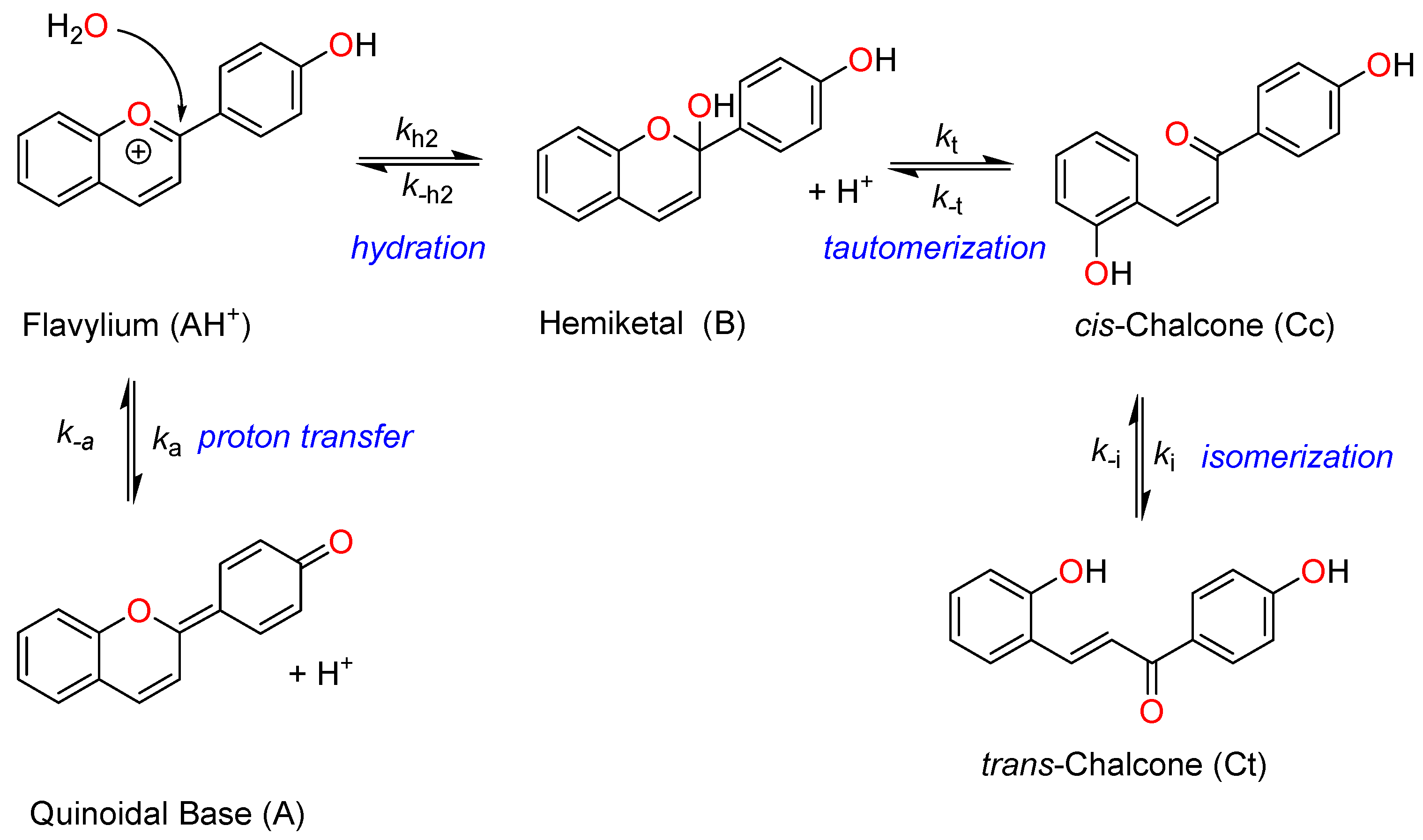
2.1. Structural Interconversion of Flavylium Compounds
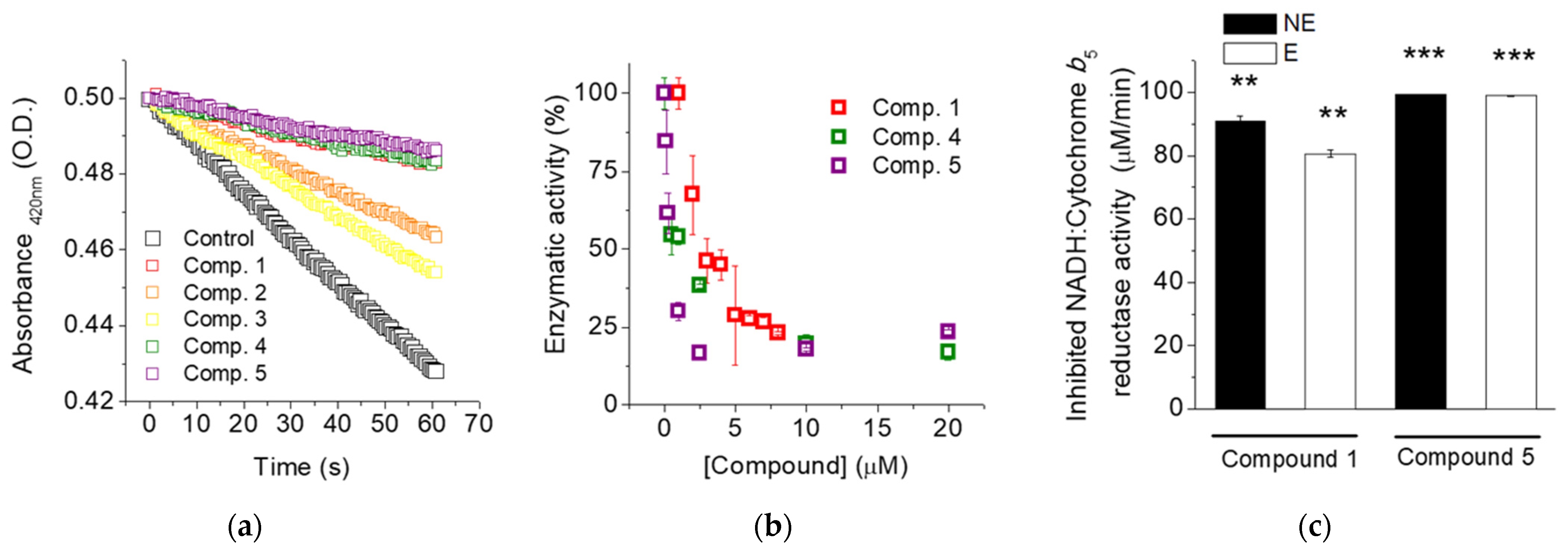
2.2. Flavylium Salts Inhibit Cb5R Activities
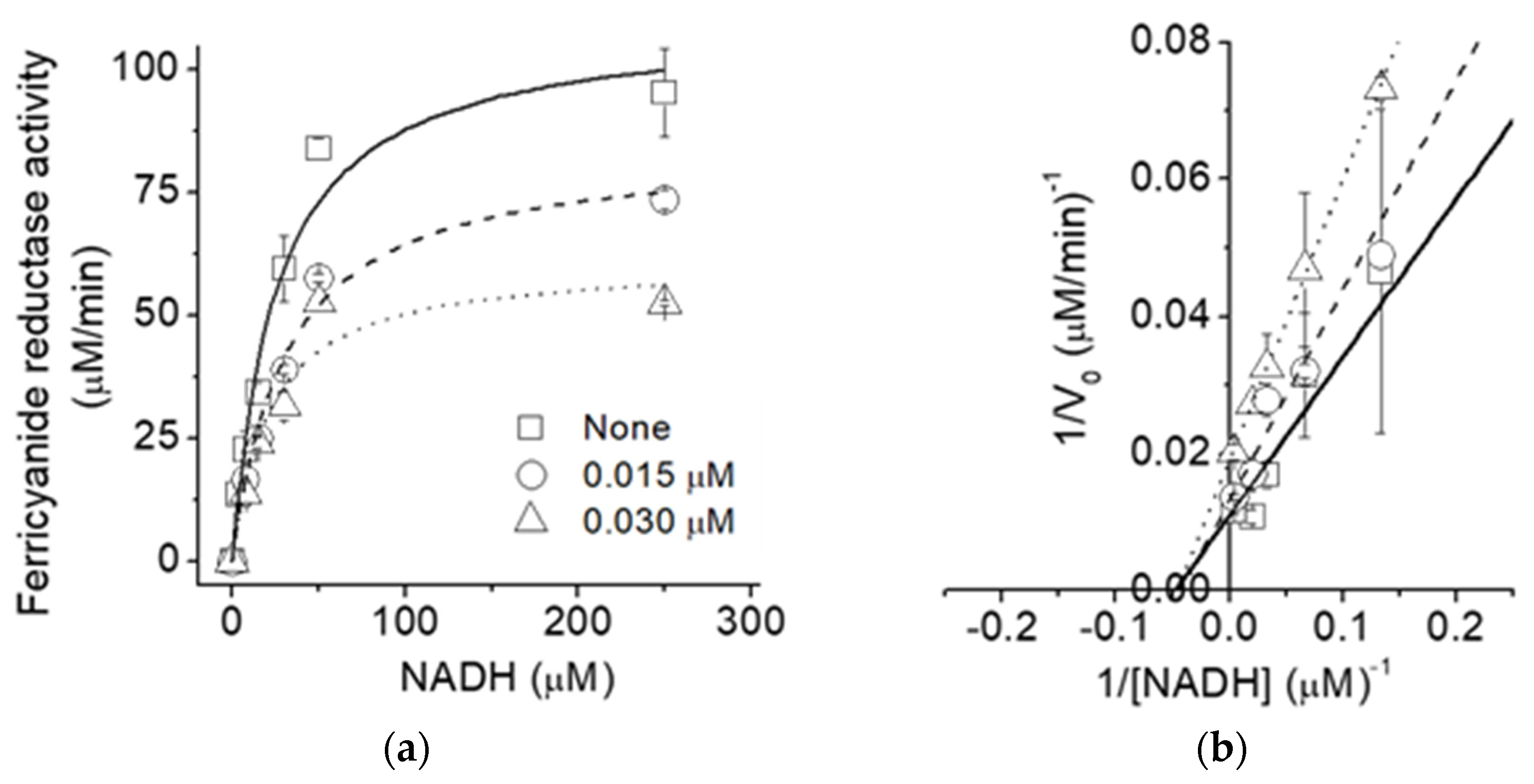
2.3. Type of Inhibition of the NADH: Ferricyanide Reductase Activity
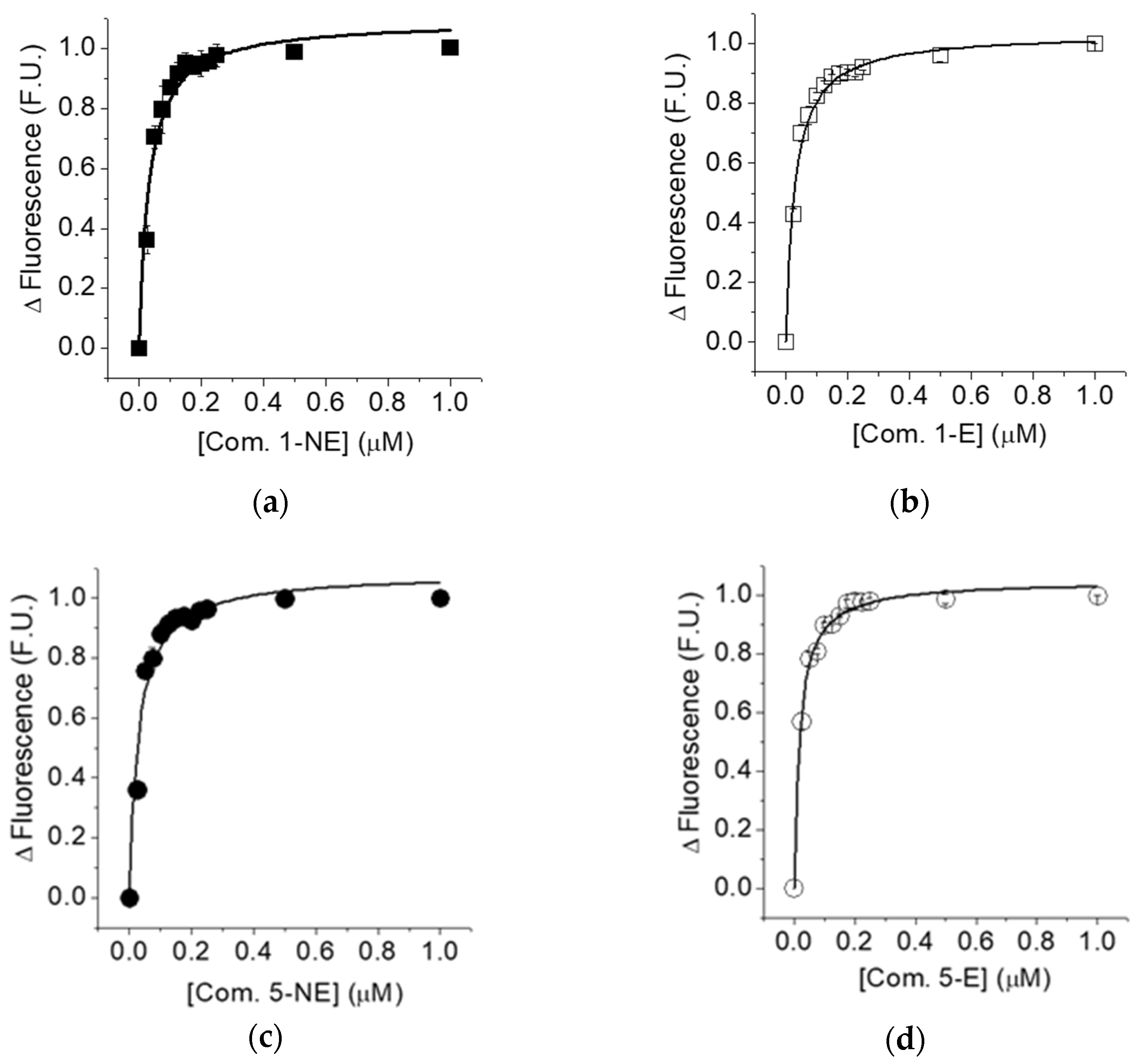
2.4. Determination of the Dissociation Constant of the Non-Equilibrated and Equilibrated Forms of compounds 1 and 5 to Cb5R
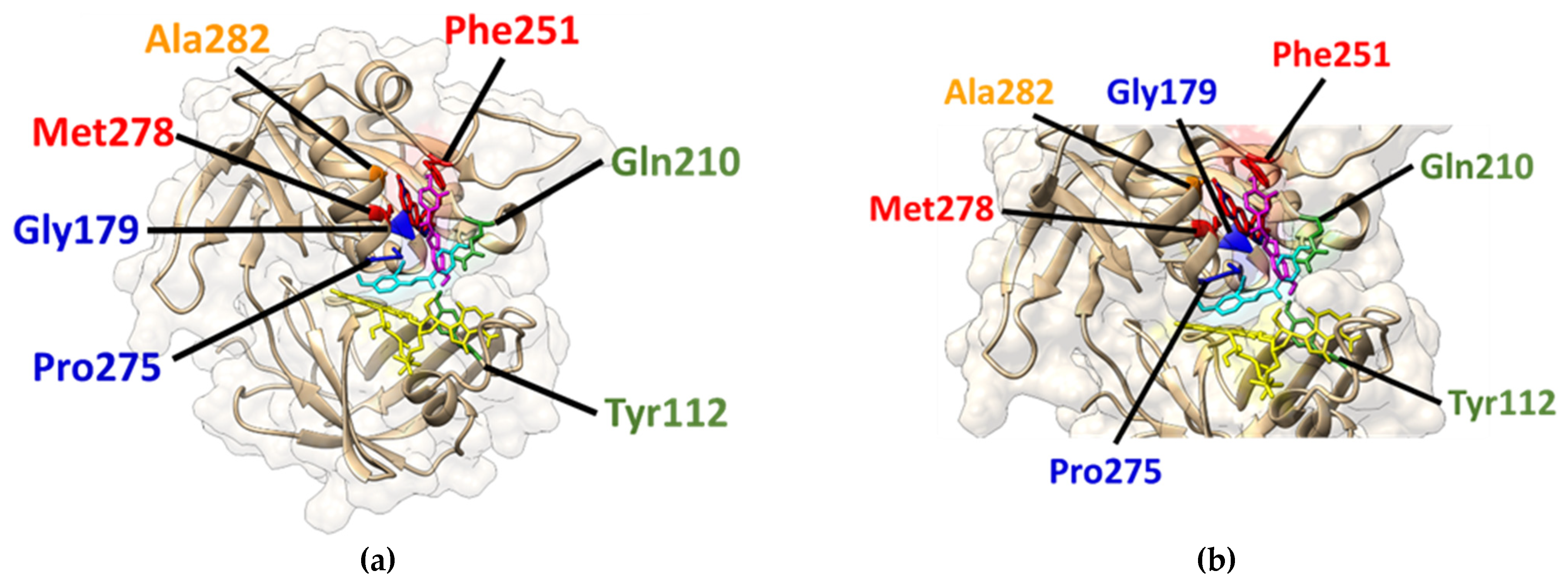
2.5. Molecular Docking for the Creation of In Silico Models for the Complex Formation between the Quinoidal Base (A) and trans-Chalcone (Ct) of compounds 1 and 5 with Cb5R
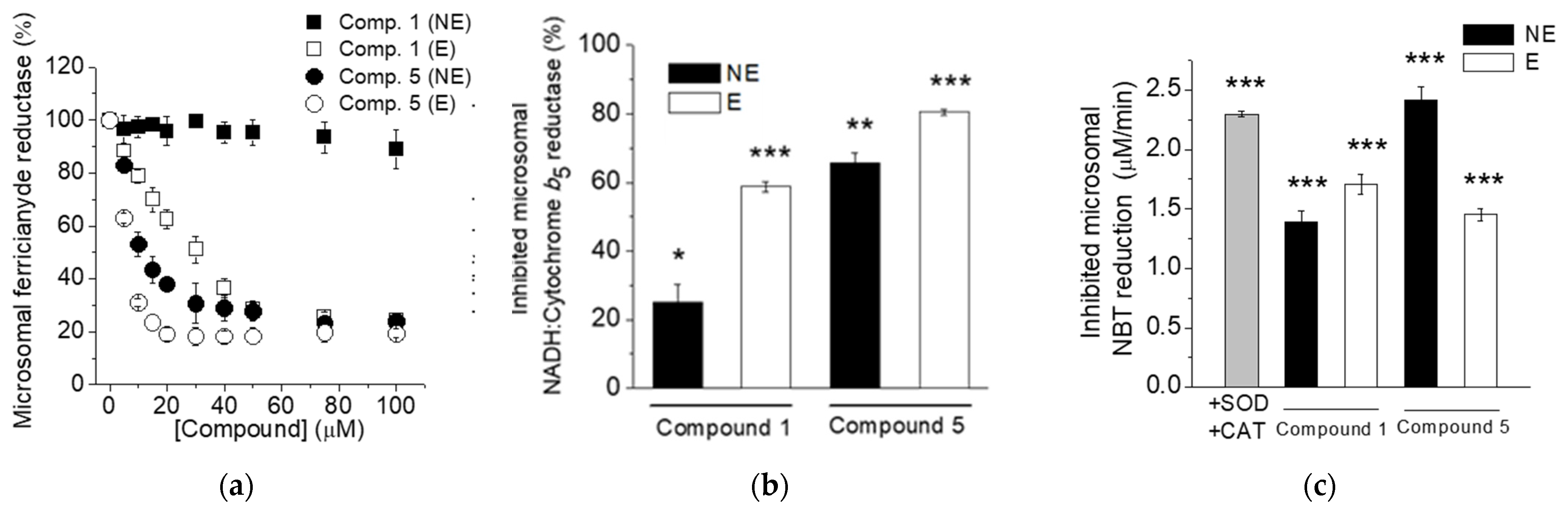
2.6. Compounds 1 and 5 Inhibit NADH-Dependent Activities of the Cb5R in Microsomes
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Synthesis
4.3. Purification of Cb5R and Cb5
4.4. Microsome Preparation from Rat Liver
4.5. Protein Determination
4.6. Preparation of the Non-Equilibrated and EQUILIBRATED Forms of the Flavylium Cations
4.7. Measurement of the Activities of the Cb5R
4.8. Determination of the Dissociation Constants for the Complex Formation between the Human Recombinant Cb5R and the Flavylium Salts
4.9. Molecular Docking Analysis
4.10. Statistic Analysis of the Data
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Samhan-Arias, A.K.; Gutierrez-Merino, C. Purified NADH-Cytochrome B5 Reductase Is a Novel Superoxide Anion Source Inhibited by Apocynin: Sensitivity to Nitric Oxide and Peroxynitrite. Free Radic. Biol. Med. 2014, 73, 174–189. [Google Scholar] [CrossRef] [PubMed]
