
Scope and Limitations of Exploiting the Ability of the Chemosensitizer NV716 to Enhance the Activity of Tetracycline Derivatives against Pseudomonas aeruginosa
 , ,
, ,  and
and 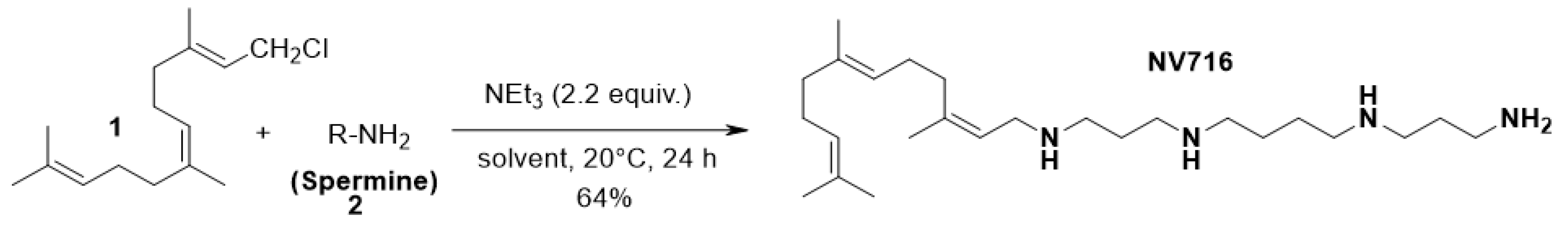{kind=link}
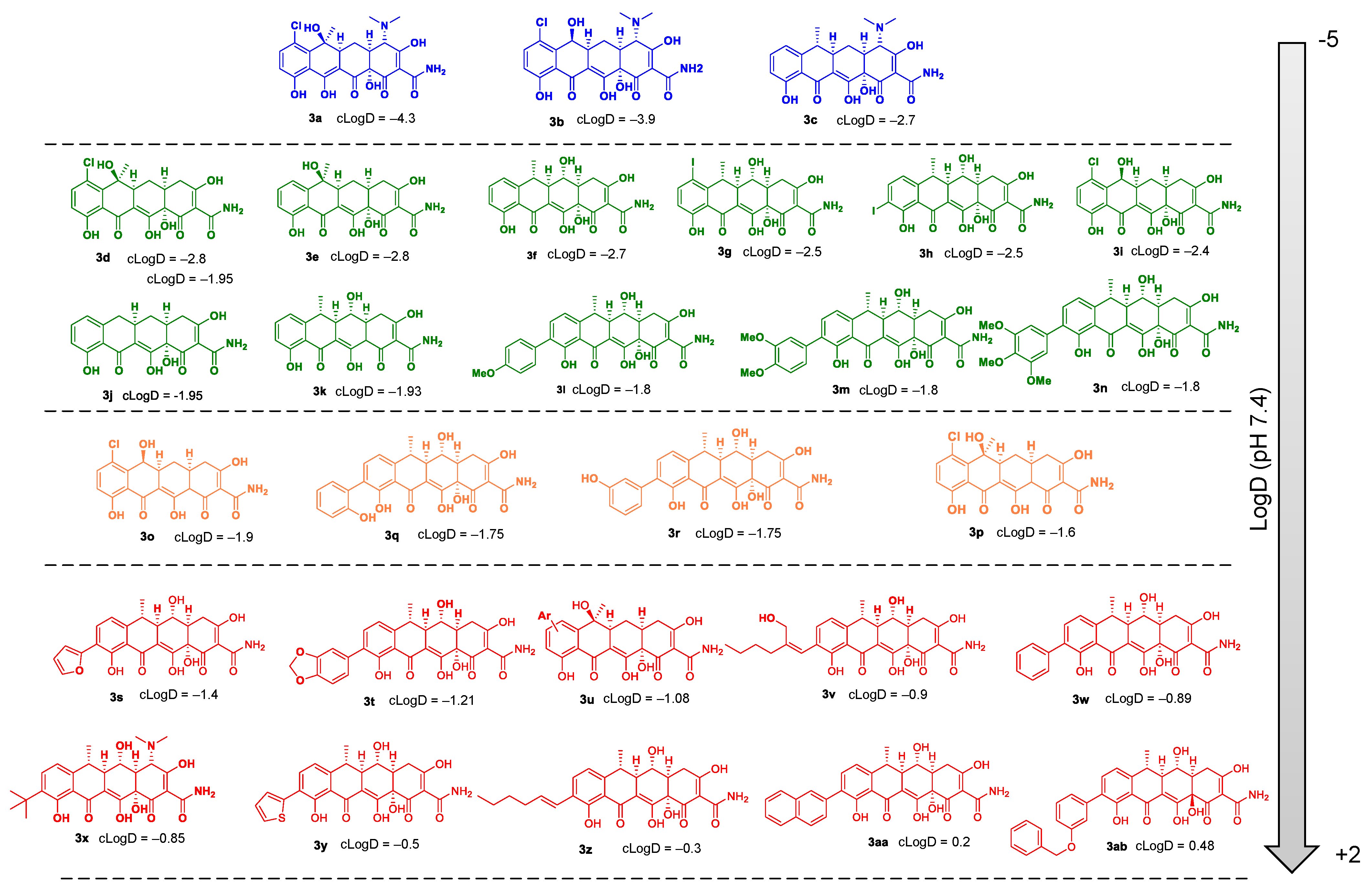{kind=link}
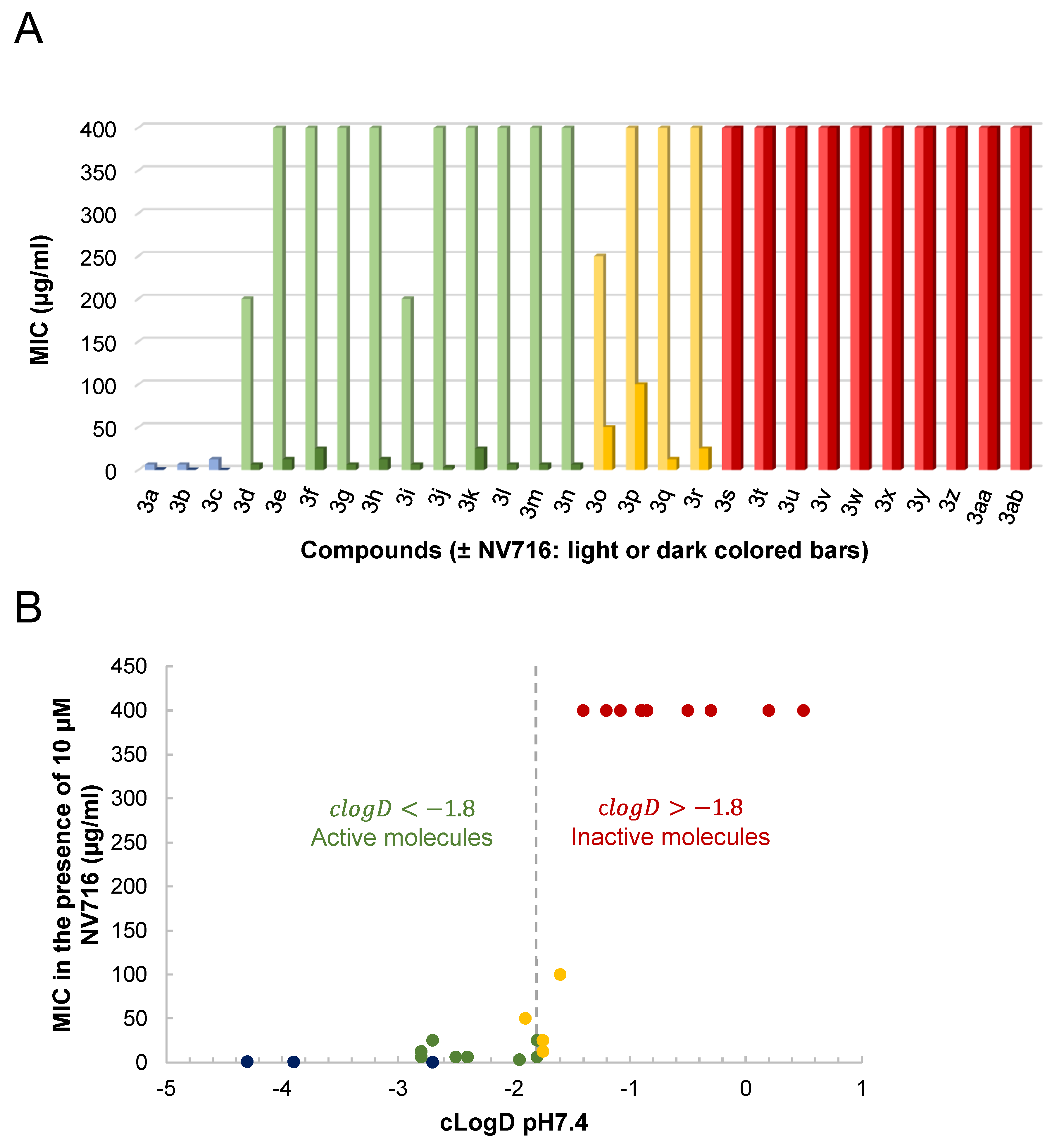{kind=link}
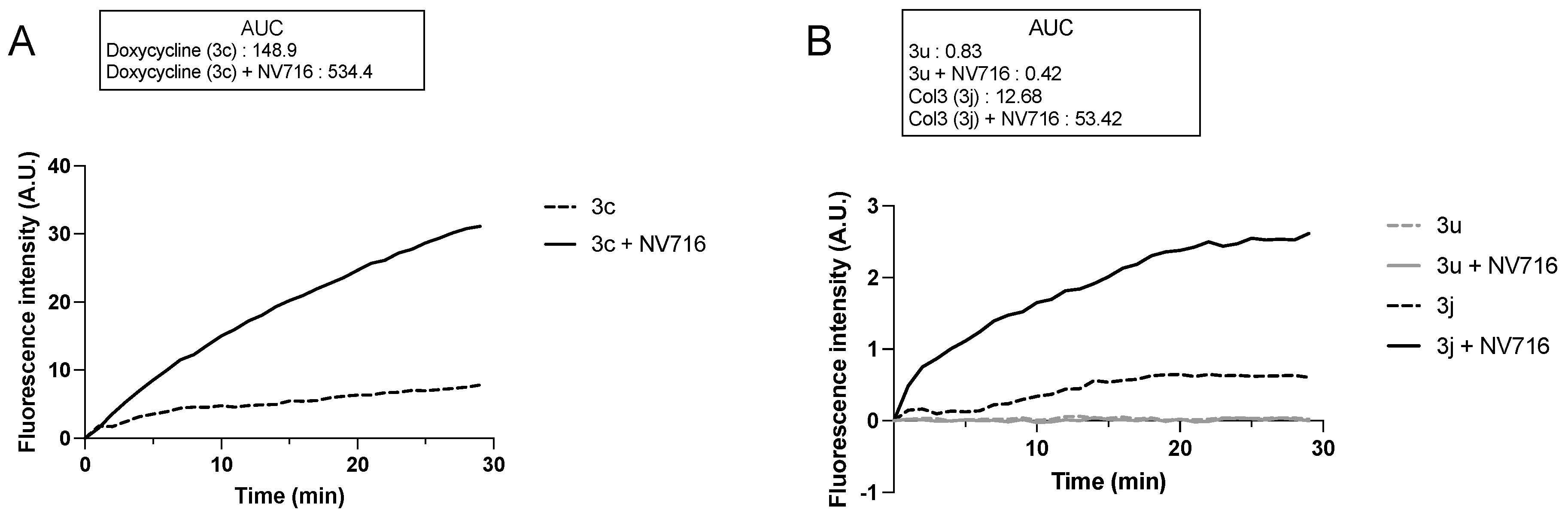{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. OM Perturbation Induced by NV716 Increases the Range of Hydrophobicity of Tetracycline Derivatives Compatible with Antibacterial Activity
2.2. Antibacterial Activity of the Tetracycline Derivatives correlates with Increased Uptake in the Presence of NV716
3. Materials and Methods
3.1. Bacterial Strains and Culture Conditions
3.2. Reagents
3.3. Potentiation Assays
3.4. Physicochemical Property Calculations
3.5. Accumulation Assays
3.6. Bacterial Cell-Free Expression Assays
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
Abbreviations
| A. U. | Arbitrary unit |
| AUC | Area under the curve |
| CLSI | Clinical and Laboratory Standards Institute |
| 1,2′-DNA | 1,2-Dinaphthylamine |
| eNTRy | ionizable Nitrogen, low Three-dimensionality, Rigidity |
| GFP | Green fluorescent protein |
| IC50 | Half maximal inhibitory concentration |
| LPS | Lipopolysaccharides |
| MIC | Minimal inhibitory concentration |
| OM | Outer membrane |
| SD | Standard deviation |
| WHO | World Health Organization |
| XDR | eXtremely Drug Resistant |
References
- Brown, E.D.; Wright, G.D. Antibacterial Drug Discovery in the Resistance Era. Nature 2016, 529, 336–343. [Google Scholar] [CrossRef] [PubMed]
- Shi, Z.; Zhang, J.; Tian, L.; Xin, L.; Liang, C.; Ren, X.; Li, M. A Comprehensive Overview of the Antibiotics Approved in the Last Two Decades: Retrospects and Prospects. Molecules 2023, 28, 1762. [Google Scholar] [CrossRef] [PubMed]
- Theuretzbacher, U.; Bush, K.; Harbarth, S.; Paul, M.; Rex, J.H.; Tacconelli, E.; Thwaites, G.E. Critical Analysis of Antibacterial Agents in Clinical Development. Nat. Rev. Microbiol. 2020, 18, 286–298. [Google Scholar] [CrossRef]
