

Tandem LC-MS Identification of Antitubercular Compounds in Zones of Growth Inhibition Produced by South African Filamentous Actinobacteria


Abstract
:1. Introduction
2. Results and Discussion
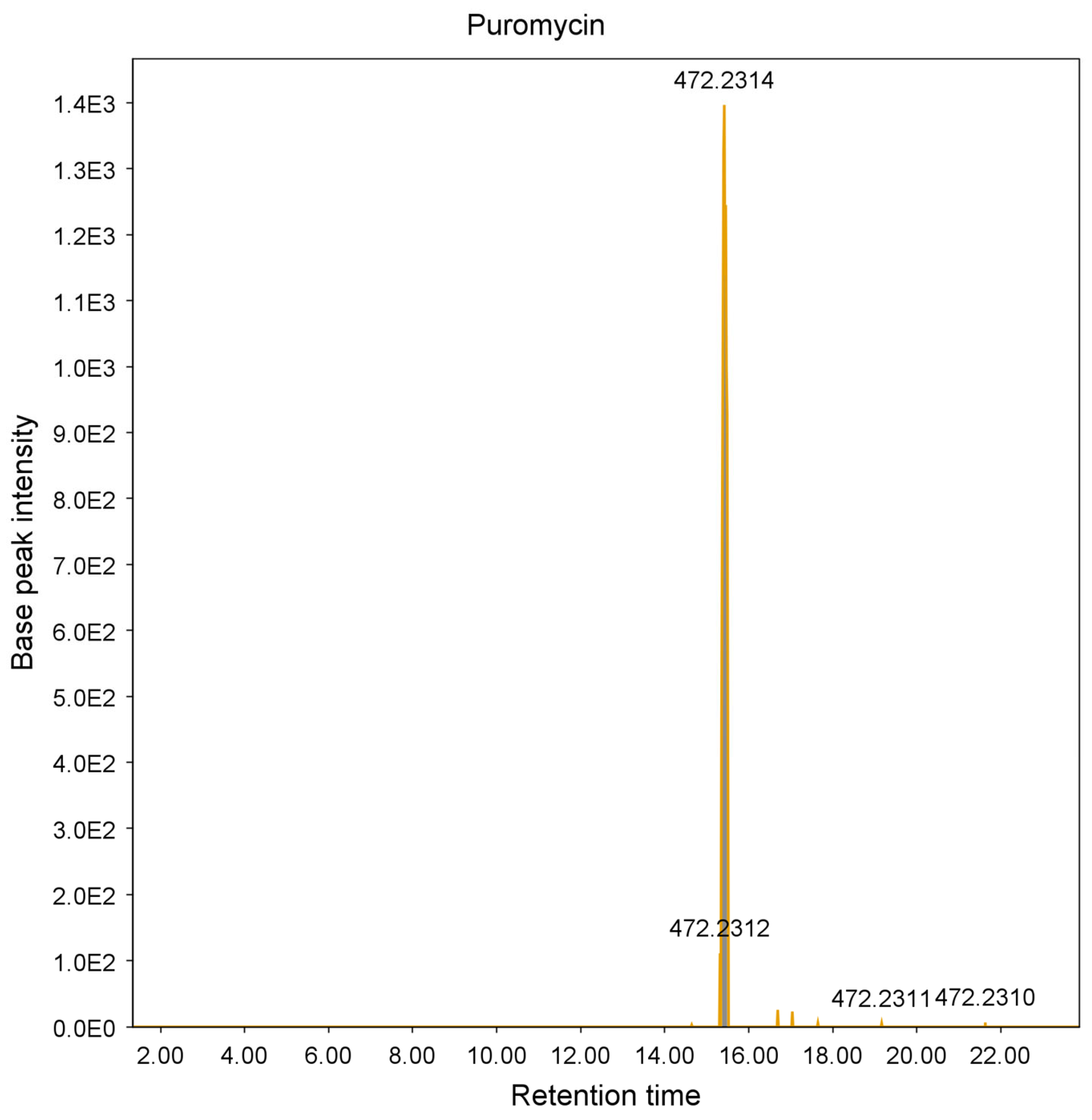
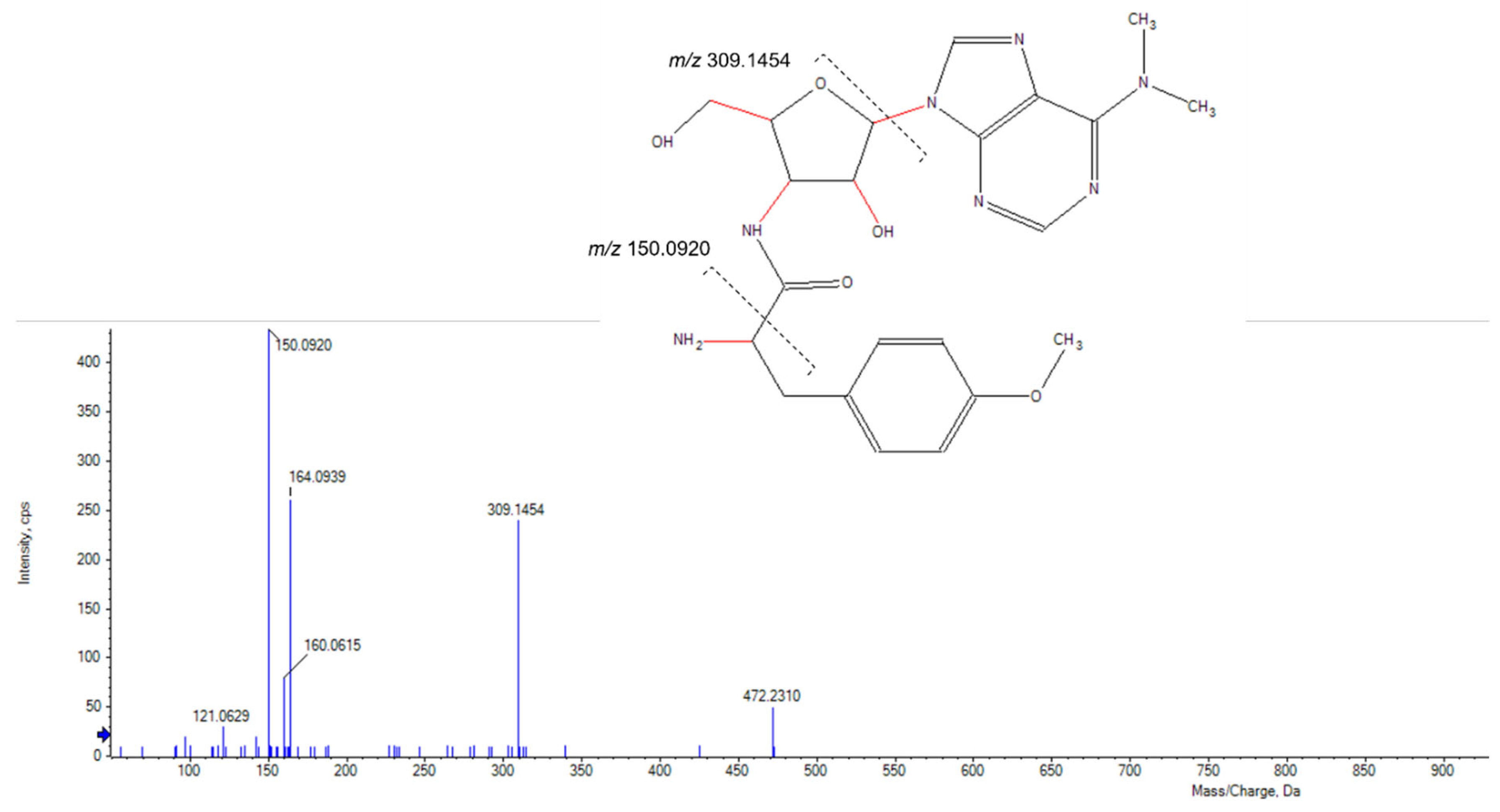
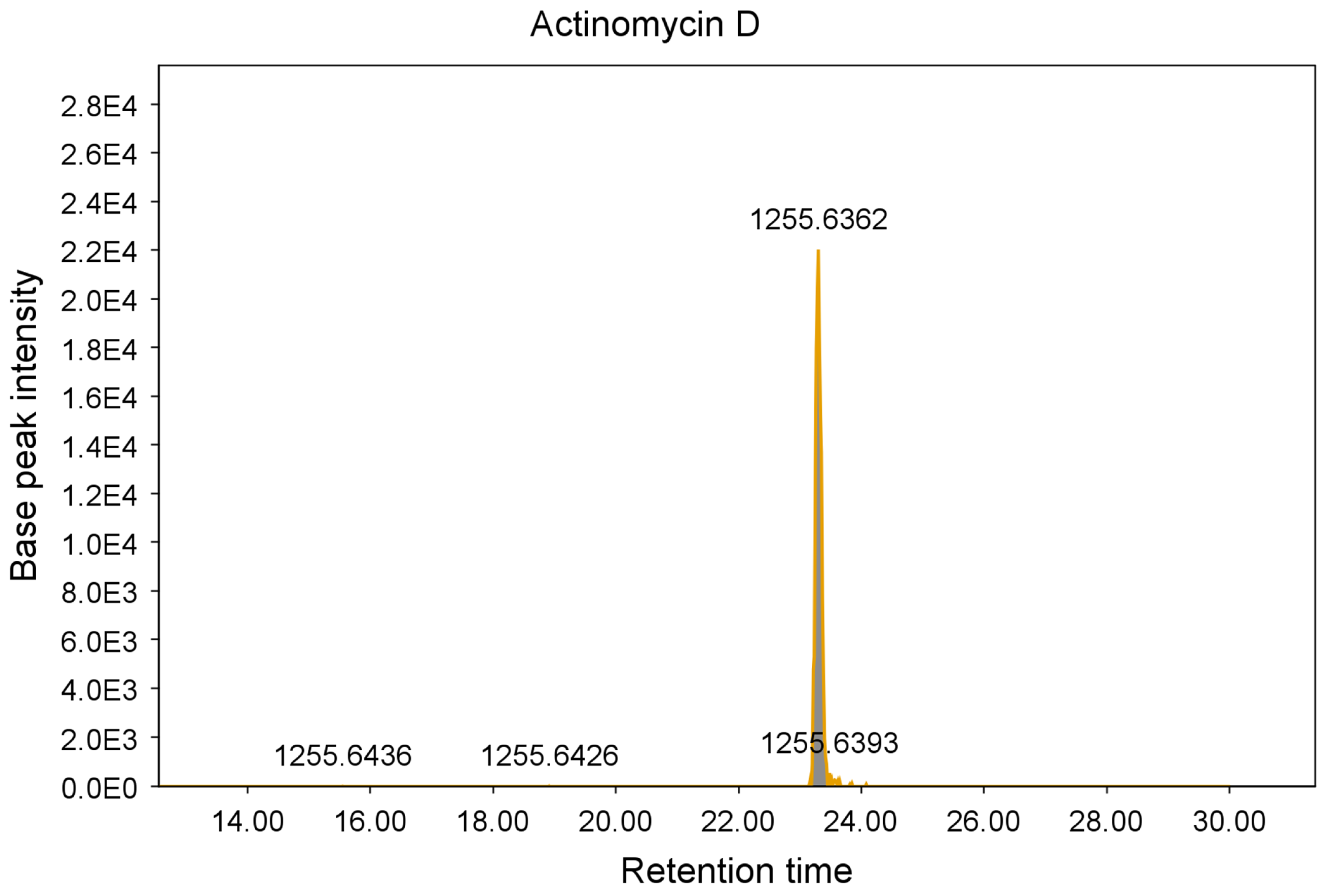
2.1. Agar Overlay Screening against Mycolicibacterium aurum
2.2. Mycobacterium tuberculosis In Vitro Screening
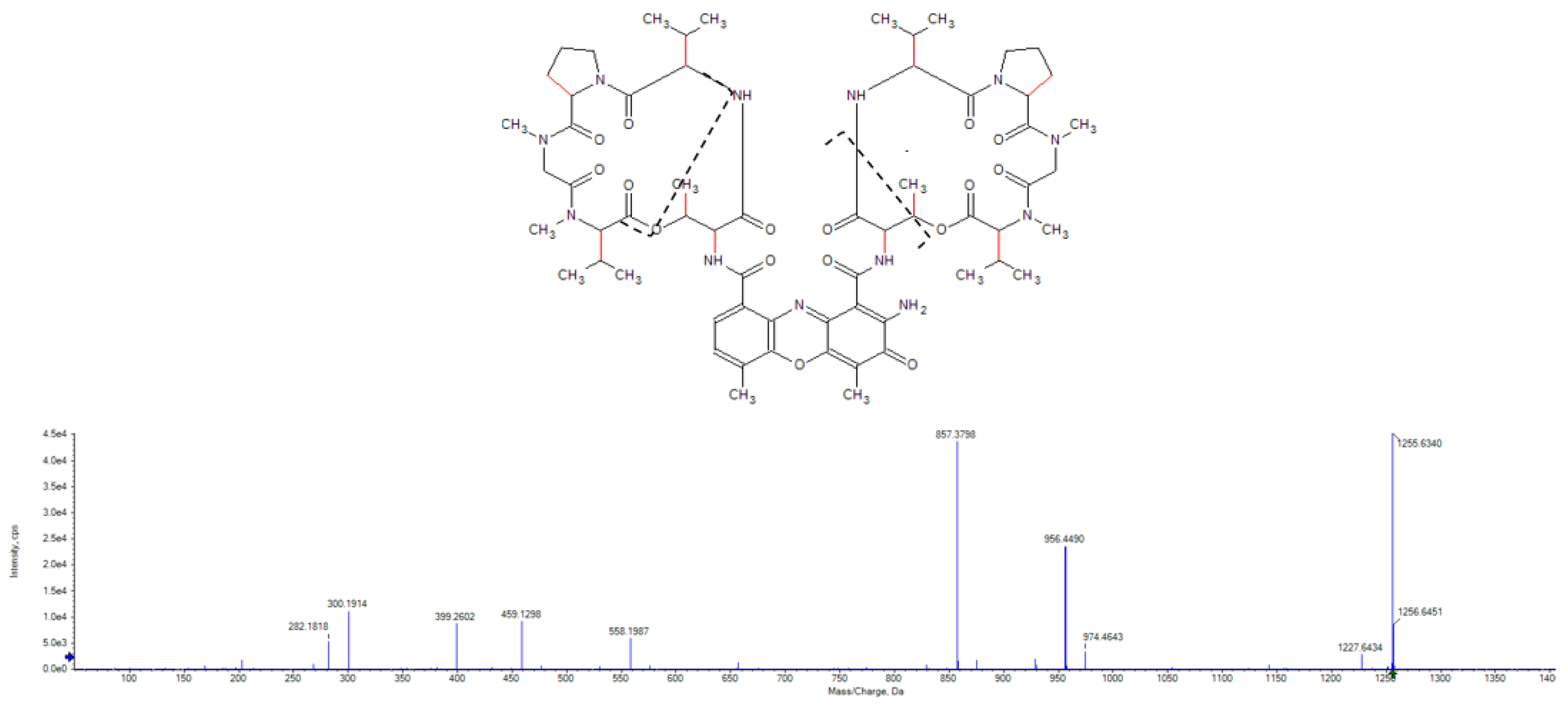
