
Variation of Chemical Composition and Antioxidant Properties of Kiwiberry (Actinidia arguta) in a Three-Year Study




Abstract
:1. Introduction
2. Results and Discussion
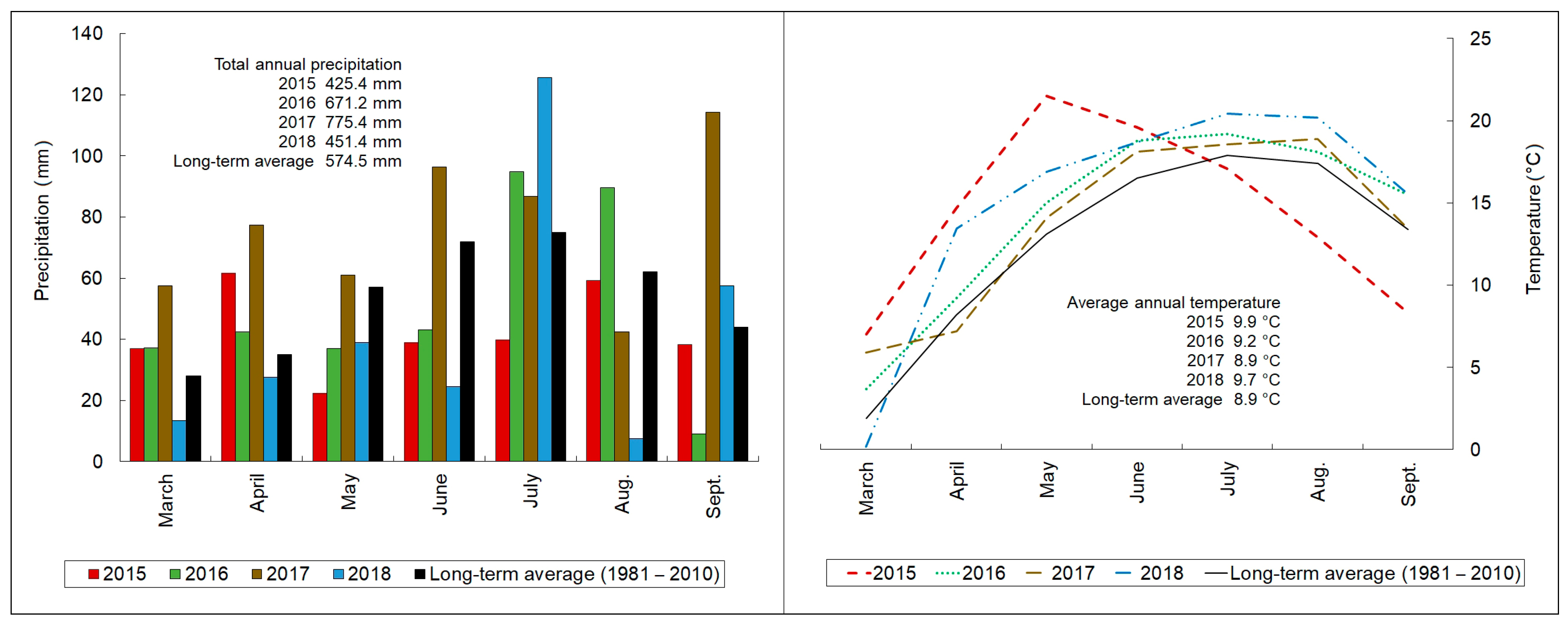
2.1. The Weather Conditions during the Experimental Seasons
2.2. The Fruit and Plant Morphology and Its Year-to-Year Variability
2.3. The Fruit Chemical Composition, Antioxidant Capacity, and Its Year-to-Year Variability
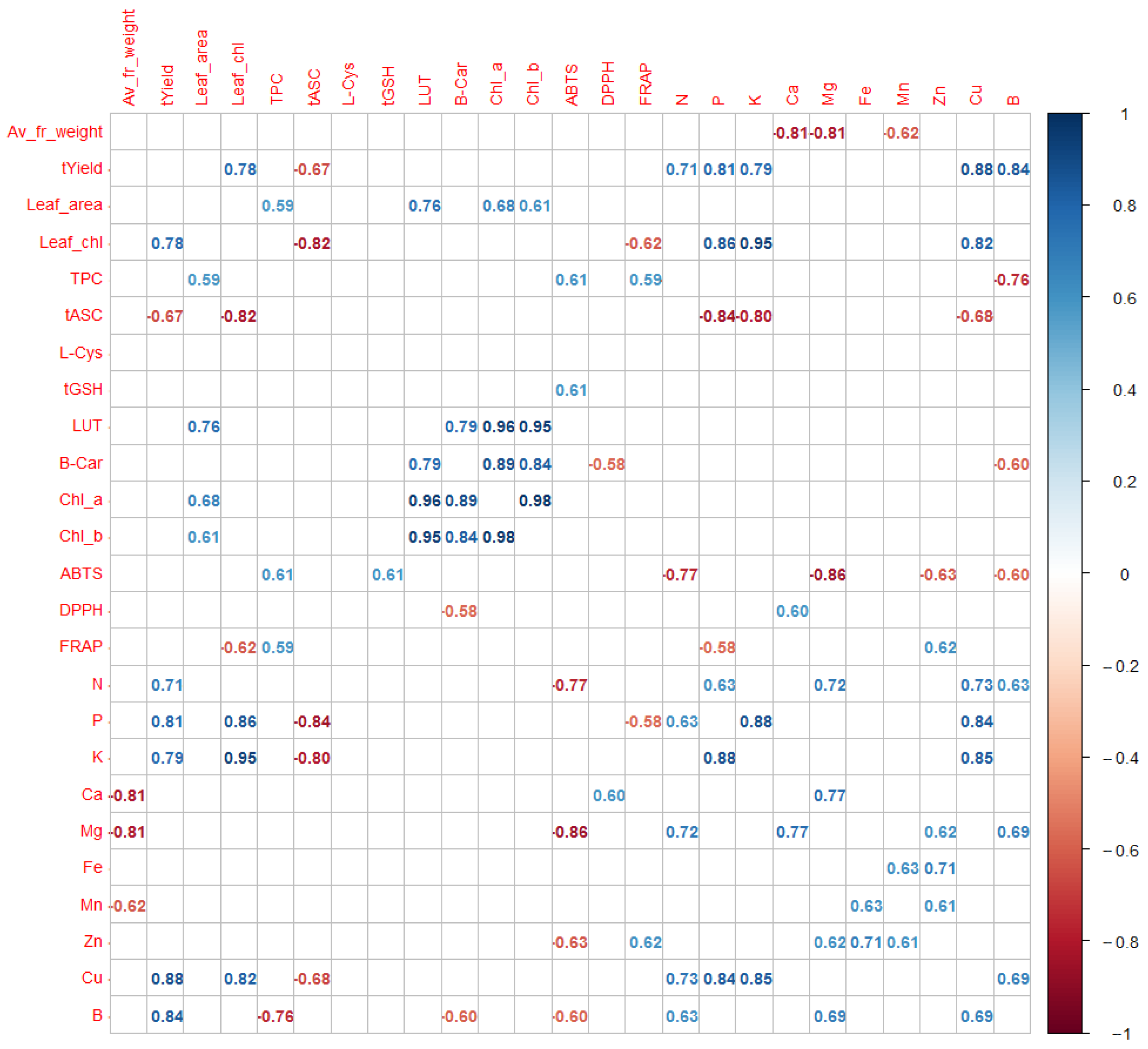
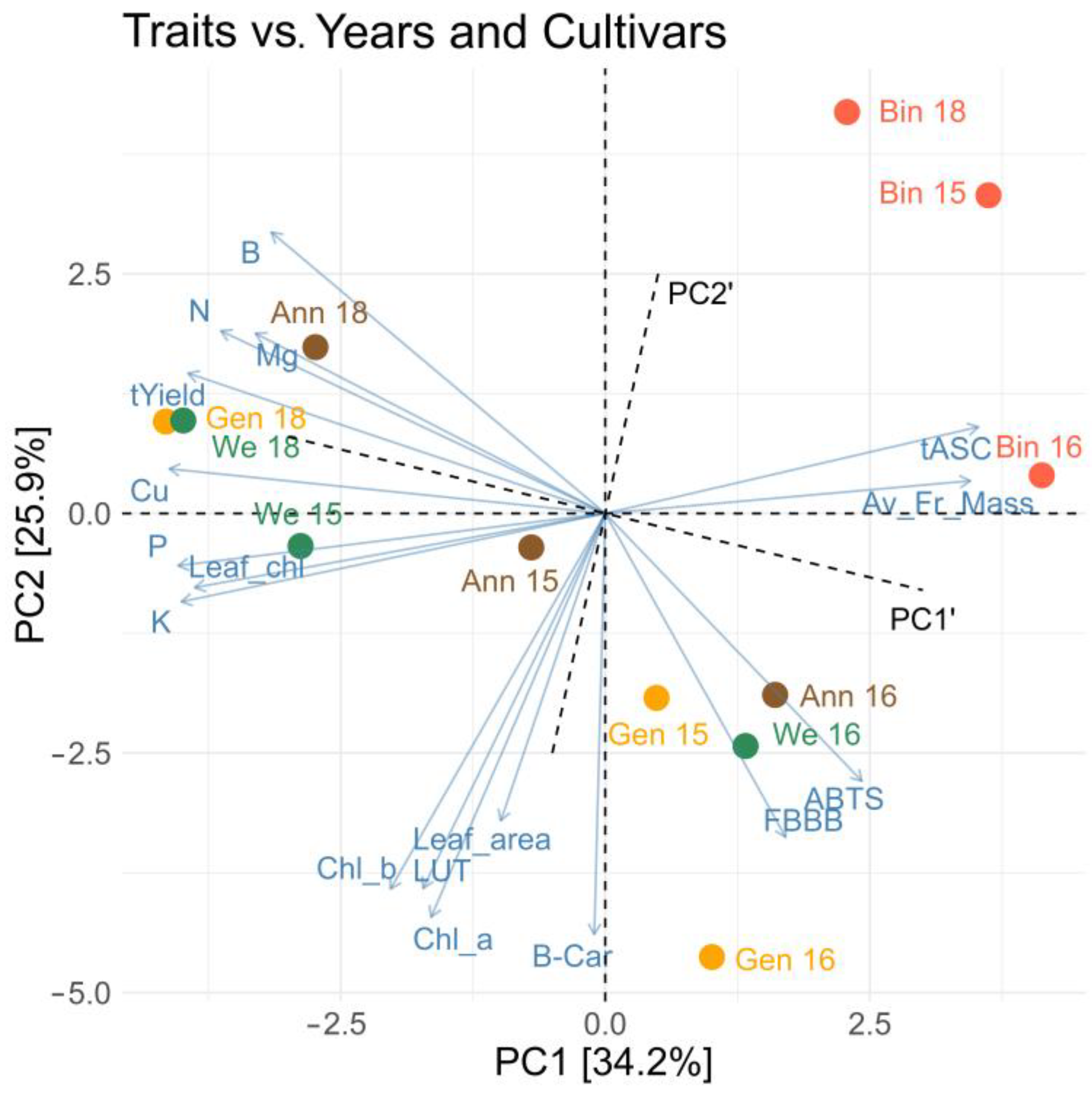
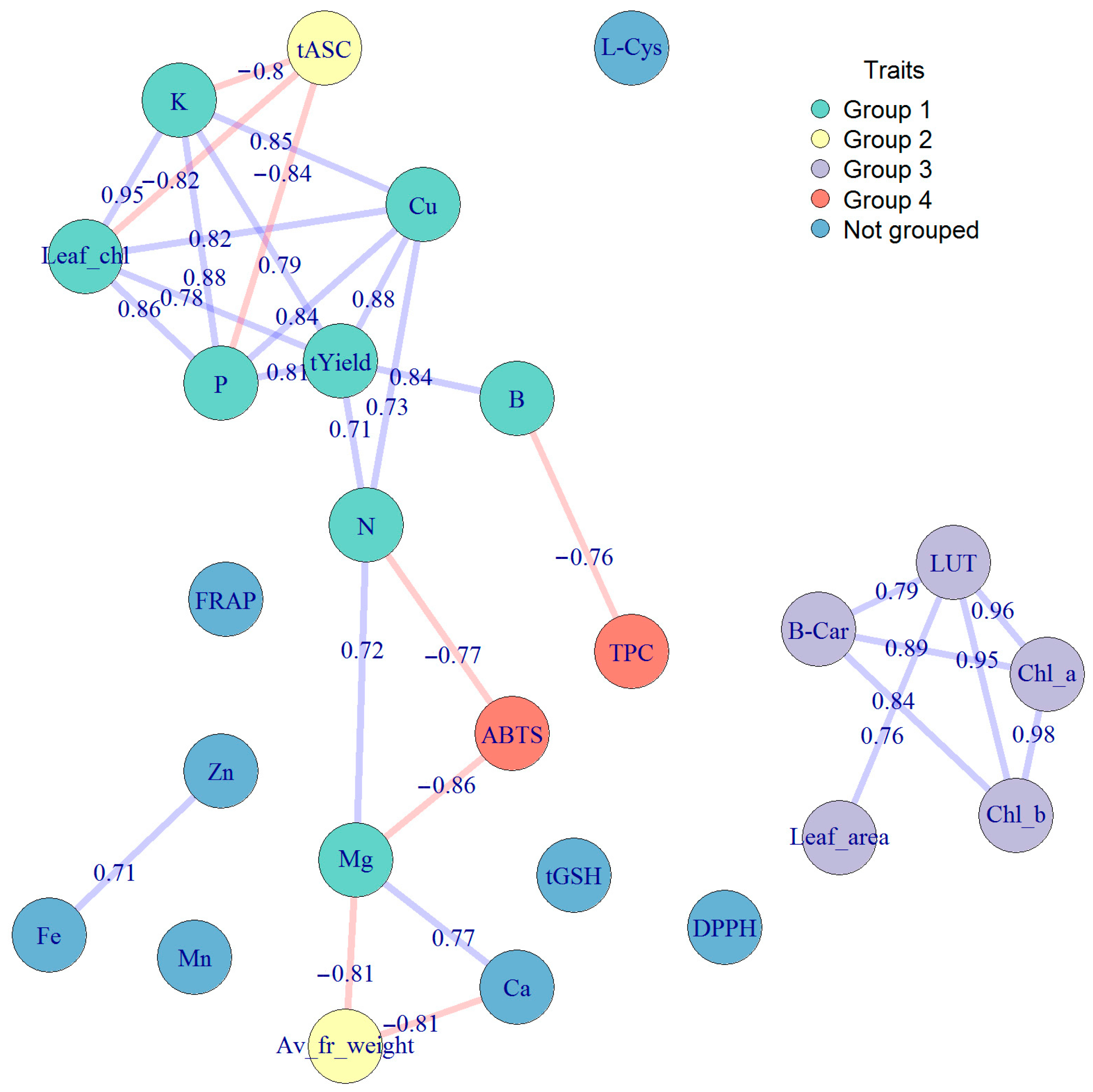
2.4. Interrelationships between Chemical Components and Physical and Physiological Properties
3. Materials and Methods
3.1. Chemicals
3.2. Experimental Design, Plant Material and Weather Conditions

3.3. Plant Morphological and Chemical Measurements
3.4. Fruit Sample Preparation and Measurements
3.5. Determination of Bioactive Compounds
3.6. Antioxidant Capacity Measurements
3.7. Determination of Fruit Macro- and Micronutrients
3.8. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Oz, A.T.; Kafkas, E. Phytochemicals in Fruits and Vegetables. In Superfood and Functional Food—An Overview of Their Processing and Utilization; Waisundara, V., Shiomi, N., Eds.; IntechOpen: London, UK, 2017. [Google Scholar] [CrossRef] [Green Version]
- Yeshi, K.; Crayn, D.; Ritmejerytė, E.; Wangchuk, P. Plant Secondary Metabolites Produced in Response to Abiotic Stresses Has Potential Application in Pharmaceutical Product Development. Molecules 2022, 27, 313. [Google Scholar] [CrossRef]
- Zhbanova, Е.V.; Lukyanchuk, I.V. Variability of the chemical composition of fruits of strawberry selected hybrid forms. Pomic. Small Fruits Cult. Russ. 2021, 64, 46–53. (In Russian) [Google Scholar] [CrossRef]
- Cossio, F.; Debersaques, F.; Latocha, P. Kiwiberry (Actinidia arguta): New perspectives for a great future. Acta Hortic. 2015, 1096, 423–434. [Google Scholar] [CrossRef]
- Latocha, P.; Vereecke, D.; Debersaques, F. Kiwiberry Commercial Production—What Stage Are We at? Acta Hortic. 2018, 1218, 559–564. [Google Scholar] [CrossRef]
- Bieniek, A.; Dragańska, E.; Pranckietis, V. Assessment of climatic conditions for Actinidia arguta cultivation in north-eastern Poland. Zemdirbyste 2016, 103, 311–318. [Google Scholar] [CrossRef] [Green Version]
- Latocha, P. The Nutritional and Health Benefits of Kiwiberry (Actinidia arguta)—A Review. Plant Foods Hum. Nutr. 2017, 72, 325–334. [Google Scholar] [CrossRef] [Green Version]
- Pinto, D.; Delerue-Matos, C.; Rodrigues, F. Bioactivity, phytochemical profile and pro-healthy properties of Actinidia arguta: A review. Food Res. Int. 2020, 136, 109449. [Google Scholar] [CrossRef]