- Samhan-Arias, A.K.; Duarte, R.O.; Martín-Romero, F.J.; Moura, J.J.G.; Gutiérrez-Merino, C. Reduction of Ascorbate Free Radical by the Plasma Membrane of Synaptic Terminals from Rat Brain. Arch. Biochem. Biophys. 2008, 469, 243–254. [Google Scholar] [CrossRef]
- Samhan-Arias, A.K.; Garcia-Bereguiain, M.A.; Gutierrez-Merino, C. Hydrogen Sulfide Is a Reversible Inhibitor of the NADH Oxidase Activity of Synaptic Plasma Membranes. Biochem. Biophys. Res. Commun. 2009, 388, 718–722. [Google Scholar] [CrossRef] [PubMed]
- Kurian, J.R.; Chin, N.A.; Longlais, B.J.; Hayes, K.L.; Trepanier, L.A. Reductive Detoxification of Arylhydroxylamine Carcinogens by Human NADH Cytochrome B5 Reductase and Cytochrome B5. Chem. Res. Toxicol. 2006, 19, 1366–1373. [Google Scholar] [CrossRef] [Green Version]
- Henderson, C.J.; McLaughlin, L.A.; Wolf, C.R. Evidence That Cytochrome B5 and Cytochrome B5 Reductase Can Act as Sole Electron Donors to the Hepatic Cytochrome P450 System. Mol. Pharmacol. 2013, 83, 1209–1217. [Google Scholar] [CrossRef] [PubMed]
- Valério, G.N.; Gutiérrez-Merino, C.; Nogueira, F.; Moura, I.; Moura, J.J.G.; Samhan-Arias, A.K. Human Erythrocytes Exposure to Juglone Leads to an Increase of Superoxide Anion Production Associated with Cytochrome B5 Reductase Uncoupling. Biochim. Et Biophys. Acta Bioenerg. 2020, 1861, 148134. [Google Scholar] [CrossRef] [PubMed]