- Antibacterial Agents in Clinical and Preclinical Development: An Overview and Analysis. 2021. Available online: https://www.who.int/publications-detail-redirect/9789240047655 (accessed on 18 March 2023).
- O’Shea, R.; Moser, H.E. Physicochemical Properties of Antibacterial Compounds: Implications for Drug Discovery. J. Med. Chem. 2008, 51, 2871–2878. [Google Scholar] [CrossRef] [PubMed]
- Brown, D.G.; May-Dracka, T.L.; Gagnon, M.M.; Tommasi, R. Trends and Exceptions of Physical Properties on Antibacterial Activity for Gram-Positive and Gram-Negative Pathogens. J. Med. Chem. 2014, 57, 10144–10161. [Google Scholar] [CrossRef] [PubMed]
- Nikaido, H. Molecular Basis of Bacterial Outer Membrane Permeability Revisited. Microbiol. Mol. Biol. Rev. 2003, 67, 593–656. [Google Scholar] [CrossRef] [PubMed]
- Nikaido, H.; Vaara, M. Molecular Basis of Bacterial Outer Membrane Permeability. Microbiol. Rev. 1985, 49, 1–32. [Google Scholar] [CrossRef]
- Vergalli, J.; Bodrenko, I.V.; Masi, M.; Moynié, L.; Acosta-Gutiérrez, S.; Naismith, J.H.; Davin-Regli, A.; Ceccarelli, M.; Van Den Berg, B.; Winterhalter, M.; et al. Porins and Small-Molecule Translocation across the Outer Membrane of Gram-Negative Bacteria. Nat. Rev. Microbiol. 2020, 18, 164–176. [Google Scholar] [CrossRef]
- Kojima, S.; Nikaido, H. Permeation Rates of Penicillins Indicate That Escherichia coli Porins Function Principally as Nonspecific Channels. Proc. Natl. Acad. Sci. USA 2013, 110, E2629–E2634. [Google Scholar] [CrossRef]
- Eren, E.; Vijayaraghavan, J.; Liu, J.; Cheneke, B.R.; Touw, D.S.; Lepore, B.W.; Indic, M.; Movileanu, L.; Van Den Berg, B. Substrate Specificity within a Family of Outer Membrane Carboxylate Channels. PLoS Biol. 2012, 10, e1001242. [Google Scholar] [CrossRef]
- Li, X.-Z.; Plésiat, P.; Nikaido, H. The Challenge of Efflux-Mediated Antibiotic Resistance in Gram-Negative Bacteria. Clin. Microbiol. Rev. 2015, 28, 337–418. [Google Scholar] [CrossRef] [PubMed]