- Leontowicz, M.; Leontowicz, H.; Jesion, I.; Bielecki, W.; Najman, K.; Latocha, P.; Park, Y.-S.; Gorinstein, S. Actinidia arguta supplementation protects aorta and liver in rats with induced hypercholesterolemia. Nutr. Res. 2016, 36, 1231–1242. [Google Scholar] [CrossRef]
- Błaszczak, W.; Latocha, P.; Jeż, M.; Wiczkowski, W. The impact of high-pressure processing on the polyphenol profile and anti-glycaemic, anti-hypertensive and anti-cholinergic activities of extracts obtained from kiwiberry (Actinidia arguta) fruits. Food Chem. 2021, 343, 128421. [Google Scholar] [CrossRef]
- Fisk, C.L.; Mc Daniel, M.R.; Strik, B.C.; Zhao, Y. Physicochemical, sensory, and nutritive qualities of hardy kiwifruit (Actinidia arguta ‘Ananasnaya’) as affected by harvest maturity and storage. J. Food Sci. 2006, 71, 204–210. [Google Scholar] [CrossRef]
- Marangi, F.; Pinto, D.; de Francisco, L.; Alves, R.C.; Puga, H.; Sut, S.; Dall’Acqua, S.; Rodrigues, F.; Oliveira, M.B.P.P. Hardy kiwi leaves extracted by multi-frequency multimode modulated technology: A sustainable and promising by-product for industry. Food Res. Int. 2018, 112, 184–191. [Google Scholar] [CrossRef]
- Silva, A.M.; Costa, P.; Delerue-Matos, C.; Latocha, P.; Rodrigues, F. Extraordinary composition of Actinidia arguta by-products as promising skin ingredient: A new challenge for cosmetic industry. Trends Food Sci. Technol. 2021, 116, 842–853. [Google Scholar] [CrossRef]
- Park, Y.-S.; Im, M.H.; Ham, K.-S.; Kang, S.-G.; Park, Y.-K.; Namiestnik, J.; Leontowicz, H.; Leontowicz, M.; Katrich, E.; Gorinstein, S. Nutritional and Pharmaceutical Properties of Bioactive Compounds in Organic and Conventional Growing Kiwifruit. Plants Foods Hum. Nutr. 2011, 68, 57–64. [Google Scholar] [CrossRef]
- Wojdyło, A.; Nowicka, P.; Oszmiański, J.; Golis, T. Phytochemical compounds and biological effects of Actinidia fruits. J. Funct. Foods 2017, 30, 194–202. [Google Scholar] [CrossRef]
- Nishiyama, I.; Yamashita, Y.; Yamanaka, M.; Shimohashi, A.; Fukuda, T.; Oota, T. Varietal difference in vitamin C content in the fruit of kiwifruit and other Actinidia species. J. Agric. Food Chem. 2004, 52, 5472–5475. [Google Scholar] [CrossRef]
- Leontowicz, H.; Leontowicz, M.; Latocha, P.; Jesion, J.; Park, Y.-S.; Katrich, E.; Barasch, D.; Nemirovski, N.; Gorinstein, S. Bioactivity and nutritional properties of hardy kiwi fruit Actinidia arguta in comparison with Actinidia deliciosa ‘Hayward’ and Actinidia eriantha ‘Bidan’. Food Chem. 2016, 196, 281–291. [Google Scholar] [CrossRef]