- Samhan-Arias, A.K.; Almeida, R.M.; Ramos, S.; Cordas, C.M.; Moura, I.; Gutierrez-Merino, C.; Moura, J.J.G. Topography of Human Cytochrome B5/Cytochrome B5 Reductase Interacting Domain and Redox Alterations upon Complex Formation. Biochim. Biophys. Acta Bioenerg. 2018, 1859, 78–87. [Google Scholar] [CrossRef]
- Samhan Arias, A.K.; Gutierrez-Merino, C. Cytochrome B5 as a Pleitropic Metabolic Modular in Mammalian Cells; Thom, R., Ed.; Protein biochemistry, synthesis, structure and cellular functions; Nova Publishers: New York, NY, USA, 2014; ISBN 978-1-63117-467-4. [Google Scholar]
- Gutiérrez-Merino, C.; Martínez-Costa, O.H.; Monsalve, M.; Samhan-Arias, A.K. Structural Features of Cytochrome B5-Cytochrome B5 Reductase Complex Formation and Implications for the Intramolecular Dynamics of Cytochrome B5 Reductase. Int. J. Mol. Sci. 2021, 23, 118. [Google Scholar] [CrossRef]
- Samhan-Arias, A.K.; Garcia-Bereguiain, M.A.; Martin-Romero, F.J.; Gutierrez-Merino, C. Clustering of Plasma Membrane-Bound Cytochrome B5 Reductase within “lipid Raft” Microdomains of the Neuronal Plasma Membrane. Mol. Cell. Neurosci. 2009, 40, 14–26. [Google Scholar] [CrossRef]
- Marques-da-Silva, D.; Gutierrez-Merino, C. Caveolin-Rich Lipid Rafts of the Plasma Membrane of Mature Cerebellar Granule Neurons Are Microcompartments for Calcium/Reactive Oxygen and Nitrogen Species Cross-Talk Signaling. Cell Calcium 2014, 56, 108–123. [Google Scholar] [CrossRef]