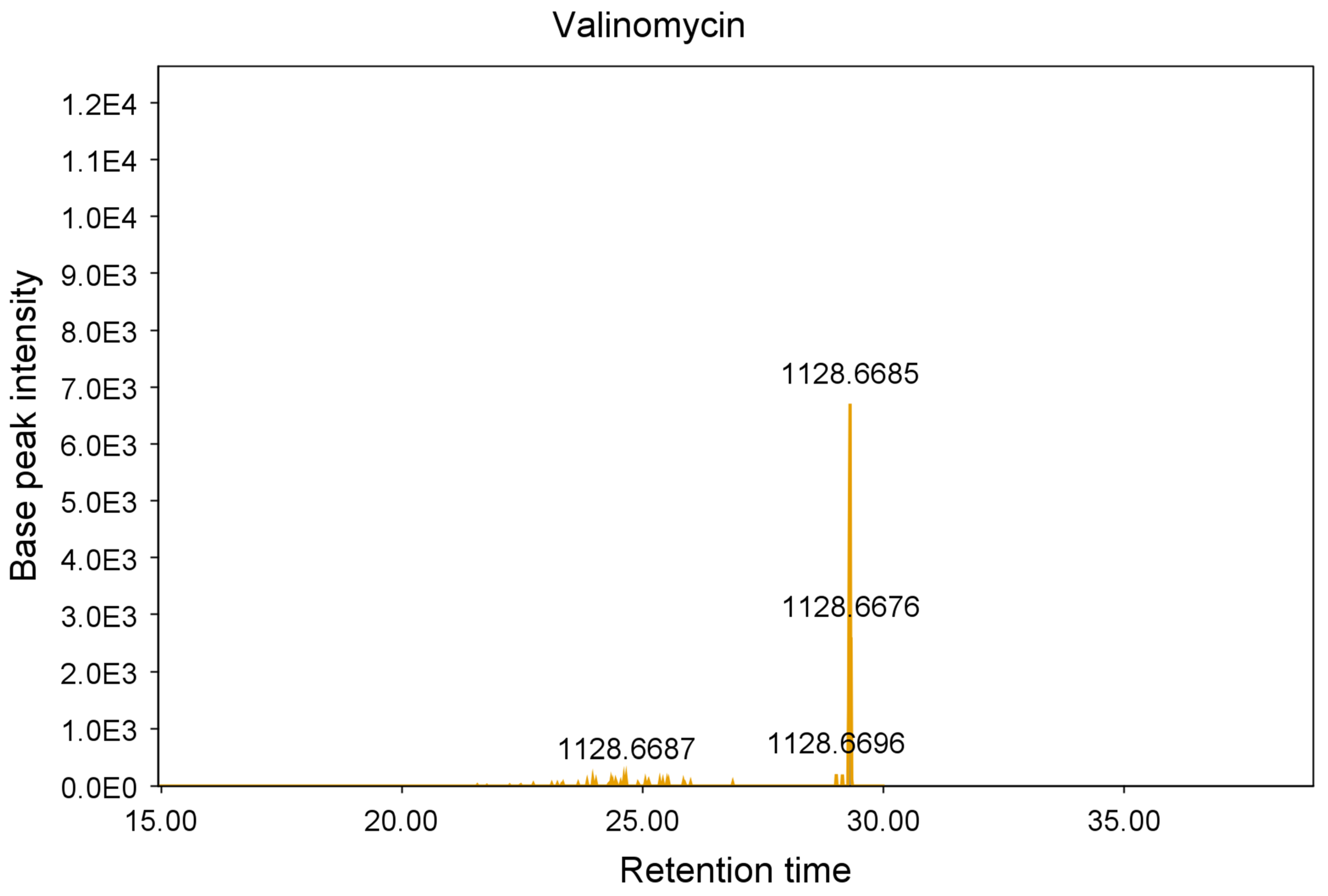
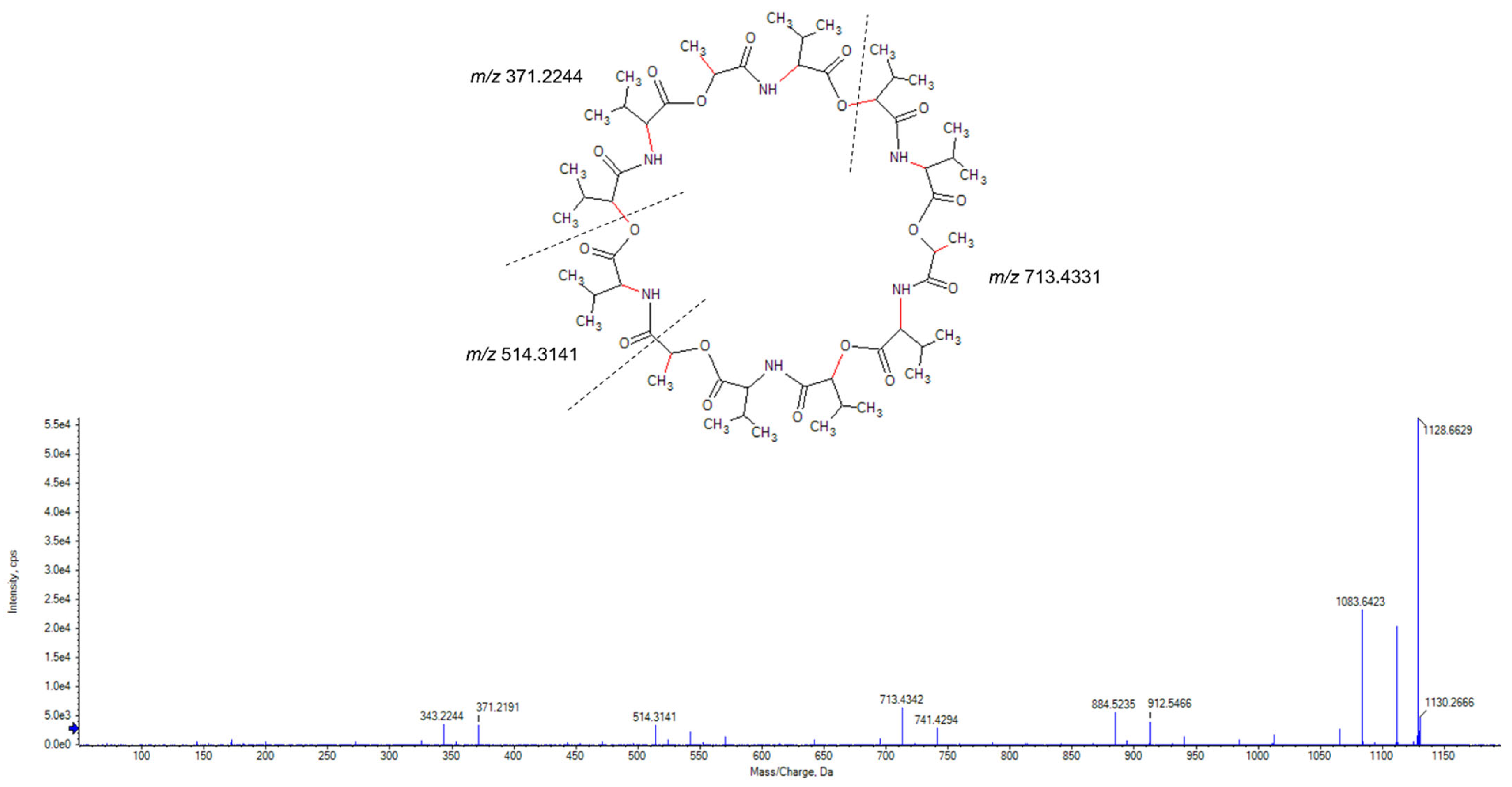
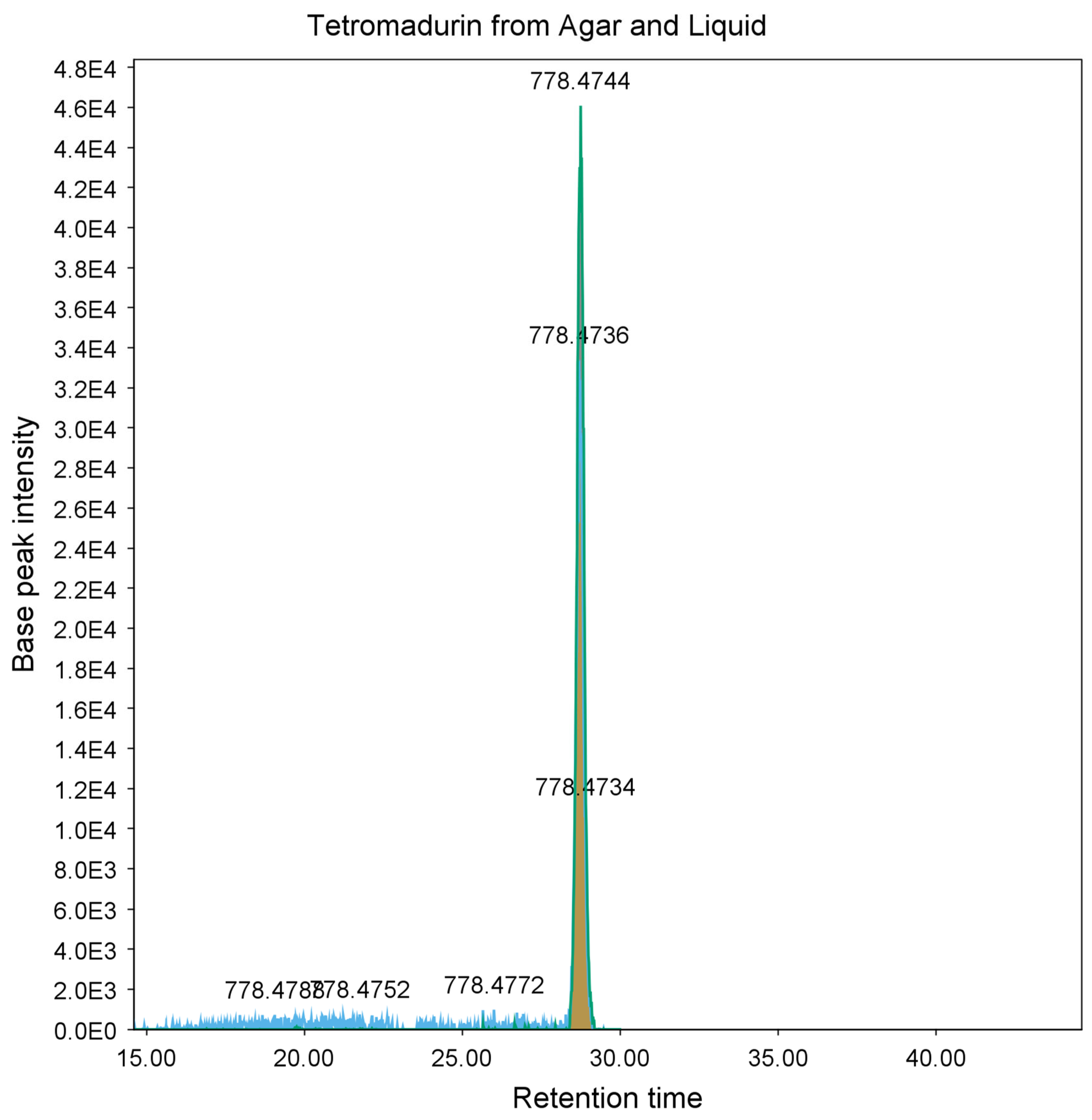
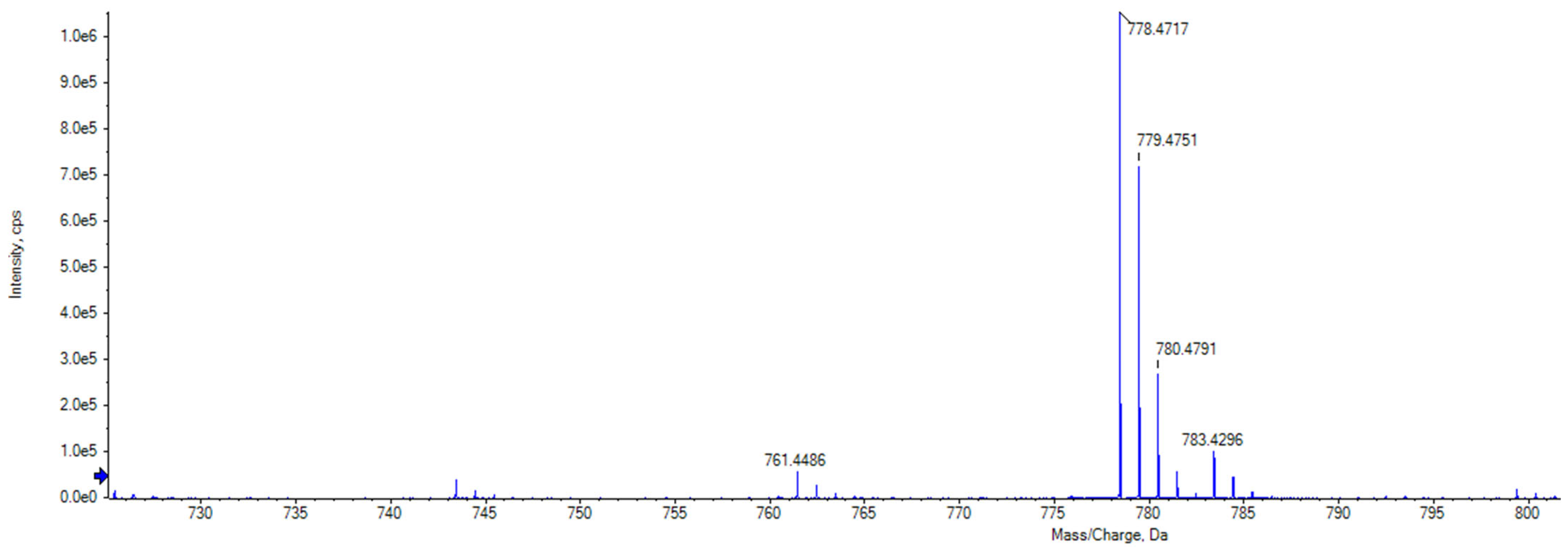
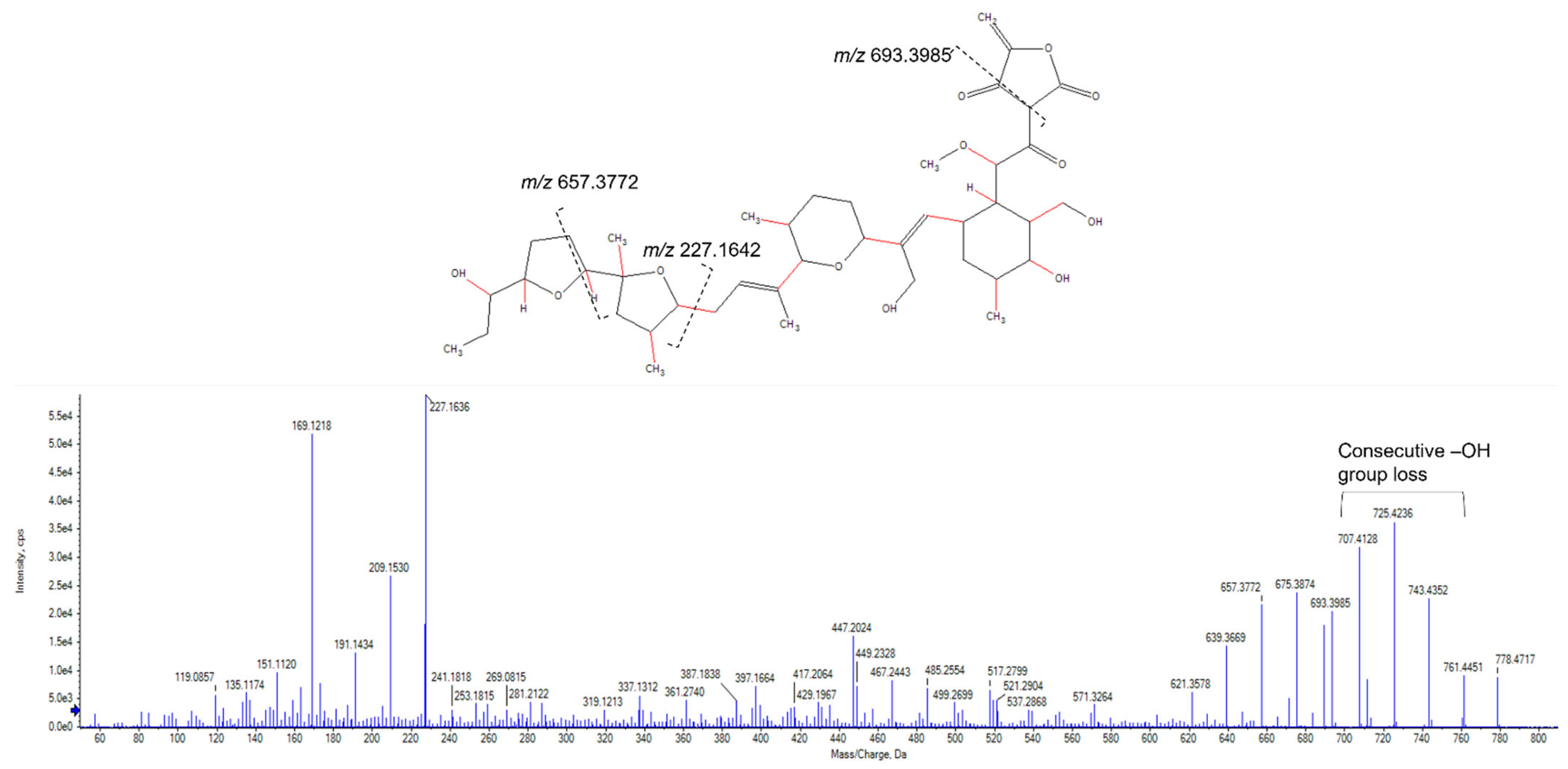
2.3. Purification and Identification of Tetromadurin
3. Conclusions
4. Materials and Methods
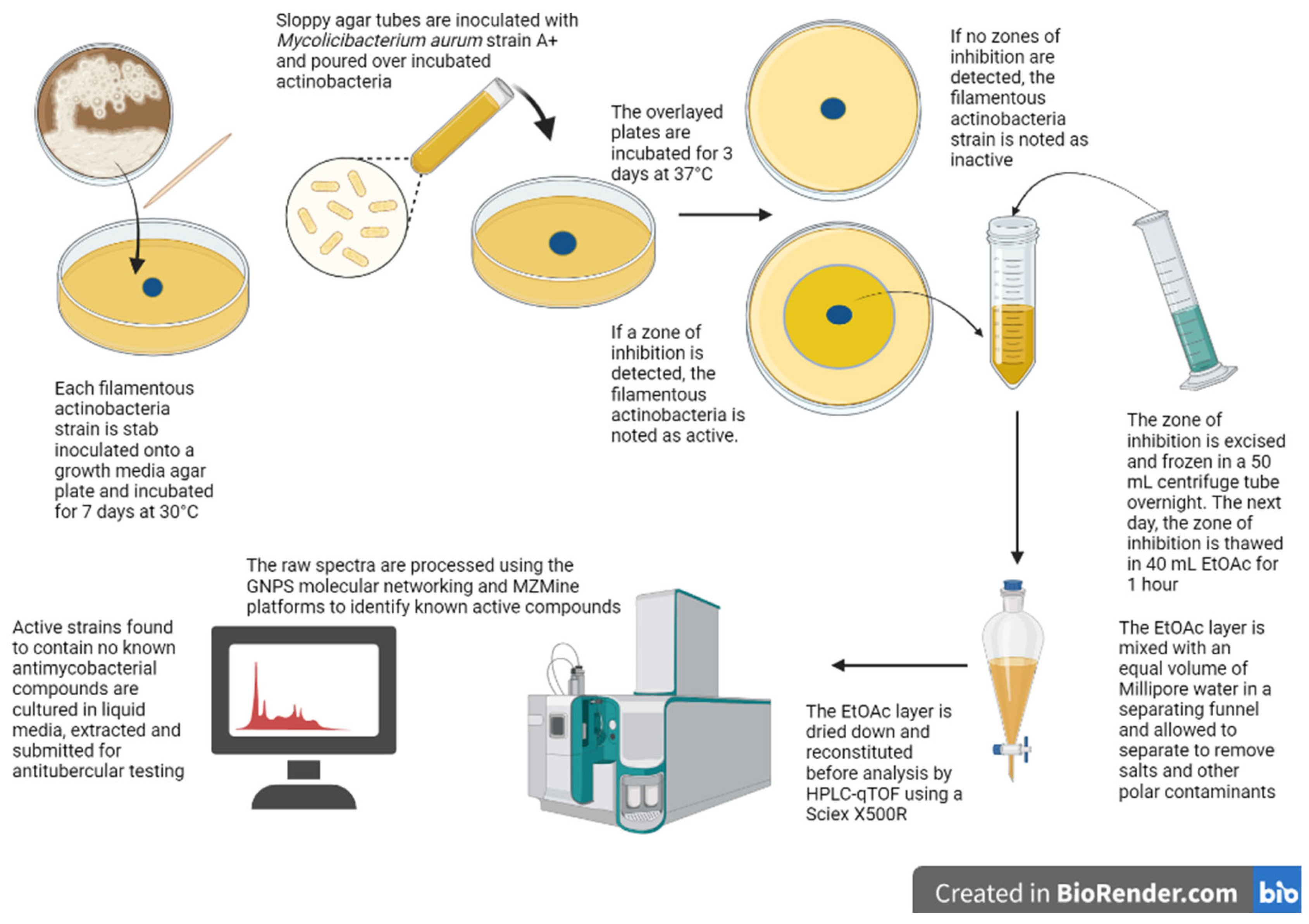
4.1. Agar Overlay Assay and Zone of Inhibition Extraction
4.2. Bacterial Cell Cultivation and Extraction