- Bieniek, A. Mineral composition of fruits of Actinidia arguta and Actinidia purpurea and some of their hybrid cultivars grown in north-eastern Poland. Pol. J. Environ. Stud. 2012, 21, 1543–1550. [Google Scholar]
- Drzewiecki, J.; Latocha, P.; Leontowicz, H.; Leontowicz, M.; Park, Y.S.; Najman, K.; Weisz, M.; Ezra, A.; Gorinstein, S. Analytical methods applied to characterization of Actinidia arguta, Actinidia deliciosa and Actinidia eriantha kiwi fruit cultivars. Food Anal. Methods 2016, 9, 1353–1366. [Google Scholar] [CrossRef] [Green Version]
- Stefaniak, J.; Przybył, J.; Latocha, P.; Łata, B. Bioactive compounds, total antioxidant activity and yield of kiwiberry fruit under different nitrogen regimes in field conditions. J. Sci. Food Agric. 2020, 100, 3832–3840. [Google Scholar] [CrossRef]
- Kim, J.G.; Beppu, K.; Kataoka, I. Varietal differences in phenolic content and astringency in skin and flesh of hardy kiwifruit resources in Japan. Sci. Hortic. 2009, 120, 551–554. [Google Scholar] [CrossRef]
- Latocha, P.; Łata, B.; Stasiak, A. Phenolics, ascorbate and the antioxidant potential of kiwiberry vs. common kiwifruit: The effect of cultivar and tissue type. J. Funct. Foods 2015, 19, 155–163. [Google Scholar] [CrossRef]
- Richardson, A.C.; Marsh, K.B.; Boldingh, H.L.; Pickering, A.H.; Bulley, S.M.; Frearson, N.J.; Ferguson, A.R.; Thornber, S.E.; Bolitho, K.M.; Macrae, E.A. High growing temperatures reduce fruit carbohydrate and vitamin C in kiwifruit. Plant Cell Environ. 2004, 27, 423–435. [Google Scholar] [CrossRef]
- Nishiyama, I.; Fukuda, T.; Oota, T. Genotype differences in chlorophyll, lutein, and b-carotene contents in the fruits of Actinidia species. J. Agric. Food Chem. 2005, 53, 6403–6407. [Google Scholar] [CrossRef]
- Latocha, P.; Wołosiak, R.; Worobiej, E.; Krupa, T. Clonal differences in antioxidant activity and bioactive constituents of hardy kiwifruit (Actinidia arguta) and its year-to-year variability. J. Sci. Food Agric. 2013, 93, 1412–1419. [Google Scholar] [CrossRef]
- Łata, B.; Przeradzka, M.; Bińkowska, M. Great differences in antioxidant properties exist between 56 apple cultivars and vegetation seasons. J. Agric. Food Chem. 2005, 53, 8970–8978. [Google Scholar] [CrossRef]
- Latocha, P.; Debersaques, F.; Decorte, J. Varietal Differences in the Mineral Composition of Kiwiberry—Actinidia arguta (Siebold et Zucc.) Planch. ex. Miq. Acta Hortic. 2015, 1096, 479–486. [Google Scholar] [CrossRef]
- Gralak, M.A.; Lasocka, I.; Leontowicz, M.; Leontowicz, H.; Latocha, P.; Gorinstein, S. Bioavailability of Macro- and Microelements in Rats Fed Hypercholesterolemic Diets Containing Actinidia arguta Fruits. Foods 2022, 11, 1633. [Google Scholar] [CrossRef]