- Samhan-Arias, A. Biochemical and Anatomical Basis of Brain Dysfunctions Caused by Cytochrome B5 Reductase Deficiency or Dysregulation. J. Neurol. Neuromedicine 2016, 1, 61–65. [Google Scholar] [CrossRef] [Green Version]
- Rahaman, M.M.; Nguyen, A.T.; Miller, M.P.; Hahn, S.A.; Sparacino-Watkins, C.; Jobbagy, S.; Carew, N.T.; Cantu-Medellin, N.; Wood, K.C.; Baty, C.J.; et al. Cytochrome B5 Reductase 3 Modulates Soluble Guanylate Cyclase Redox State and CGMP Signaling. Circ. Res. 2017, 121, 137–148. [Google Scholar] [CrossRef] [PubMed]
- Ghimire, K.; Altmann, H.M.; Straub, A.C.; Isenberg, J.S. Nitric Oxide: What’s New to NO? Am. J. Physiol. Cell Physiol. 2017, 312, C254–C262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samhan-Arias, A.K.; Fortalezas, S.; Cordas, C.M.; Moura, I.; Moura, J.J.G.; Gutierrez-Merino, C. Cytochrome B5 Reductase Is the Component from Neuronal Synaptic Plasma Membrane Vesicles That Generates Superoxide Anion upon Stimulation by Cytochrome c. Redox Biol. 2018, 15, 109–114. [Google Scholar] [CrossRef] [PubMed]
- Maeda, H.; Sawa, T.; Yubisui, T.; Akaike, T. Free Radical Generation from Heterocyclic Amines by Cytochrome B5 Reductase in the Presence of NADH. Cancer Lett. 1999, 143, 117–121. [Google Scholar] [CrossRef] [PubMed]
- Maddu, N. Diseases Related to Types of Free Radicals; Shalaby, E., Ed.; Antioxidants; IntechOpen: London, UK, 2019. [Google Scholar] [CrossRef] [Green Version]