4.3. Antitubercular Testing
4.4. Cytotoxicity Testing
4.5. Large-Scale Cultivation
4.6. HPLC-MS Analysis and Molecular Networking
4.7. NMR Structural Assignment of Tetromadurin (SF2487/A80577)
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Global Tuberculosis Report 2022; World Health Organization: Geneva, Switzerland, 2022. [Google Scholar]
- Jang, J.G.; Chung, J.H. Diagnosis and treatment of multidrug-resistant tuberculosis. Yeungnam Univ. J. Med. 2020, 37, 277–285. Available online: http://e-yujm.org/journal/view.php?doi=10.12701/yujm.2020.00626 (accessed on 8 December 2022). [CrossRef] [PubMed]
- Grzelak, E.M.; Choules, M.P.; Gao, W.; Cai, G.; Wan, B.; Wang, Y.; McAlpine, J.B.; Cheng, J.; Jin, Y.; Lee, H.; et al. Strategies in anti-Mycobacterium tuberculosis drug discovery based on phenotypic screening. J. Antibiot. 2019, 72, 719–728. Available online: http://www.nature.com/articles/s41429-019-0205-9 (accessed on 22 March 2020). [CrossRef] [PubMed]
- Schatz, A.; Bugie, E.; Waksman, S.A.; Hanssen, A.D.; Patel, R.; Osmon, D.R. The Classic: Streptomycin, a Substance Exhibiting Antibiotic Activity against Gram-Positive and Gram-Negative Bacteria. Clin. Orthop. Relat. Res. 1944, 55, 66–69. Available online: http://gateway.ovid.com/ovidweb.cgi?T=JS&PAGE=crossref&AN=00003086-200508000-00002 (accessed on 26 March 2020).