- Bieniek, A.; Dragańska, E. Content of macroelements in fruits of Ukrainian cultivars of Hardy kiwifruit and Actinidia charta depending on the weather conditions during the phonological phases. J. Elem. 2013, 18, 23–38. [Google Scholar] [CrossRef]
- Gündüz, K.; Serҫe, S.; Hancock, J.F. Variation among highbush and rabbiteye cultivars of blueberry for fruit quality and phytochemical characteristics. J. Food Compos. Anal. 2015, 38, 69–79. [Google Scholar] [CrossRef]
- Swoczyna, T.; Łata, B.; Stasiak, A.; Stefaniak, J.; Latocha, P. JIP-test in assessing sensitivity to nitrogen deficiency in two cultivars of Actinidia arguta (Siebold et Zucc.) Planch. ex Miq. Photosynthetica 2019, 57, 646–658. [Google Scholar] [CrossRef]
- Stefanelli, D.; Goodwin, I.; Jones, R. Minimal nitrogen and water use in horticulture: Effects on quality and content of selected nutrients. Food Res. Int. 2010, 43, 1833–1843. [Google Scholar] [CrossRef]
- Stefaniak, J.; Stasiak, A.; Latocha, P.; Łata, B. Seasonal changes in macronutrients in the leaves and fruit of kiwiberry: Nitrogen level and cultivar effects. Commun. Soil Sci. Plant Anal. 2019, 50, 2913–2926. [Google Scholar] [CrossRef]
- Osborne, B.A.; Raven, J.A. Light absorption by plants and its implications for photosynthesis. Biol. Rev. Camb. Philos. Soc. 1986, 61, 1–60. [Google Scholar] [CrossRef]
- Sid’ko, A.F.; Botvich, I.Y.; Pis’man, T.I.; Shevyrnogov, A.P. Estimation of the chlorophyll content and yield of grain crops via their chlorophyll potential. Biophysics 2017, 62, 456–459. [Google Scholar] [CrossRef]
- Gholamin, R.; Khayatnezhad, M. The effect of end season drought stress on the chlorophyll content, chlorophyll fluorescence parameters and yield in maize cultivars. Sci. Res. Essays 2011, 6, 5351–5357. [Google Scholar] [CrossRef]
- Ghosh, M.; Pal, A.K.; Pal, S.K.; De, D.K. Relationship of leaf area and chlorophyll content with yield of aromatic rice. Indian J. Plant Physiol. 2003, 8, 199–200. [Google Scholar]
- Smith, G.S.; Buwalda, J.G.; Clark, C.J. Nutrient dynamics of a kiwifruit ecosystem. Sci. Hortic. 1988, 37, 87–109. [Google Scholar] [CrossRef]
- Ludemann, C.I.; Gruere, A.; Heffer, P.; Dobermann, A. Global data on fertilizer use by crop and by country. Sci. Data 2022, 9, 501. [Google Scholar] [CrossRef]
- Zhao, Z.; Tong, Y.; Wang, J. Nutrient Uptake and Distribution in Field-Grown Kiwifruit Vines. Acta Hortic. 2013, 984, 219–226. [Google Scholar] [CrossRef]