- Gutierrez-Merino, C.; Lopez-Sanchez, C.; Lagoa, R.; Samhan-Arias, A.K.; Bueno, C.; Garcia-Martinez, V. Neuroprotective Actions of Flavonoids. Curr. Med. Chem. 2011, 18, 1195–1212. [Google Scholar] [CrossRef] [PubMed]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An Overview. J. Nutr. Sci. 2016, 5, e47. [Google Scholar] [CrossRef] [Green Version]
- Pina, F.; Melo, M.J.; Laia, C.A.T.; Parola, A.J.; Lima, J.C. Chemistry and Applications of Flavylium Compounds: A Handful of Colours. Chem. Soc. Rev. 2012, 41, 869–908. [Google Scholar] [CrossRef]
- Verma, S.; Singh, A.; Mishra, A. Molecular Construction of NADH-Cytochrome B5 Reductase Inhibition by Flavonoids and Chemical Basis of Difference in Inhibition Potential: Molecular Dynamics Simulation Study. J. App. Pharm. Sci. 2012, 2, 33–39. [Google Scholar] [CrossRef]
- Çelik, H.; Koşar, M. Inhibitory Effects of Dietary Flavonoids on Purified Hepatic NADH-Cytochrome B5 Reductase: Structure-Activity Relationships. Chem. Biol. Interact. 2012, 197, 103–109. [Google Scholar] [CrossRef]
- Cruz, L.; Basílio, N.; Mateus, N.; de Freitas, V.; Pina, F. Natural and Synthetic Flavylium-Based Dyes: The Chemistry Behind the Color. Chem. Rev. 2022, 122, 1416–1481. [Google Scholar] [CrossRef] [PubMed]
- Gavara, R.; Petrov, V.; Pina, F. Characterization of the 4′-Hydroxynaphthoflavylium Network of Chemical Reactions. Photochem. Photobiol. Sci. 2010, 9, 298–303. [Google Scholar] [CrossRef] [PubMed]