- Krishnamoorthy, G.; Wolloscheck, D.; Weeks, J.W.; Croft, C.; Rybenkov, V.V.; Zgurskaya, H.I. Breaking the Permeability Barrier of Escherichia coli by Controlled Hyperporination of the Outer Membrane. Antimicrob. Agents Chemother. 2016, 60, 7372–7381. [Google Scholar] [CrossRef]
- Krishnamoorthy, G.; Leus, I.V.; Weeks, J.W.; Wolloscheck, D.; Rybenkov, V.V.; Zgurskaya, H.I. Synergy between Active Efflux and Outer Membrane Diffusion Defines Rules of Antibiotic Permeation into Gram-Negative Bacteria. mBio 2017, 8, e01172-17. [Google Scholar] [CrossRef] [PubMed]
- Zgurskaya, H.I.; Rybenkov, V.V. Permeability Barriers of Gram-negative Pathogens. Ann. N. Y. Acad. Sci. 2020, 1459, 5–18. [Google Scholar] [CrossRef] [PubMed]
- Richter, M.F.; Drown, B.S.; Riley, A.P.; Garcia, A.; Shirai, T.; Svec, R.L.; Hergenrother, P.J. Predictive Compound Accumulation Rules Yield a Broad-Spectrum Antibiotic. Nature 2017, 545, 299–304. [Google Scholar] [CrossRef]
- Richter, M.F.; Hergenrother, P.J. The Challenge of Converting Gram-Positive-Only Compounds into Broad-Spectrum Antibiotics: Challenges in Developing Broad-Spectrum Antibiotics. Ann. N. Y. Acad. Sci. 2019, 1435, 18–38. [Google Scholar] [CrossRef]
- Parker, E.N.; Drown, B.S.; Geddes, E.J.; Lee, H.Y.; Ismail, N.; Lau, G.W.; Hergenrother, P.J. Implementation of Permeation Rules Leads to a FabI Inhibitor with Activity against Gram-Negative Pathogens. Nat. Microbiol. 2019, 5, 67–75. [Google Scholar] [CrossRef]
- Parker, E.N.; Cain, B.N.; Hajian, B.; Ulrich, R.J.; Geddes, E.J.; Barkho, S.; Lee, H.Y.; Williams, J.D.; Raynor, M.; Caridha, D.; et al. An Iterative Approach Guides Discovery of the FabI Inhibitor Fabimycin, a Late-Stage Antibiotic Candidate with In Vivo Efficacy against Drug-Resistant Gram-Negative Infections. ACS Cent. Sci. 2022, 8, 1145–1158. [Google Scholar] [CrossRef]