- Tränknera, M.; Ershad Tavakol, E.; Jákli, B. Functioning of potassium and magnesium in photosynthesis, photosynthate translocation and photoprotection. Physiol. Plant. 2018, 163, 414–431. [Google Scholar] [CrossRef] [Green Version]
- Yruela, I. Copper in plants. Braz. J. Plant Physiol. 2005, 17, 145–156. [Google Scholar] [CrossRef]
- Brown, P.H.; Bellaloui, N.; Wimmer, M.A.; Bassil, E.S.; Ruiz, J.; Hu, H.; Pfeffer, H.; Dannel, F.; Römheld, V. Boron in Plant Biology. Plant Biol. 2002, 4, 205–223. [Google Scholar] [CrossRef]
- Palmer, J.W.; Giuliani, R.; Adams, H.M. Effect of crop load on fruiting and leaf photosynthesis of ‘Braeburn’/M.26 apple trees. Tree Physiol. 1997, 17, 741–746. [Google Scholar] [CrossRef] [Green Version]
- Sidhu, R.S.; Bound, A.A.; Hunt, I. Crop Load and Thinning Methods Impact Yield, Nutrient Content, Fruit Quality, and Physiological Disorders in ‘Scilate’ Apples. Agronomy 2022, 12, 1989. [Google Scholar] [CrossRef]
- Testolin, R. Kiwifruit yield efficiency, plant density, and bud number per surface unit. J. Am. Soc. Hortic. Sci. 1990, 115, 704–707. [Google Scholar] [CrossRef] [Green Version]
- Kopsell, D.A.; Kopsell, D.E.; Lefsrud, M.G. Variation in lutein, β-carotene, and chlorophyll concentrations among Brassica oleraceae cultings and seasons. HortScience 2004, 39, 361–364. [Google Scholar] [CrossRef]
- Ihl, M.; Shene, C.; Scheuermann, E.; Bifani, V. Correlation of pigment content through colour determination using tristimulus values in a green leafy vegetable, Swiss chard. J. Sci. Food Agric. 2006, 66, 527–531. [Google Scholar] [CrossRef]
- Mou, B. Genetic Variation of Beta-carotene and Lutein contents in Lettuce. J. Am. Soc. Hortic. Sci. 2005, 130, 870–876. [Google Scholar] [CrossRef] [Green Version]
- Holasová, M.; Dostálová, R.; Fiedlerová, V.; Horáček, J. Variability of Lutein Content in Peas (Pisum sativum L.) in Relation to the Variety, Season and Chlorophyll Content. Czech J. Food Sci. 2009, 27, S188–S191. [Google Scholar] [CrossRef] [Green Version]
- Delgado-Pelayo, R.; Gallardo-Guerrero, L.; Hornero-Méndez, D. Chlorophyll and carotenoid pigments in the peel and flesh of commercial apple fruit varieties. Food Res. Int. 2014, 65, 272. [Google Scholar] [CrossRef] [Green Version]
- Strik, B.C.; Davis, A.J. Growing Kiwifruit. A Guide to Kiwiberries and Fuzzy Kiwifruit for Pacific Northwest Producers, PNW 507; A Pacific Northwest Extension Publication; PNW Publishing: Eugene, OR, USA, 2021; Available online: https://horticulture.oregonstate.edu/sites/agscid7/files/horticulture/attachments/54_ananasnaya_japs_60106-112_2006_strik_hummer.pdf (accessed on 12 August 2022).