- Basílio, N.; Petrov, V.; Pina, F. Host-Guest Complexes of Flavylium Cations and Cucurbit [7]Uril: The Influence of Flavylium Substituents on the Structure and Stability of the Complex. ChemPlusChem 2015, 80, 1779–1785. [Google Scholar] [CrossRef]
- Calogero, G.; Sinopoli, A.; Citro, I.; Marco, G.D.; Petrov, V.; Diniz, A.M.; Parola, A.J.; Pina, F. Synthetic Analogues of Anthocyanins as Sensitizers for Dye-Sensitized Solar Cells. Photochem. Photobiol. Sci. 2013, 12, 883–894. [Google Scholar] [CrossRef] [PubMed]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the Speed and Accuracy of Docking with a New Scoring Function, Efficient Optimization and Multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [Green Version]
- Sanner, M.F. Python: A Programming Language for Software Integration and Development. J. Mol. Graph. Model 1999, 17, 57–61. [Google Scholar]
- Samhan-Arias, A.K.; Marques-da-Silva, D.; Yanamala, N.; Gutierrez-Merino, C. Stimulation and Clustering of Cytochrome B5 Reductase in Caveolin-Rich Lipid Microdomains Is an Early Event in Oxidative Stress-Mediated Apoptosis of Cerebellar Granule Neurons. J. Proteom. 2012, 75, 2934–2949. [Google Scholar] [CrossRef]
- Lagoa, R.; Lopez-Sanchez, C.; Samhan-Arias, A.K.; Gañan, C.M.; Garcia-Martinez, V.; Gutierrez-Merino, C. Kaempferol Protects against Rat Striatal Degeneration Induced by 3-Nitropropionic Acid. J. Neurochem. 2009, 111, 473–487. [Google Scholar] [CrossRef]