- Vaara, M. Agents That Increase the Permeability of the Outer Membrane. Microbiol. Rev. 1992, 56, 395–411. [Google Scholar] [CrossRef]
- MacNair, C.R.; Tsai, C.N.; Brown, E.D. Creative Targeting of the Gram-negative Outer Membrane in Antibiotic Discovery. Ann. N. Y. Acad. Sci. 2020, 1459, 69–85. [Google Scholar] [CrossRef]
- Trimble, M.J.; MLynárčik, P.; Kolář, M.; Hancock, R.E.W. Polymyxin: Alternative Mechanisms of Action and Resistance. Cold Spring Harb. Perspect. Med. 2016, 6, a025288. [Google Scholar] [CrossRef] [PubMed]
- Vaara, M. Polymyxin Derivatives That Sensitize Gram-Negative Bacteria to Other Antibiotics. Molecules 2019, 24, 249. [Google Scholar] [CrossRef]
- Corbett, D.; Wise, A.; Langley, T.; Skinner, K.; Trimby, E.; Birchall, S.; Dorali, A.; Sandiford, S.; Williams, J.; Warn, P.; et al. Potentiation of Antibiotic Activity by a Novel Cationic Peptide: Potency and Spectrum of Activity of SPR741. Antimicrob. Agents Chemother. 2017, 61, e00200-17. [Google Scholar] [CrossRef] [PubMed]
- Brown, P.; Dawson, M.J. Development of New Polymyxin Derivatives for Multi-Drug Resistant Gram-Negative Infections. J. Antibiot. 2017, 70, 386–394. [Google Scholar] [CrossRef]
- Blanchet, M.; Borselli, D.; Brunel, J.M. Polyamine Derivatives: A Revival of an Old Neglected Scaffold to Fight Resistant Gram-Negative Bacteria? Future Med. Chem. 2016, 8, 963–973. [Google Scholar] [CrossRef] [PubMed]
- Borselli, D.; Lieutaud, A.; Thefenne, H.; Garnotel, E.; Pagès, J.-M.; Brunel, J.M.; Bolla, J.-M. Polyamino-Isoprenic Derivatives Block Intrinsic Resistance of P. aeruginosa to Doxycycline and Chloramphenicol In Vitro. PLoS ONE 2016, 11, e0154490. [Google Scholar] [CrossRef] [PubMed]
- Borselli, D.; Brunel, J.M.; Gorgé, O.; Bolla, J.M. Polyamino-Isoprenyl Derivatives as Antibiotic Adjuvants and Motility Inhibitors for Bordetella bronchiseptica Porcine Pulmonary Infection Treatment. Front. Microbiol. 2019, 10, 1771. [Google Scholar] [CrossRef]