- Komosa, A.; Roszyk, J.; Mieloch, M. Content of nutrients in soils of highbush blueberry (Vaccinium corymbosum L.) plantations in Poland in a long-term study. J. Elem. 2017, 22, 1193–1207. [Google Scholar] [CrossRef]
- Escarpa, A.; Gonzalez, M.C. High-performance liquid chromatography with diode-array detection for the determination of phenolic compounds in peel and pulp from different apple varieties. J. Chromatogr. A 1998, 823, 331–337. [Google Scholar] [CrossRef] [PubMed]
- Medina, M.B. Determination of the total phenolics in juices and superfruits by novel chemical method. J. Funct. Foods 2011, 3, 79–87. [Google Scholar] [CrossRef]
- Łata, B. Variability in enzymatic and non-enzymatic antioxidants in red and green-leafy kale in relation to soil type and N-level. Sci. Hortic. 2014, 68, 38–45. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free. Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- R Core Team R. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022; Available online: https://www.R-project.org/ (accessed on 10 October 2022).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Average Fruit Weight (g) | Total Yield (kg/plant) | Leaf Blade Area (mm2) | Leaf Chlorophyll Concentration (a. u.) | Fruit Chemical Composition | |||
|---|---|---|---|---|---|---|---|---|
| Phenolic Compounds(TPC) | Ascorbate (tASC) | Thiol Compounds | ||||||
| L-Cys | tGSH | |||||||
| mg/kg FW | ||||||||
| Geneva | ||||||||
| 2015 | 6.5 ± 0.1 b | 27.8 ± 7.6 b | 7196 ± 83 | 18.7 ± 1.9 c | 1462 ± 21 a | 750 ± 134 a | 2.04 ± 0.03 a | 197 ± 2 a |
| 2016 | 8.1 ± 0.4 a | 25.4 ± 3.7 b | 7333 ± 687 | 23.8 ± 3.3 b | 1353 ± 190 a | 650 ± 68 a | 1.34 ± 0.19 c | 219 ± 9 a |
| 2018 | 5.8 ± 0.3 c | 72.8 ± 7.0 a | 6389 ± 984 | 28.0 ± 3.6 a | 851 ± 96 b | 484 ± 25 b | 1.69 ± 0.10 b | 208 ± 1 a |
| Weiki | ||||||||
| 2015 | 6.9 ± 0.2 c | 40.7 ± 6.8 a | 5102 ± 282 | 21.6 ± 0.9 b | 815 ± 174 a | 610 ± 63 a | 2.00 ± 0.71 a | 113 ± 26 c |
| 2016 | 9.5 ± 0.7 a | 25.6 ± 3.8 b | 5259 ± 263 | 22.6 ± 1.6 b | 873 ± 74 a | 561 ± 81 a | 2.31 ± 0.01 a | 189 ± 14 a |
| 2018 | 7.9 ± 0.2 b | 60.1 ± 19.2 a | 5281 ± 432 | 28.8 ± 2.5 a | 367 ± 76 b | 484 ± 18 a | 2.15 ± 0.36 a | 151 ± 6 b |
| Bingo | ||||||||
| 2015 | 8.6 ± 0.8 b | 21.0 ± 4.7 | 4371 ± 217 | 15.8 ± 2.5 b | 681 ± 133 b | 986 ± 56 a | 1.48 ± 0.33 c | 117 ± 14 c |
| 2016 | 9.4 ± 1.4 ab | 30.6 ± 7.5 | 5020 ± 429 | 16.1 ± 1.4 ab | 1305 ± 137 a | 758 ± 69 b | 2.56 ± 0.15 a | 174 ± 25 a |
| 2018 | 9.4 ± 0.5 a | 34.4 ± 8.7 | 4329 ± 182 | 19.5 ± 0.8 a | 374 ± 95 c | 643 ± 16 c | 2.01 ± 0.24 b | 145 ± 19 b |
| Anna | ||||||||
| 2015 | 6.7 ± 0.1 c | 33.1 ± 3.5 b | 4722 ± 31 | 20.0 ± 1.4 b | 1157 ± 54 a | 552 ± 84 | 5.11 ± 0.34 a | 178 ± 5 c |
| 2016 | 8.9 ± 0.3 a | 29.6 ± 2.0 b | 5050 ± 434 | 22.0 ± 1.0 ab | 1093 ± 132 a | 656 ± 30 | 1.41 ± 0.19 c | 227 ± 7 a |
| 2018 | 7.2 ± 0.3 b | 60.4 ± 9.1 a | 4950 ± 823 | 24.8 ± 2.3 a | 372 ± 82 b | 575 ± 21 | 3.26 ± 0.23 b | 202 ± 6 b |
| Global cultivar effect (2015, 2016, and 2018) | ||||||||
| Geneva | 6.8 ± 1.1 C | 42.0 ± 23.8 AB | 6973 ± 746 A | 23.5 ± 4.8 A | 1222± 302 A | 628 ± 139 B | 1.69 ± 0.32 B | 208 ± 14 A |
| Weiki | 8.1 ± 1.2 B | 42.1 ± 18.2 A | 5214 ± 302 B | 24.3 ± 3.7 A | 685 ± 261 C | 552 ± 76 B | 2.15 ± 0.42 B | 151 ± 36 B |
| Bingo | 9.1 ± 0.9 A | 28.7 ± 8.6 B | 4573 ± 423 B | 17.1 ± 2.3 B | 787 ± 424 BC | 796 ± 158 A | 2.02 ± 0.52 B | 145 ± 30 B |
| Anna | 7.6 ± 1.0 BC | 41.0 ± 15.4 AB | 4907 ± 488 B | 22.3 ± 2.6 A | 874 ± 386 B | 594 ± 66 B | 3.26 ± 1.52 A | 202 ± 22 A |
| Year effect | ||||||||
| 2015 | 7.2 ± 0.9 C | 30.6 ± 9.0 B | 5347 ± 1158 | 19.0 ± 2.7 C | 1029 ± 332 A | 724 ± 191 A | 2.66 ± 1.54 A | 151 ± 41 C |
| 2016 | 9.0 ± 0.9 A | 27.8 ± 4.7 B | 5665 ± 1089 | 21.1 ± 3.6 B | 1156 ± 232 A | 656 ± 91 A | 1.91 ± 0.58 C | 202 ± 26 A |