- Samhan-Arias, A.K.; Martín-Romero, F.J.; Gutiérrez-Merino, C. Kaempferol Blocks Oxidative Stress in Cerebellar Granule Cells and Reveals a Key Role for Reactive Oxygen Species Production at the Plasma Membrane in the Commitment to Apoptosis. Free Radic. Biol. Med. 2004, 37, 48–61. [Google Scholar] [CrossRef]
- López-Sánchez, C.; Martín-Romero, F.J.; Sun, F.; Luis, L.; Samhan-Arias, A.K.; García-Martínez, V.; Gutiérrez-Merino, C. Blood Micromolar Concentrations of Kaempferol Afford Protection against Ischemia/Reperfusion-Induced Damage in Rat Brain. Brain Res. 2007, 1182, 123–137. [Google Scholar] [CrossRef]
- Al Bittar, S.; Mora, N.; Loonis, M.; Dangles, O. A Simple Synthesis of 3-Deoxyanthocyanidins and Their O-Glucosides. Tetrahedron 2016, 72, 4294–4302. [Google Scholar] [CrossRef]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Bewley, M.C.; Marohnic, C.C.; Barber, M.J. The Structure and Biochemistry of NADH-Dependent Cytochrome B5 Reductase Are Now Consistent. Biochemistry 2001, 40, 13574–13582. [Google Scholar] [CrossRef]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A Visualization System for Exploratory Research and Analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | IC50 (µM) | Maximum Inhibition (%) |
|---|---|---|
| 1 | 2.14 ± 0.15 | 78 ± 1 |
| 4 | 0.82 ± 0.19 | 85 ± 3 |
| 5 | 0.18 ± 0.03 | 93 ± 1 |
| Compound | IC50 (µM) | |
|---|---|---|
| Equilibrated | Non-Equilibrated | |
| 1 | 4.31 ± 0.83 | 2.14 ± 0.15 |
| 5 | 2.14 ± 0.50 | 0.18 ± 0.03 |
| Ligand | Affinity (kcal/mol) | Residues from the Receptor that Contact the Ligand | |