- Wang, G.; Brunel, J.-M.; Rodriguez-Villalobos, H.; Bolla, J.-M.; Van Bambeke, F. The Polyamino-Isoprenyl Potentiator NV716 Revives Disused Antibiotics against Gram-Negative Bacteria in Broth, Infected Monocytes, or Biofilms, by Disturbing the Barrier Effect of Their Outer Membrane. Eur. J. Med. Chem. 2022, 238, 114496. [Google Scholar] [CrossRef]
- Wang, G.; Brunel, J.-M.; Bolla, J.-M.; Van Bambeke, F. The Polyaminoisoprenyl Potentiator NV716 Revives Old Disused Antibiotics against Intracellular Forms of Infection by Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2021, 65, e02028-20. [Google Scholar] [CrossRef]
- Wang, G.; Brunel, J.-M.; Preusse, M.; Mozaheb, N.; Willger, S.D.; Larrouy-Maumus, G.; Baatsen, P.; Häussler, S.; Bolla, J.-M.; Van Bambeke, F. The Membrane-Active Polyaminoisoprenyl Compound NV716 Re-Sensitizes Pseudomonas aeruginosa to Antibiotics and Reduces Bacterial Virulence. Commun. Biol. 2022, 5, 871. [Google Scholar] [CrossRef]
- Fuoco, D. Classification Framework and Chemical Biology of Tetracycline-Structure-Based Drugs. Antibiotics 2012, 1, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Thomas, M.; Le, W.D.; Jankovic, J. Minocycline and Other Tetracycline Derivatives: A Neuroprotective Strategy in Parkinson’s Disease and Huntington’s Disease. Clin. Neuropharmacol. 2003, 26, 18–23. [Google Scholar] [CrossRef]
- Tomas-Grau, R.; González-Lizárraga, F.; Ploper, D.; Avila, C.L.; Socías, S.B.; Besnault, P.; Tourville, A.; Mella, R.M.; Villacé, P.; Salado, C.; et al. Neuroprotective Effects of a Novel Demeclocycline Derivative Lacking Antibiotic Activity: From a Hit to a Promising Lead Compound. Cells 2022, 11, 2759. [Google Scholar] [CrossRef] [PubMed]
- Tourville, A.; Viguier, S.; González-Lizárraga, F.; Tomas-Grau, R.H.; Ramirez, P.; Brunel, J.-M.; Dos Santos Pereira, M.; Del-Bel, E.; Chehin, R.; Ferrié, L.; et al. Rescue of Dopamine Neurons from Iron-Dependent Ferroptosis by Doxycycline and Demeclocycline and Their Non-Antibiotic Derivatives. Antioxidants 2023, 12, 575. [Google Scholar] [CrossRef] [PubMed]