- Saxena, A.; Kumari, R.; Mukherjee, U.; Singh, P.; Lal, R. Draft Genome Sequence of the Rifamycin Producer Amycolatopsis rifamycinica DSM 46095. Genome Announc. 2014, 2, e00662-14. Available online: http://genomea.asm.org/cgi/doi/10.1128/genomeA.00662-14 (accessed on 26 March 2020). [CrossRef] [PubMed]
- Nothias, L.-F.; Nothias-Esposito, M.; da Silva, R.; Wang, M.; Protsyuk, I.; Zhang, Z.; Sarvepalli, A.; Leyssen, P.; Touboul, D.; Costa, J.; et al. Bioactivity-Based Molecular Networking for the Discovery of Drug Leads in Natural Product Bioassay-Guided Fractionation. J. Nat. Prod. 2018, 81, 758–767. Available online: https://pubs.acs.org/doi/10.1021/acs.jnatprod.7b00737 (accessed on 26 March 2020). [CrossRef]
- Wang, M.; Carver, J.J.; Phelan, V.V.; Sanchez, L.M.; Garg, N.; Peng, Y.; Nguyen, D.D.; Watrous, J.; Kapono, C.A.; Luzzatto-Knaan, T.; et al. Sharing and community curation of mass spectrometry data with Global Natural Products Social Molecular Networking. Nat. Biotechnol. 2016, 34, 828–837. Available online: http://www.nature.com/articles/nbt.3597 (accessed on 7 November 2019). [CrossRef] [PubMed]
- Hoffmann, T.; Krug, D.; Bozkurt, N.; Duddela, S.; Jansen, R.; Garcia, R.; Gerth, K.; Steinmetz, H. Correlating chemical diversity with taxonomic distance for discovery of natural products in myxobacteria. Nat. Commun. 2018, 9, 803. Available online: http://www.nature.com/articles/s41467-018-03184-1 (accessed on 8 December 2022). [CrossRef]
- Nett, M.; Ikeda, H.; Moore, B.S. Genomic basis for natural product biosynthetic diversity in the actinomycetes. Nat. Prod. Rep. 2009, 26, 1362. Available online: http://xlink.rsc.org/?DOI=b817069j (accessed on 8 December 2022). [CrossRef] [PubMed]
- Bérdy, J. Bioactive Microbial Metabolites. J. Antibiot. 2005, 58, 1–26. Available online: http://www.nature.com/articles/ja20051 (accessed on 19 September 2019). [CrossRef] [PubMed]
- Subramani, R.; Aalbersberg, W. Culturable rare Actinomycetes: Diversity, isolation and marine natural product discovery. Appl. Microbiol. Biotechnol. 2013, 97, 9291–9321. [Google Scholar] [CrossRef]
- Cadman, M.; Petersen, C.; Driver, A.; Sekrhan, N.; Maze, K.; Munzhedzi, S. Biodiversity for Development: South Africa’s Landscape Approach to Conserving Biodiversity and Promoting Ecosystem Resilience; South African National Biodiversity Institute: Cape Town, South Africa, 2010. [Google Scholar]
- Everest, G.J.; le Roes-Hill, M.; Rohland, J.; Enslin, S.; Meyers, P.R. Amycolatopsis roodepoortensis sp. nov. and Amycolatopsis speibonae sp. nov.: Antibiotic-producing actinobacteria isolated from South African soils. J. Antibiot. 2014, 67, 813–818. Available online: http://www.nature.com/articles/ja201479 (accessed on 8 December 2022). [CrossRef] [PubMed]
- Curtis, S.M.; Norton, I.; Everest, G.J.; Meyers, P.R. Kribbella podocarpi sp. nov., isolated from the leaves of a yellowwood tree (Podocarpus latifolius). Antonie Van Leeuwenhoek 2018, 111, 875–882. Available online: http://link.springer.com/10.1007/s10482-017-0984-6 (accessed on 8 December 2022). [CrossRef] [PubMed]
- Meyers, P.R.; Porter, D.S.; Omorogie, C.; Pule, J.M. Streptomyces speibonae sp. Nov., a novel streptomycete with blue substrate mycelium isolated from South African soil. Int. J. Syst. Evol. Microbiol. 2003, 53, 801–805. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.B.; Jayasuriya, H.; Ondeyka, J.G.; Herath, K.B.; Zhang, C.; Zink, D.L.; Tsou, N.N.; Ball, R.G.; Basilio, A.; Genilloud, O.; et al. Isolation, Structure, and Absolute Stereochemistry of Platensimycin, A Broad Spectrum Antibiotic Discovered Using an Antisense Differential Sensitivity Strategy. J. Am. Chem. Soc. 2006, 128, 11916–11920. Available online: https://pubs.acs.org/doi/10.1021/ja062232p (accessed on 8 December 2022). [CrossRef]
- Kim, K.H.; Ramadhar, T.R.; Beemelmanns, C.; Cao, S.; Poulsen, M.; Currie, C.R.; Clardy, J. Natalamycin A, an ansamycin from a termite-associated Streptomyces sp. Chem. Sci. 2014, 5, 4333–4338. Available online: http://xlink.rsc.org/?DOI=C4SC01136H (accessed on 15 October 2019). [CrossRef]
- Wood, S.A.; Kirby, B.M.; Goodwin, C.M.; Le Roes, M.; Meyers, P.R. PCR screening reveals unexpected antibiotic biosynthetic potential in Amycolatopsis sp. strain UM16. J. Appl. Microbiol. 2007, 102, 245–253. Available online: http://doi.wiley.com/10.1111/j.1365-2672.2006.03043.x (accessed on 2 April 2020). [CrossRef]
- Namouchi, A.; Cimino, M.; Favre-Rochex, S.; Charles, P.; Gicquel, B. Phenotypic and genomic comparison of Mycobacterium aurum and surrogate model species to Mycobacterium tuberculosis: Implications for drug discovery. BMC Genom. 2017, 18, 530. Available online: http://bmcgenomics.biomedcentral.com/articles/10.1186/s12864-017-3924-y (accessed on 26 March 2020). [CrossRef]
- Bode, H.B.; Bethe, B.; Höfs, R.; Zeeck, A. Big effects from small changes: Possible ways to explore nature’s chemical diversity. Chembiochem 2002, 3, 619–627. Available online: http://www.ncbi.nlm.nih.gov/pubmed/12324995 (accessed on 26 March 2020). [CrossRef]
- Boya, P.C.A.; Fernández-Marín, H.; Mejía, L.C.; Spadafora, C.; Dorrestein, P.C.; Gutiérrez, M. Imaging mass spectrometry and MS/MS molecular networking reveals chemical interactions among cuticular bacteria and pathogenic fungi associated with fungus-growing ants. Sci. Rep. 2017, 7, 5604. Available online: http://www.nature.com/articles/s41598-017-05515-6 (accessed on 24 November 2022). [CrossRef]
