
The Effect of Progestins on Cytokine Production in the Peripheral Blood Mononuclear Cells of Menopausal Women and Their Luminol-Dependent Chemiluminescence

Abstract
:1. Introduction
2. Results
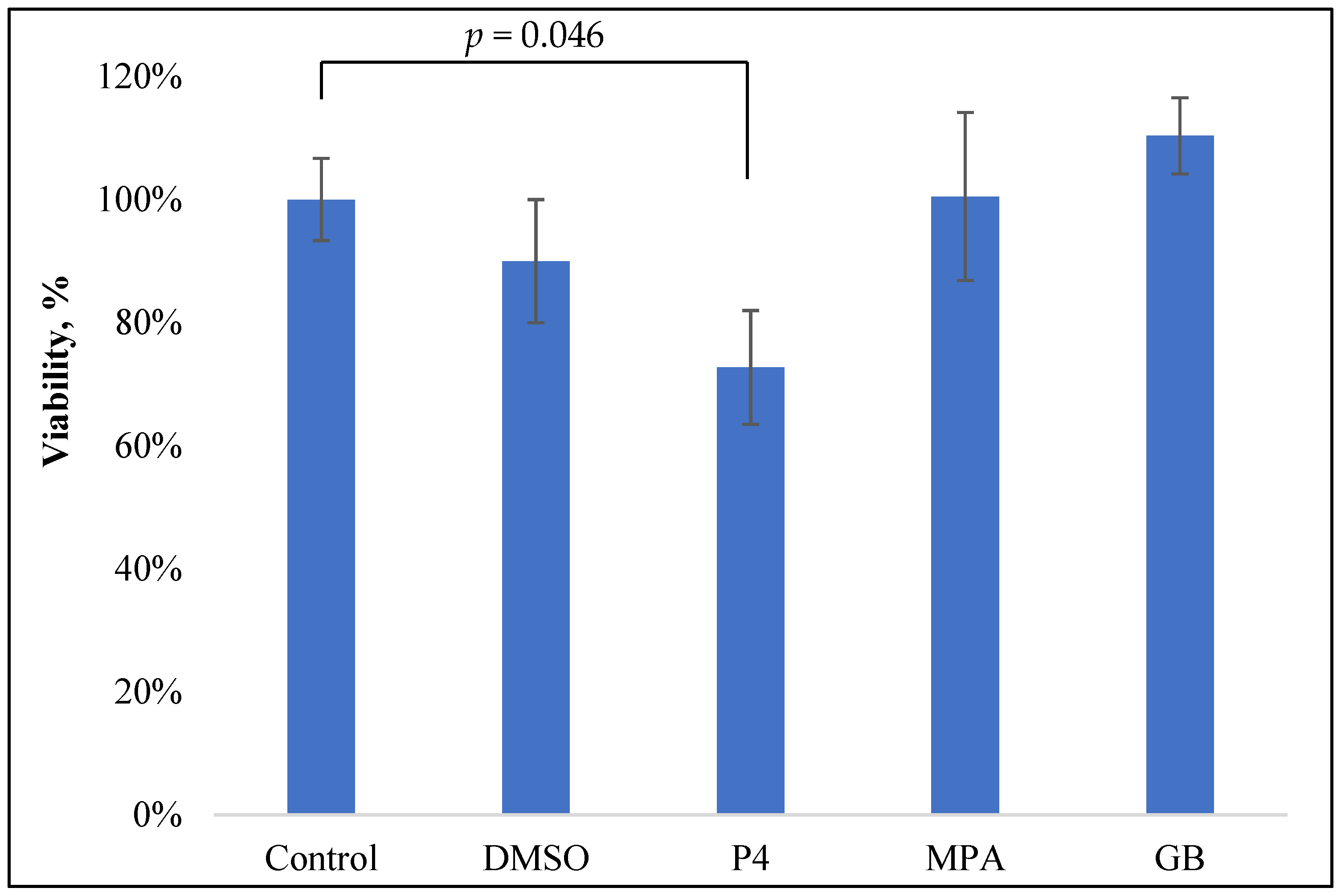
2.1. Progestin Influence on the Viability of PBMCs
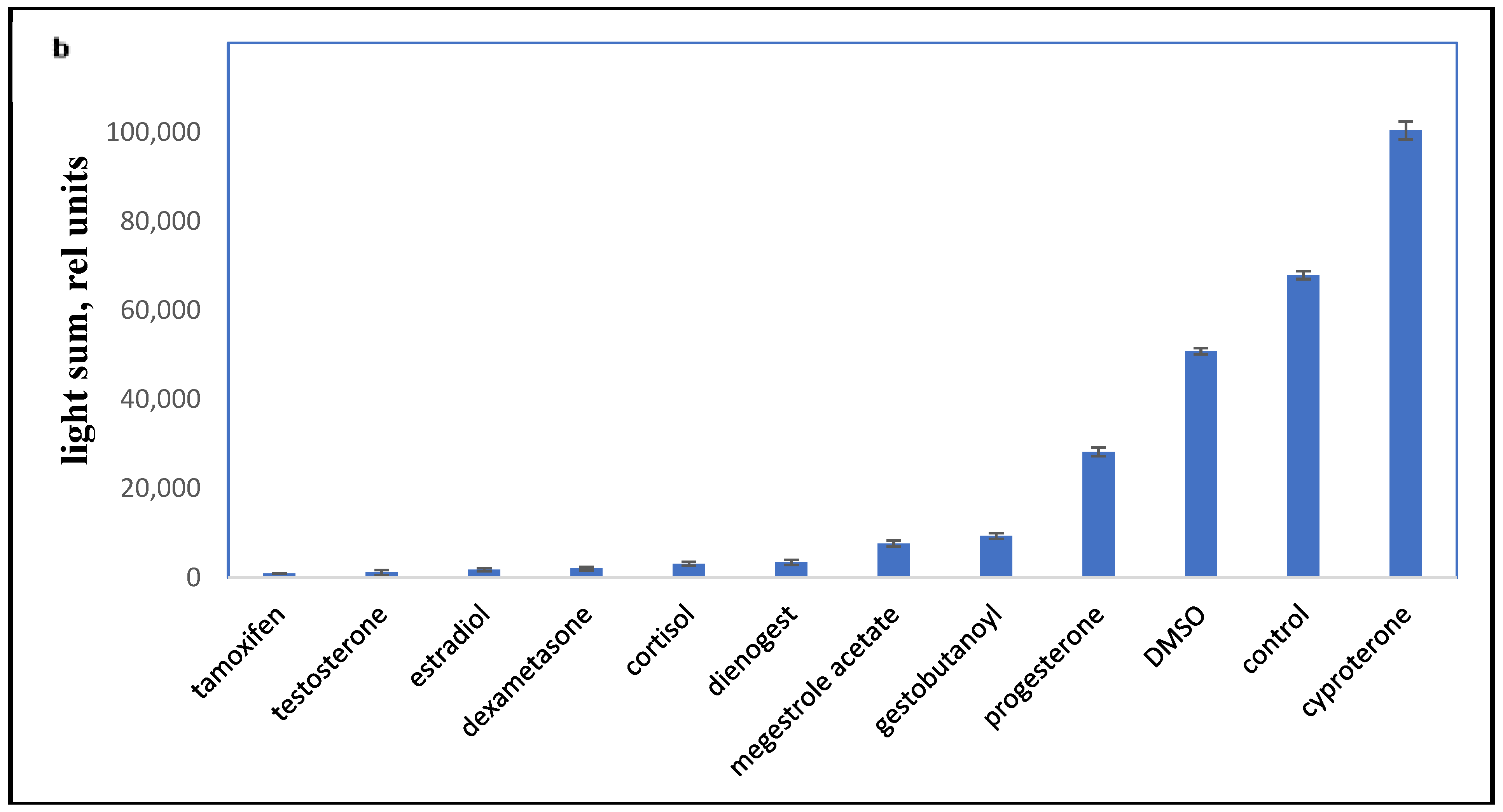
2.2. Influence of Progestins on the PBMC Oxidant Capacity
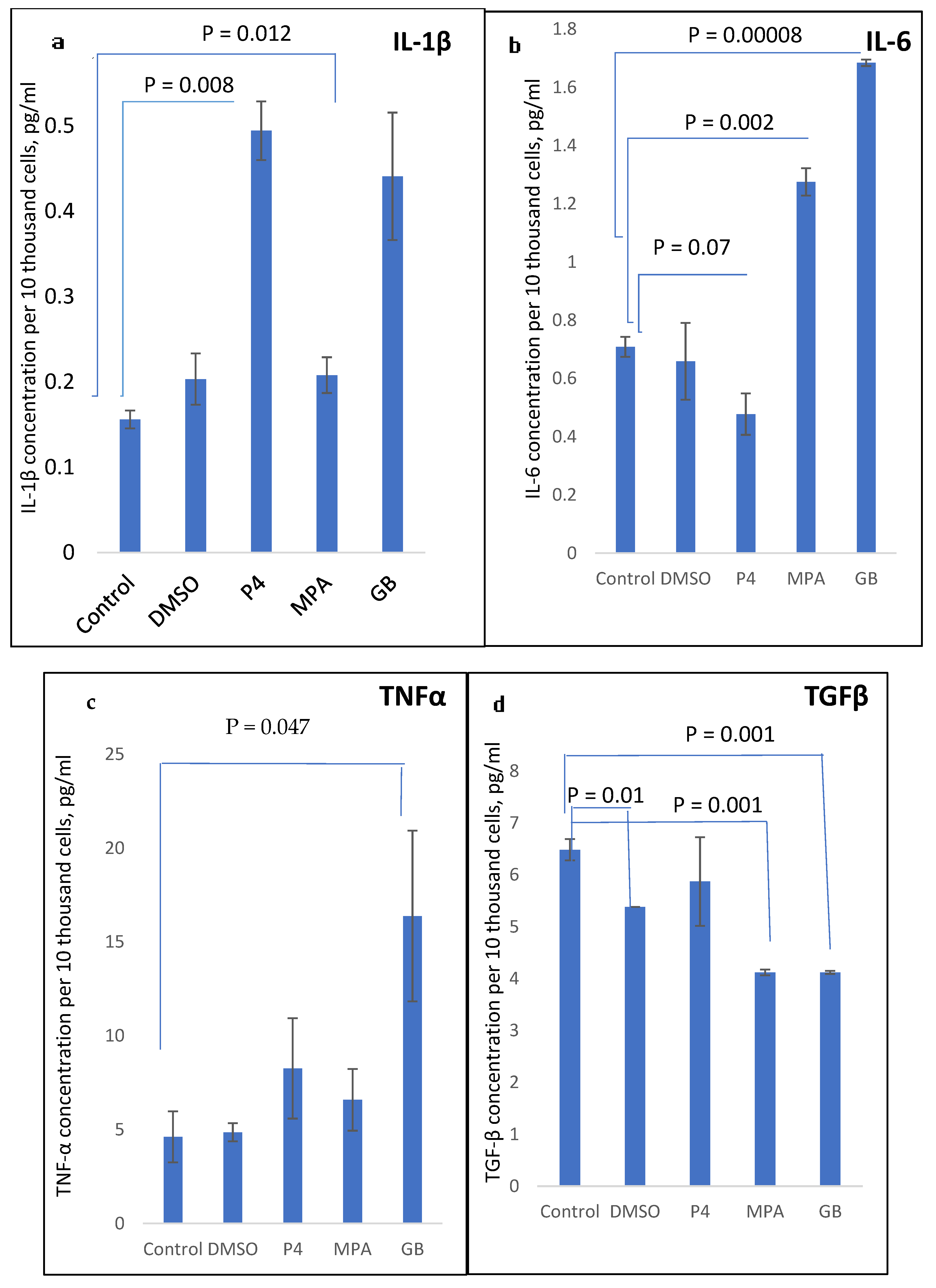
2.3. Influence of Progestins on the Production of Cytokines in PBMCs
3. Discussion
4. Materials and Methods
4.1. Drug Preparation
4.2. Participants
4.3. MTT Assay
4.4. LCC Measurement by the Luuminol-Dependent Chemiluminescence Assay
4.5. ELISA
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
Abbreviations
| GPX | Glutathione peroxidase |
| GST | Glutathione S Transferase |
| HRT | Hormone replacement therapy |
| LDC | Luminol-dependent chemiluminescence |
| LPS | lipopolysaccharide |
| MTT | 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenytetrazolium bromide |
| NOS | Nitric oxide synthases |
| PBMC | Peripheral blood mononuclear cells |
| PHA | Phytohaemagglutinin |
| SOD | Superoxide dismutase |
References
- Piccinni, M.P.; Lombardelli, L.; Logiodice, F.; Kullolli, O.; Maggi, E.; Barkley, M.S. Medroxyprogesterone Acetate Decreases Th1, Th17, and Increases Th22 Responses via AHR Signaling Which Could Affect Susceptibility to Infections and Inflammatory Disease. Front. Immunol. 2019, 10, 642. [Google Scholar] [CrossRef] [PubMed]
- Available online: https://www.cancer.org/cancer/endometrial-cancer/treating/hormone-therapy.html (accessed on 7 March 2023).