| 2018 | 7.6 ± 1.4 B | 56.9 ± 17.8 A | 5237 ± 974 | 25.3 ± 4.4 A | 491 ± 230 B | 547 ± 72 B | 2.28 ± 0.65 B | 177 ± 32 B |
| Interaction | <0.05 | <0.01 | Ns | <0.05 | <0.01 | <0.001 | <0.001 | <0.05 |
| Component | Fruit Pigment Composition | Antioxidant Capacity | |||||
|---|---|---|---|---|---|---|---|
| Lutein (LUT) | βeta-Carotene (B-Car) | Chlorophyll_a (Chl_a) | Chlorophyll_b (Chl_b) | ABTS | DPPH | FRAP | |
| mg/kg FW | mmol TE/kg FW | ||||||
| Geneva | |||||||
| 2015 | 2.99 ± 0.37 a | 2.62 ± 0.38 b | 20.34 ± 2.46 b | 7.58 ± 0.78 a | 21.7 ± 1.7 b | 35.5 ± 2.8 a | 47.3 ± 1.4 a |
| 2016 | 3.12 ± 0.18 a | 3.69 ± 0.05 a | 24.67 ± 1.32 a | 8.01 ± 0.79 a | 43.0 ± 5.5 a | 19.2 ± 4.0 b | 21.4 ± 3.6 b |
| 2018 | 1.97 ± 0.02 b | 1.70 ± 0.07 c | 14.95 ± 0.15 c | 5.47 ± 0.04 b | 21.5 ± 2.3 b | 31.9 ± 3.1 a | 17.9 ± 1.8 c |
| Weiki | |||||||
| 2015 | 2.72 ± 0.20 | 2.75 ± 0.09 b | 20.16 ± 0.99 a | 7.86 ± 0.16 a | 12.2 ± 2.4 b | 24.4 ± 1.4 a | 27.5 ± 4.6 a |
| 2016 | 2.51 ± 0.09 | 3.44 ± 0.04 a | 20.09 ± 0.85 a | 7.17 ± 0.10 ab | 28.6 ± 1.8 a | 11.0 ± 2.3 b | 14.5 ± 1.6 b |
| 2018 | 2.45 ± 0.09 | 2.40 ± 0.06 b | 18.57 ± 0.20 b | 6.93 ± 0.13 b | 15.4 ± 1.2 b | 22.4 ± 1.2 a | 12.0 ± 0.3 b |
| Bingo | |||||||
| 2015 | 1.31 ± 0.06 a | 1.44 ± 0.10 b | 9.12 ± 0.52 ab | 3.31 ± 0.15 a | 15.1 ± 0.9 c | 21.9 ± 0.5 b | 35.6 ± 2.6 a |
| 2016 | 1.52 ± 0.02 a | 2.26 ± 0.05 a | 11.10 ± 0.26 a | 3.70 ± 0.13 a | 39.8 ± 3.3 a | 21.8 ± 2.9 b | 24.0 ± 0.8 b |
| 2018 | 0.92 ± 0.03 b | 1.30 ± 0.02 b | 7.13 ± 0.27 b | 2.27 ± 0.07 b | 20.3 ± 2.5 b | 29.4 ± 3.2 a | 15.9 ± 2.5 c |
| Anna | |||||||
| 2015 | 1.96 ± 0.03 | 2.65 ± 0.27 b | 16.12 ± 0.48 b | 6.07 ± 0.01 a | 16.1 ± 0.4 b | 17.3 ± 0.8 b | 39.6 ± 0.7 a |
| 2016 | 2.03 ± 0.25 | 3.10 ± 0.05 a | 19.09 ± 0.86 a | 6.95 ± 0.34 a | 33.7 ± 5.4 a | 17.1 ± 3.2 b | 17.7 ± 2.6 b |
| 2018 | 1.82 ± 0.05 | 2.25 ± 0.03 c | 14.45 ± 0.53 b | 5.12 ± 0.22 b | 16.5 ± 2.4 b | 24.8 ± 3.1 a | 11.6 ± 0.9 c |
| Global cultivar effect (2015, 2016, and 2018) | |||||||
| Geneva | 2.69 ± 0.59 A | 2.67 ± 0.88 A | 20.0 ± 4.4 A | 7.02 ± 1.30 A | 28.8 ± 11.1 A | 28.9 ± 7.9 A | 28.9 ± 14.1 A |
| Weiki | 2.56 ± 0.17 A | 2.86 ± 0.46 A | 19.6 ± 1.0 A | 7.32 ± 0.44 A | 18.7 ± 7.7 C | 19.3 ± 6.4 C | 18.0 ± 7.6 C |
| Bingo | 1.25 ± 0.27 C | 1.67 ± 0.45 B | 9.1 ± 1.8 C | 3.10 ± 0.65 C | 25.1 ± 11.5 AB | 24.4 ± 4.4 AB | 25.1 ± 8.8 AB |
| Anna | 1.94 ± 0.15 B | 2.67 ± 0.39 A | 16.6 ± 2.1 B | 6.05 ± 0.82 B | 22.1 ± 9.2 BC | 19.7 ± 4.4 BC | 22.9 ± 12.8 B |
| Year effect | |||||||
| 2015 | 2.25 ± 0.71 A | 2.37 ± 0.60 B | 16.4 ±4.9 B | 6.21 ± 1.91 A | 16.3 ± 3.8 C | 24.8 ± 7.1 A | 37.5 ± 7.8 A |
| 2016 | 2.29 ± 0.63 A | 3.12 ± 0.56 A | 18.7 ± 5.2 A | 6.46 ± 1.75 A | 36.3 ± 6.9 A | 17.3 ± 4.9 B | 19.4 ± 4.3 B |
| 2018 | 1.79 ± 0.58 B | 1.92 ± 0.46 C | 13.8 ± 4.3 C | 4.94 ± 1.77 B | 18.4 ± 3.2 B | 27.1 ± 4.6 A | 14.3 ± 3.1 C |
| Interaction | <0.001 | <0.001 | <0.001 | <0.001 | Ns | <0.001 | <0.001 |
| Group | Macronutrients (% DW) | ||||
|---|---|---|---|---|---|
| N | P | K | Ca | Mg | |
| Geneva | |||||
| 2015 | 0.71 ± 0.06 | 0.16 ± 0.01 b | 1.04 ± 0.04 b | 0.38 ± 0.04 | 0.10 ± 0.01 |
| 2016 | 0.34 ± 0.11 | 0.18 ± 0.00 b | 1.17 ± 0.05 b | 0.35 ± 0.03 | 0.07 ± 0.01 |
| 2018 | 1.19 ± 0.17 | 0.22 ± 0.01 a | 1.47 ± 0.04 a | 0.39 ± 0.04 | 0.10 ± 0.01 |
| Weiki | |||||
| 2015 | 1.45 ± 0.20 | 0.21 ± 0.03 a | 1.20 ± 0.14 ab | 0.38 ± 0.02 | 0.10 ± 0.01 |
| 2016 | 0.67 ± 0.11 | 0.17 ± 0.01 b | 1.19 ± 0.05 b | 0.24 ± 0.03 | 0.06 ± 0.00 |
| 2018 | 1.52 ± 0.25 | 0.22 ± 0.03 a | 1.41 ± 0.12 a | 0.36 ± 0.03 | 0.10 ± 0.01 |
| Bingo | |||||
| 2015 | 0.90 ± 0.68 | 0.12 ± 0.02 b | 0.93 ± 0.06 | 0.33 ± 0.01 | 0.09 ± 0.01 |
| 2016 | 0.69 ± 0.11 | 0.16 ± 0.01 a | 0.95 ± 0.02 | 0.26 ± 0.04 | 0.06 ± 0.01 |
| 2018 | 0.67 ± 0.11 | 0.17 ± 0.01 a | 0.97 ± 0.12 | 0.30 ± 0.07 | 0.09 ± 0.01 |
| Anna | |||||