- Lieutaud, A.; Pieri, C.; Bolla, J.M.; Brunel, J.M. New Polyaminoisoprenyl Antibiotics Enhancers against Two Multidrug-Resistant Gram-Negative Bacteria from Enterobacter and Salmonella Species. J. Med. Chem. 2020, 63, 10496–10508. [Google Scholar] [CrossRef]
- Connell, S.R.; Tracz, D.M.; Nierhaus, K.H.; Taylor, D.E. Ribosomal Protection Proteins and Their Mechanism of Tetracycline Resistance. Antimicrob. Agents Chemother. 2003, 47, 3675–3681. [Google Scholar] [CrossRef]
- Brodersen, D.E.; Clemons, W.M.; Carter, A.P.; Morgan-Warren, R.J.; Wimberly, B.T.; Ramakrishnan, V. The Structural Basis for the Action of the Antibiotics Tetracycline, Pactamycin, and Hygromycin B on the 30S Ribosomal Subunit. Cell 2000, 103, 1143–1154. [Google Scholar] [CrossRef]
- Ferreira Junior, N.C.; Dos Santos Pereira, M.; Francis, N.; Ramirez, P.; Martorell, P.; González-Lizarraga, F.; Figadère, B.; Chehin, R.; Del Bel, E.; Raisman-Vozari, R.; et al. The Chemically-Modified Tetracycline COL-3 and Its Parent Compound Doxycycline Prevent Microglial Inflammatory Responses by Reducing Glucose-Mediated Oxidative Stress. Cells 2021, 10, 2163. [Google Scholar] [CrossRef]
- Gasser, V.; Kuhn, L.; Hubert, T.; Aussel, L.; Hammann, P.; Schalk, I.J. The Esterase PfeE, the Achilles’ Heel in the Battle for Iron between Pseudomonas aeruginosa and Escherichia coli. Int. J. Mol. Sci. 2021, 22, 2814. [Google Scholar] [CrossRef]
- Hancock, R.E. Peptide Antibiotics. Lancet 1997, 349, 418–422. [Google Scholar] [CrossRef]
- Stokes, J.M.; MacNair, C.R.; Ilyas, B.; French, S.; Côté, J.-P.; Bouwman, C.; Farha, M.A.; Sieron, A.O.; Whitfield, C.; Coombes, B.K.; et al. Pentamidine Sensitizes Gram-Negative Pathogens to Antibiotics and Overcomes Acquired Colistin Resistance. Nat. Microbiol. 2017, 2, 17028. [Google Scholar] [CrossRef]
- Hancock, R.E.; Wong, P.G. Compounds Which Increase the Permeability of the Pseudomonas aeruginosa Outer Membrane. Antimicrob. Agents Chemother. 1984, 26, 48–52. [Google Scholar] [CrossRef] [PubMed]