- Fedotcheva, T.A. Clinical Use of Progestins and Their Mechanisms of Action: Present and Future (Review). Sovrem. Tekhnologii Med. 2021, 13, 93–106. [Google Scholar] [CrossRef] [PubMed]
- Petrosyan, M.A.; Balashova, N.N.; Polyanskikh, L.S.; Baziyan, E.V.; Tral’, T.G.; Fasakhutdinova, L.K.; Razygraev, A.V.; Sapronov, N.S. Influence of progesterone analogs on endometrioid heterotopia in experimental model of endometriosis. Exp. Clin. Pharmacol. 2018, 81, 14–19. [Google Scholar]
- Sergeev, P.V.; Semeĭkin, A.V.; Smirnova, Z.S.; Rzheznikov, V.M.; Grinenko, G.S.; Fedosov, A.V.; Fedotcheva, T.A.; Shimanovskiĭ, N.L. Antitumor activity of the new gestagen 17alpha-acetoxy-3beta-butanoyloxy-6-methyl-pregna-4,6-dien-20-one. Exp. Clin. Pharmacol. 2004, 67, 54–56. [Google Scholar]
- Polikarpova, A.V.; Levina, I.S.; Sigai, N.V.; Zavarzin, I.V.; Morozov, I.A.; Rubtsov, P.M.; Guseva, A.A.; Smirnova, O.V.; Shchelkunova, T.A. Immunomodulatory effects of progesterone and selective ligands of membrane progesterone receptors. Steroids 2019, 145, 5–18. [Google Scholar] [CrossRef] [PubMed]
- Marks, M.A.; Gravitt, P.E.; Burk, R.D.; Studentsov, Y.; Farzadegan, H.; Klein, S.L. Progesterone and 17beta-estradiol enhance regulatory responses to human papillomavirus type 16 virus-like particles in peripheral blood mononuclear cells from healthy women. Clin. Vaccine Immunol. 2010, 17, 609–617. [Google Scholar] [CrossRef]
- AbdulHussain, G.; Azizieh, F.; Makhseed, M.; Raghupathy, R. Effects of Progesterone, Dydrogesterone and Estrogen on the Production of Th1/Th2/Th17 Cytokines by Lymphocytes from Women with Recurrent Spontaneous Miscarriage. J. Reprod. Immunol. 2020, 140, 103132. [Google Scholar] [CrossRef]
- Pfeilschifter, J.; Köditz, R.; Pfohl, M.; Schatz, H. Changes in proinflammatory cytokine activity after menopause. Endocr. Rev. 2002, 23, 90–119. [Google Scholar] [CrossRef]
- Yuan, Y.; Shimizu, I.; Shen, M.; Aoyagi, E.; Takenaka, H.; Itagaki, T.; Urata, M.; Sannomiya, K.; Kohno, N.; Tamaki, K.; et al. Effects of estradiol and progesterone on the proinflammatory cytokine production by mononuclear cells from patients with chronic hepatitis C. World J. Gastroenterol. 2008, 14, 2200–2207. [Google Scholar] [CrossRef]
- Straub, R.H. The complex role of estrogens in inflammation. Endocr. Rev. 2007, 28, 521–574. [Google Scholar] [CrossRef]
- Fedotcheva, T.A.; Sheichenko, O.P.; Fedotcheva, N.I. New Properties and Mitochondrial Targets of Polyphenol Agrimoniin as a Natural Anticancer and Preventive. Agent Pharm. 2021, 13, 2089. [Google Scholar] [CrossRef] [PubMed]