- Chambers, M.C.; Maclean, B.; Burke, R.; Amodei, D.; Ruderman, D.L.; Neumann, S.; Gatto, L.; Fischer, B.; Pratt, B.; Egertson, J.; et al. A cross-platform toolkit for mass spectrometry and proteomics. Nat. Biotechnol. 2012, 30, 918–920. Available online: http://www.nature.com/articles/nbt.2377 (accessed on 11 November 2019). [CrossRef]
- Aviner, R. The science of puromycin: From studies of ribosome function to applications in biotechnology. Comput. Struct. Biotechnol. J. 2020, 18, 1074–1083. Available online: http://www.ncbi.nlm.nih.gov/pubmed/32435426 (accessed on 12 October 2022). [CrossRef]
- Fong, C.W. Physiology of ionophore transport of potassium and sodium ions across cell membranes: Valinomycin and 18-crown-6 ether. Int. J. Comput. Biol. Drug Des. 2016, 9, 228. Available online: http://www.inderscience.com/link.php?id=78284 (accessed on 13 November 2019). [CrossRef]
- Qureshi, K.A.; Bholay, A.D.; Rai, P.K.; Mohammed, H.A.; Khan, R.A.; Azam, F.; Jaremko, M.; Emwas, A.-H.; Stefanowicz, P.; Waliczek, M.; et al. Isolation, characterization, anti-MRSA evaluation, and in-silico multi-target anti-microbial validations of actinomycin X2 and actinomycin D produced by novel Streptomyces smyrnaeus UKAQ_23. Sci. Rep. 2021, 11, 14539. Available online: http://www.nature.com/articles/s41598-021-93285-7 (accessed on 12 October 2022). [CrossRef] [PubMed]
- Pink, R.; Hudson, A.; Mouriès, M.; Bendig, M. Opportunities and Challenges in Antiparasitic Drug Discovery. Nat. Rev. Drug Discov. 2005, 4, 727–741. Available online: https://www.nature.com/articles/nrd1824 (accessed on 14 November 2018). [CrossRef] [PubMed]
- Wiley Science Solutions (2017) Antibase: The Natural Compound Identifier. 2017. Available online: https://sciencesolutions.wiley.com/wp-content/uploads/2020/06/Wiley_AntiBase_-The-Natural-Compound-Identifier_978-3-527-34359-1.pdf (accessed on 25 November 2022).
- Little, R.F.; Samborskyy, M.; Leadlay, P.F. The biosynthetic pathway to tetromadurin (SF2487/A80577), a polyether tetronate antibiotic. PLoS ONE 2020, 15, e0239054. Available online: http://www.ncbi.nlm.nih.gov/pubmed/32925967 (accessed on 25 November 2022). [CrossRef] [PubMed]
- Hatsu, M.; Sasaki, T.; Miyadoh, S.; Watabe, H.-O.; Takeuchi, Y.; Kodama, Y.; Orikasa, Y.; Kajii, K.; Shomura, T.; Yamamoto, H.; et al. SF2487, a new polyether antibiotic produced by Actinomadura. J. Antibiot. 1990, 43, 259–266. Available online: http://joi.jlc.jst.go.jp/JST.Journalarchive/antibiotics1968/43.259?from=CrossRef (accessed on 25 November 2022). [CrossRef] [PubMed]
- Adovelande, J.; Schrével, J. Carboxylic ionophores in malaria chemotherapy: The effects of monensin and nigericin on Plasmodium falciparum in vitro and Plasmodium vinckei petteri in vivo. Life Sci. 1996, 59, PL309–PL315. Available online: https://linkinghub.elsevier.com/retrieve/pii/S0024320596005140 (accessed on 16 October 2020). [CrossRef]
- Bharti, H.; Singal, A.; Raza, M.; Ghosh, P.C.; Nag, A. Ionophores as Potent Anti-malarials: A Miracle in the Making. Curr. Top. Med. Chem. 2019, 18, 2029–2041. Available online: http://www.eurekaselect.com/167891/article (accessed on 14 October 2020). [CrossRef]
- Taketuchi, T.; Wataya, Y.; Iinuma, M.; Kim, H.-S.; Watabe, H.; Naganawa, H. Compositions for Treating or Preventing Malaria and Method of Treating Malaria. 2000. Available online: https://patents.google.com/patent/US6939892B2/en (accessed on 25 November 2022).
- Ryoo, I.-J.; Park, H.-R.; Choo, S.-J.; Hwang, J.-H.; Park, Y.-M.; Bae, K.-H.; Shin-Ya, K.; Yoo, I.-D. Selective cytotoxic activity of valinomycin against HT-29 Human colon carcinoma cells via down-regulation of GRP78. Biol. Pharm. Bull. 2006, 29, 817–820. Available online: http://www.ncbi.nlm.nih.gov/pubmed/16595926 (accessed on 12 November 2019). [CrossRef]
- Kaushik, V.; Yakisich, J.; Kumar, A.; Azad, N.; Iyer, A. Ionophores: Potential Use as Anticancer Drugs and Chemosensitizers. Cancers 2018, 10, 360. Available online: https://www.mdpi.com/2072-6694/10/10/360 (accessed on 20 October 2020). [CrossRef]
- An, J.S.; Shin, B.; Kim, T.H.; Hwang, S.; Shin, Y.-H.; Cui, J.; Du, Y.E.; Yi, J.; Nam, S.-J.; Hong, S.; et al. Dumulmycin, an Antitubercular Bicyclic Macrolide from a Riverine Sediment-Derived Streptomyces sp. Org. Lett. 2021, 23, 3359–3363. Available online: https://pubs.acs.org/doi/10.1021/acs.orglett.1c00847 (accessed on 2 April 2020). [CrossRef]
- Oppong-Danquah, E.; Parrot, D.; Blümel, M.; Labes, A.; Tasdemir, D. Molecular Networking-Based Metabolome and Bioactivity Analyses of Marine-Adapted Fungi Co-cultivated With Phytopathogens. Front. Microbiol. 2018, 9, 2072. Available online: https://www.frontiersin.org/article/10.3389/fmicb.2018.02072/full (accessed on 29 March 2020). [CrossRef]
- Watson, D.J.; Meyers, P.R.; Acquah, K.S.; Dziwornu, G.A.; Barnett, C.B.; Wiesner, L. Discovery of Novel Cyclic Ethers with Synergistic Antiplasmodial Activity in Combination with Valinomycin. Molecules 2021, 26, 7494. Available online: https://www.mdpi.com/1420-3049/26/24/7494 (accessed on 9 December 2022). [CrossRef]
- Shirling, E.B.; Gottlieb, D. Methods for characterization of Streptomyces species. Int. J. Syst. Bacteriol. 1966, 16, 313–340. Available online: http://www.ncbi.nlm.nih.gov/pubmed/19172199 (accessed on 2 September 2019). [CrossRef]
- German Culture Collection. DSMZ #553 Recipe. 2007. Available online: https://www.dsmz.de/microorganisms/medium/pdf/DSMZ_Medium553.pdf (accessed on 7 April 2020).
- Atlas, R. Handbook of Microbiological Media, 4th ed.; Parks, L.C., Ed.; CRC Press: Boca Raton, FL, USA, 2010; 1935p, Available online: https://www.taylorfrancis.com/books/9781439804087 (accessed on 15 May 2019).