- Klobucar, K.; French, S.; Côté, J.-P.; Howes, J.R.; Brown, E.D. Genetic and Chemical-Genetic Interactions Map Biogenesis and Permeability Determinants of the Outer Membrane of Escherichia coli. mBio 2020, 11, e00161-20. [Google Scholar] [CrossRef] [PubMed]
- Theuretzbacher, U.; Outterson, K.; Engel, A.; Karlén, A. The Global Preclinical Antibacterial Pipeline. Nat. Rev. Microbiol. 2020, 18, 275–285. [Google Scholar] [CrossRef] [PubMed]
- MacNair, C.R.; Brown, E.D. Outer Membrane Disruption Overcomes Intrinsic, Acquired, and Spontaneous Antibiotic Resistance. mBio 2020, 11, e01615-20. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Draveny, M.; Rose, C.; Pinet, A.; Ferrié, L.; Figadère, B.; Brunel, J.-M.; Masi, M. Scope and Limitations of Exploiting the Ability of the Chemosensitizer NV716 to Enhance the Activity of Tetracycline Derivatives against Pseudomonas aeruginosa. Molecules 2023, 28, 4262. https://doi.org/10.3390/molecules28114262
Draveny M, Rose C, Pinet A, Ferrié L, Figadère B, Brunel J-M, Masi M. Scope and Limitations of Exploiting the Ability of the Chemosensitizer NV716 to Enhance the Activity of Tetracycline Derivatives against Pseudomonas aeruginosa. Molecules. 2023; 28(11):4262. https://doi.org/10.3390/molecules28114262
Chicago/Turabian StyleDraveny, Margot, Clémence Rose, Alexis Pinet, Laurent Ferrié, Bruno Figadère, Jean-Michel Brunel, and Muriel Masi. 2023. "Scope and Limitations of Exploiting the Ability of the Chemosensitizer NV716 to Enhance the Activity of Tetracycline Derivatives against Pseudomonas aeruginosa" Molecules 28, no. 11: 4262. https://doi.org/10.3390/molecules28114262
APA StyleDraveny, M., Rose, C., Pinet, A., Ferrié, L., Figadère, B., Brunel, J. -M., & Masi, M. (2023). Scope and Limitations of Exploiting the Ability of the Chemosensitizer NV716 to Enhance the Activity of Tetracycline Derivatives against Pseudomonas aeruginosa. Molecules, 28(11), 4262. https://doi.org/10.3390/molecules28114262