| 2015 | 0.97 ± 0.06 | 0.17 ± 0.01 | 1.16 ± 0.04 | 0.35 ± 0.01 | 0.10 ± 0.01 |
| 2016 | 0.67 ± 0.19 | 0.19 ± 0.02 | 1.27 ± 0.13 | 0.27 ± 0.05 | 0.06 ± 0.01 |
| 2018 | 1.15 ± 0.23 | 0.21 ± 0.01 | 1.29 ± 0.17 | 0.29 ± 0.04 | 0.10 ± 0.01 |
| Global cultivar effect (2015, 2016, and 2018) | |||||
| Geneva | 0.75 ± 0.38 B | 0.19 ± 0.3 B | 1.23 ± 0.20 A | 0.37 ± 0.04 A | 0.09 ± 0.02 A |
| Weiki | 1.22 ± 0.44 A | 0.20 ± 0.03 A | 1.27 ± 0.15 A | 0.32 ± 0.07 AB | 0.09 ± 0.02 AB |
| Bingo | 0.75 ± 0.37 B | 0.15 ± 0.02 C | 0.95 ± 0.07 B | 0.30 ± 0.05 B | 0.08 ± 0.02 B |
| Anna | 0.93 ± 0.26 AB | 0.19 ± 0.02 AB | 1.24 ± 0.12 A | 0.30 ± 0.05 B | 0.09 ± 0.02 AB |
| Year effect | |||||
| 2015 | 1.01 ± 0.42 A | 0.17 ± 0.01 B | 1.08 ± 0.13 B | 0.36 ± 0.03 A | 0.10 ± 0.01 A |
| 2016 | 0.59 ± 0.19 B | 0.18 ± 0.01 B | 1.14 ± 0.14 B | 0.28 ± 0.05 B | 0.06 ± 0.01 B |
| 2018 | 1.13 ± 0.36 A | 0.21 ± 0.01 A | 1.29 ± 0.23 A | 0.33 ± 0.06 A | 0.10 ± 0.01 A |
| Interaction | Ns | <0.05 | <0.05 | Ns | Ns |
| Group | Micronutrients (mg/kg DW) | ||||
|---|---|---|---|---|---|
| Fe | Mn | Zn | Cu | B | |
| Geneva | |||||
| 2015 | 24.8 ± 0.9 | 13.3 ± 1.7 | 9.97 ± 1.68 | 5.42 ± 0.51 b | 15.5 ± 0.6 |
| 2016 | 22.0 ± 1.0 | 18.0 ± 3.6 | 5.90 ± 1.21 | 4.97 ± 0.67 b | 15.1 ± 0.5 |
| 2018 | 23.6 ± 1.1 | 15.0 ± 0.7 | 6.93 ± 0.46 | 8.50 ± 1.15 a | 22.2 ± 0.6 |
| Weiki | |||||
| 2015 | 41.8 ± 4.6 | 20.6 ± 1.1 a | 13.27 ± 2.50 | 6.84 ± 0.71 b | 17.4 ± 0.7 |
| 2016 | 35.1 ± 12.3 | 13.6 ± 2.4 b | 7.17 ± 1.18 | 6.03 ± 1.15 b | 13.8 ± 0.7 |
| 2018 | 28.7 ± 2.2 | 10.3 ± 0.1 b | 8.22 ± 0.42 | 9.27 ± 0.98 a | 21.6 ± 0.4 |
| Bingo | |||||
| 2015 | 23.0 ± 0.6 | 9.2 ± 0.4 | 9.12 ± 0.56 | 4.35 ± 0.53 | 16.5 ± 0.6 |
| 2016 | 23.1 ± 4.3 | 9.1 ± 5.3 | 7.87 ± 4.56 | 4.47 ± 1.51 | 14.1 ± 0.7 |
| 2018 | 18.6 ± 0.9 | 8.3 ± 2.7 | 6.38 ± 0.65 | 4.35 ± 0.88 | 21.1 ± 3.0 |
| Anna | |||||
| 2015 | 32.1 ± 2.4 | 22.1 ± 2.0 a | 12.30 ± 0.10 | 5.80 ± 0.45 b | 17.0 ± 0.5 |
| 2016 | 22.9 ± 3.8 | 9.6 ± 5.0 b | 5.33 ± 2.68 | 5.87 ± 1.67 b | 14.0 ± 0.8 |
| 2018 | 25.6 ± 1.7 | 14.2 ± 3.8 b | 8.11 ± 0.96 | 9.97 ± 1.30 a | 22.5 ± 0.6 |
| Global cultivar effect (2015, 2016, and 2018) | |||||
| Geneva | 23.5 ± 1.5 B | 15.4 ± 2.9 A | 7.60 ± 2.12 | 6.30 ± 1.81 A | 17.6 ± 3.5 |
| Weiki | 35.2 ± 8.7 A | 14.8 ± 4.8 A | 9.55 ± 3.15 | 7.38 ± 1.68 A | 17.6 ± 3.4 |
| Bingo | 21.6 ± 3.1 B | 8.8 ± 3.0 B | 7.79 ± 2.61 | 4.39 ± 0.92 B | 17.3 ± 3.5 |
| Anna | 26.9 ± 4.8 B | 15.3 ± 6.4 A | 8.58 ± 3.35 | 7.21 ± 2.33 A | 17.8 ± 3.8 |
| Year effect | |||||
| 2015 | 30.4 ± 8.0 A | 16.3 ± 5.7 A | 11.16 ± 2.19 A | 5.60 ± 1.04 B | 16.6 ± 0.9 B |
| 2016 | 25.8 ± 8.1 AB | 12.6 ± 5.2 B | 6.57 ± 2.59 B | 5.33 ± 1.30 B | 14.3 ± 0.8 C |
| 2018 | 24.1 ± 4.1 B | 11.9 ± 3.5 B | 7.41 ± 0.99 B | 8.02 ± 2.46 A | 21.9 ± 1.5 A |
| Interaction | Ns | <0.01 | Ns | <0.05 | Ns |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Latocha, P.; Łata, B.; Jankowski, P. Variation of Chemical Composition and Antioxidant Properties of Kiwiberry (Actinidia arguta) in a Three-Year Study. Molecules 2023, 28, 455. https://doi.org/10.3390/molecules28010455
Latocha P, Łata B, Jankowski P. Variation of Chemical Composition and Antioxidant Properties of Kiwiberry (Actinidia arguta) in a Three-Year Study. Molecules. 2023; 28(1):455. https://doi.org/10.3390/molecules28010455
Chicago/Turabian StyleLatocha, Piotr, Barbara Łata, and Paweł Jankowski. 2023. "Variation of Chemical Composition and Antioxidant Properties of Kiwiberry (Actinidia arguta) in a Three-Year Study" Molecules 28, no. 1: 455. https://doi.org/10.3390/molecules28010455
APA StyleLatocha, P., Łata, B., & Jankowski, P. (2023). Variation of Chemical Composition and Antioxidant Properties of Kiwiberry (Actinidia arguta) in a Three-Year Study. Molecules, 28(1), 455. https://doi.org/10.3390/molecules28010455