- Ara, I.; Kudo, T. Sphaerosporangium gen. nov., a new member of the family Streptosporangiaceae, with descriptions of three new species as Sphaerosporangium melleum sp. nov., Sphaerosporangium rubeum sp. nov. and Sphaerosporangium cinnabarinum sp. nov., and transfer of Stre. Actinomycetologica 2007, 21, 11–21. Available online: http://joi.jlc.jst.go.jp/JST.JSTAGE/saj/SAJ210102?from=CrossRef (accessed on 2 September 2019). [CrossRef]
- Bartholomew, J.W.; Mittwer, T. The Gram stain. Bacteriol. Rev. 1952, 16, 1–29. Available online: http://www.ncbi.nlm.nih.gov/pubmed/14925025 (accessed on 13 December 2018). [CrossRef]
- de Melo, C.S.; Feng, T.S.; van der Westhuyzen, R.; Gessner, R.K.; Street, L.J.; Morgans, G.L.; Warner, D.F.; Moosa, A.; Naran, K.; Lawrence, N.; et al. Aminopyrazolo[1,5-a]pyrimidines as potential inhibitors of Mycobacterium tuberculosis: Structure activity relationships and ADME characterization. Bioorg. Med. Chem. 2015, 23, 7240–7250. Available online: https://linkinghub.elsevier.com/retrieve/pii/S0968089615300985 (accessed on 24 November 2022). [CrossRef] [PubMed]
- Franzblau, S.G.; DeGroote, M.A.; Cho, S.H.; Andries, K.; Nuermberger, E.; Orme, I.M.; Mdluli, K.; Angulo-Barturen, I.; Dick, T.; Dartois, V.; et al. Comprehensive analysis of methods used for the evaluation of compounds against Mycobacterium tuberculosis. Tuberculosis 2012, 92, 453–488. Available online: https://linkinghub.elsevier.com/retrieve/pii/S1472979212001400 (accessed on 24 November 2022). [CrossRef]
- Tang, Y.J.; Shui, W.; Myers, S.; Feng, X.; Bertozzi, C.; Keasling, J.D. Central metabolism in Mycobacterium smegmatis during the transition from O2-rich to O2-poor conditions as studied by isotopomer-assisted metabolite analysis. Biotechnol. Lett. 2009, 31, 1233–1240. Available online: http://link.springer.com/10.1007/s10529-009-9991-7 (accessed on 24 November 2022). [CrossRef] [PubMed]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Cook, A.E.; le Roes, M.; Meyers, P.R. Actinomadura napierensis sp. nov., isolated from soil in South Africa. Int. J. Syst. Evol. Microbiol. 2005, 55, 703–706. Available online: https://www.microbiologyresearch.org/content/journal/ijsem/10.1099/ijs.0.63359-0 (accessed on 18 October 2022). [CrossRef] [PubMed]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. Available online: http://www.genome.org/cgi/doi/10.1101/gr.1239303 (accessed on 7 November 2019). [CrossRef] [PubMed]
- Pluskal, T.; Castillo, S.; Villar-Briones, A.; Orešič, M. MZmine 2: Modular framework for processing, visualizing, and analyzing mass spectrometry-based molecular profile data. BMC Bioinform. 2010, 11, 395. Available online: https://bmcbioinformatics.biomedcentral.com/articles/10.1186/1471-2105-11-395 (accessed on 2 April 2020). [CrossRef] [PubMed]
- Myers, O.D.; Sumner, S.J.; Li, S.; Barnes, S.; Du, X. One Step Forward for Reducing False Positive and False Negative Compound Identifications from Mass Spectrometry Metabolomics Data: New Algorithms for Constructing Extracted Ion Chromatograms and Detecting Chromatographic Peaks. Anal. Chem. 2017, 89, 8696–8703. Available online: https://pubs.acs.org/doi/10.1021/acs.analchem.7b00947 (accessed on 25 June 2020). [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain Name | Growth Medium | Antitubercular Activity (MIC90, µg/mL) | Chinese Hamster Ovary IC50 (µg/mL) | ||
|---|---|---|---|---|---|
| 7H9_ADC_GLU_ TX | 7H9_CAS_GLU_ TX | 7H9_ADC_GLU_ TW | |||
| Actinomadura napierensis B60T | ISP-2 | 1.53 ± 0.80 | 1.25 ± 0.15 | 1.20 ± 0.80 | >100 |
| Streptomyces africanus CPJVR-HT | ISP-2 | 8.05 ± 4.6 | >62.5 ± ND | 0.74 ± 0.35 | >100 |
| Streptomyces strain CW5 | JCM #61 | 0.34 ± 0.22 | 0.21 ± 0.036 | 1.01 ± 0.90 | 0.032 |
| Streptomyces strain HMC5 | JCM #61 | 19.2 ± 6.4 | 2.75 ± 0.040 | 14.2 ± 1.3 | >100 |
| Streptomyces fractus MV32T | DSMZ #553 | 6.30 ± 5.5 | <0.12 ± ND | <0.12 ± ND | 5.6 |
| Streptomyces strain Y10 | DSMZ #553 | <0.12 ± ND | <0.12 ± ND | <0.12 ± ND | 2.4 |
| Streptomyces strain Y10 | JCM #61 | 0.59 ± 0.14 | <0.12 ± ND | <0.12 ± ND | 0.13 |
| Strain Name | Growth Medium | Source | Compound | Theoretical Monoisotopic Mass (Da) | Experimental Mass (Da) * | Accurate Mass Difference (Da) | Mass Error (ppm) |
|---|---|---|---|---|---|---|---|
| Actinomadura napierensis B60T | ISP-2 | Agar | Tetromadurin | 760.4398 | 760.4394 | 0.0004 | 0.526 |
| Liquid | 760.4398 | 760.4403 | −0.0005 | −0.658 |
| Culture Medium | M. tuberculosis H37RvT MIC90 (nM) | Chinese Hamster Ovary IC50 (µM) | Selectivity Index |
|---|---|---|---|
| 7H9_ADC_GLU_TW | 148.1 ± 49 | 13 | |
| 7H9_ADC_GLU_TX | 151.6 ± 70 | 12 | |
| 7H9_CAS_GLU_TW | 73.7 ± 18 | 1.94 ± 8.0 | 26 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Watson, D.J.; Wiesner, L.; Matimela, T.; Beukes, D.; Meyers, P.R. Tandem LC-MS Identification of Antitubercular Compounds in Zones of Growth Inhibition Produced by South African Filamentous Actinobacteria. Molecules 2023, 28, 4276. https://doi.org/10.3390/molecules28114276
Watson DJ, Wiesner L, Matimela T, Beukes D, Meyers PR. Tandem LC-MS Identification of Antitubercular Compounds in Zones of Growth Inhibition Produced by South African Filamentous Actinobacteria. Molecules. 2023; 28(11):4276. https://doi.org/10.3390/molecules28114276
Chicago/Turabian StyleWatson, Daniel J., Lubbe Wiesner, Tlhalefo Matimela, Denzil Beukes, and Paul R. Meyers. 2023. "Tandem LC-MS Identification of Antitubercular Compounds in Zones of Growth Inhibition Produced by South African Filamentous Actinobacteria" Molecules 28, no. 11: 4276. https://doi.org/10.3390/molecules28114276
APA StyleWatson, D. J., Wiesner, L., Matimela, T., Beukes, D., & Meyers, P. R. (2023). Tandem LC-MS Identification of Antitubercular Compounds in Zones of Growth Inhibition Produced by South African Filamentous Actinobacteria. Molecules, 28(11), 4276. https://doi.org/10.3390/molecules28114276