|---|---|---|---|
| Cb5R | Compound 1-A | −7.5 | Gly179, Ala203, Pro275, Met278, Phe251, Ala282 |
| Compound 1-Ct | −7.2 | Tyr112, Gly179, Thr181, Gly182, Gln210, Asp214, Gly274, Pro275 | |
| Compound 5-A | −7.5 | Asn209, Phe251, Pro275, Pro277, Met278, Phe 281, Ala282 | |
| Compound 5-Ct | −7.3 | Tyr112, Gly180, Ala208, Asn209, Gln210, Thr237, Phe251, Val252, Pro275, Met278 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martínez-Costa, O.H.; Rodrigues-Miranda, L.; Clemente, S.M.; Parola, A.J.; Basilio, N.; Samhan-Arias, A.K. The Use of Flavylium Salts as Dynamic Inhibitor Moieties for Human Cb5R. Molecules 2023, 28, 123. https://doi.org/10.3390/molecules28010123
Martínez-Costa OH, Rodrigues-Miranda L, Clemente SM, Parola AJ, Basilio N, Samhan-Arias AK. The Use of Flavylium Salts as Dynamic Inhibitor Moieties for Human Cb5R. Molecules. 2023; 28(1):123. https://doi.org/10.3390/molecules28010123
Chicago/Turabian StyleMartínez-Costa, Oscar H., Laura Rodrigues-Miranda, Sofia M. Clemente, António Jorge Parola, Nuno Basilio, and Alejandro K. Samhan-Arias. 2023. "The Use of Flavylium Salts as Dynamic Inhibitor Moieties for Human Cb5R" Molecules 28, no. 1: 123. https://doi.org/10.3390/molecules28010123
APA StyleMartínez-Costa, O. H., Rodrigues-Miranda, L., Clemente, S. M., Parola, A. J., Basilio, N., & Samhan-Arias, A. K. (2023). The Use of Flavylium Salts as Dynamic Inhibitor Moieties for Human Cb5R. Molecules, 28(1), 123. https://doi.org/10.3390/molecules28010123