- Maeda, Y.; Ohtsuka, H.; Tomioka, M.; Tanabe, T.; Nambo, Y.; Uematsu, H.; Oikawa, M. Effect of progesterone on the in vitro response of peripheral blood mononuclear cells stimulated by Escherichia coli in mares. J. Vet. Med. Sci. 2012, 74, 629–632. [Google Scholar] [CrossRef] [PubMed]
- Huber, N.; Marasco, V.; Painer, J.; Vetter, S.G.; Göritz, F.; Kaczensky, P.; Walzer, C. Leukocyte Coping Capacity: An Integrative Parameter for Wildlife Welfare Within Conservation Interventions. Front. Vet. Sci. 2019, 6, 105. [Google Scholar] [CrossRef] [PubMed]
- Chausova, S.V.; Gurevich, K.G.; Bondareva, G.P.; Filatov, O.J.; Malyshev, I.Y. The role of cellular mediators in the development of the phenomenon of inhibition induced by barium sulfate luminol-dependent chemiluminescence of blood under the influence of non-steroidal anti-inflammatory drugs in patients with intolerance to these drugs. Patol. Fiziol. I Eksperimental’naia Ter. 2015, 3, 89–93. [Google Scholar]
- Marren, K. Dimethyl sulfoxide: An effective penetration enhancer for topical administration of NSAIDs. Phys. Sportsmed. 2011, 39, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Drugbank. 2005. Available online: https://go.drugbank.com/drugs/DB01093 (accessed on 30 January 2023).
- Ivkin, D.Y.; Okovitiy, S.V.; Ivkina, A.S.; Anisimova, N.A. Dimetilsulfoxid-veschestvo s pleyotropnyimi effektami, aktual’nyimi pri zabolevaniyah oporno-dvigatel’nogo apparata. Lechaschiy Vrach 2019, 4, 19–23. [Google Scholar]
- Hughes, D.L.; Richards, R.S.; Lexis, L.A. Using chemiluminescence to determine whole blood antioxidant capacity in rheumatoid arthritis and Parkinson’s disease patients. Luminescence 2018, 33, 764–770. [Google Scholar] [CrossRef]
- Costa, D.; Moutinho, L.; Lima, J.L.F.C.; Fernandes, E. Antioxidant activity and inhibition of human neutrophil oxidative burst mediated by arylpropionic acid non-steroidal anti-inflammatory drugs. Biol. Pharm. Bull. 2006, 29, 1659–1670. [Google Scholar] [CrossRef]
- Raghupathy, R.; Al-Azemi, M. Modulation of Cytokine Production by the Dydrogesterone Metabolite Dihydrodydrogesterone. Am. J. Reprod. Immunol. 2015, 74, 419–426. [Google Scholar] [CrossRef]
- Raghupathy, R.; Mutawa, E.A.; Makhseed, M.; Al-Azemi, M.; Azizieh, F. Redirection of cytokine production by lymphocytes from women with pre-term delivery by dydrogesterone. Am. J. Reprod. Immunol. 2007, 58, 31–38. [Google Scholar] [CrossRef]
- Hantoushzadeh, S.; Aliabad, R.A.; Norooznezhad, A.H. Antibiotics, Inflammation, and Preterm Labor: A Missed Conclusion. J. Inflamm. Res. 2020, 13, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Mantovani, G.; Macciò, A.; Lai, P.; Massa, E.; Ghiani, M.; Santona, M.C. Cytokine involvement in cancer anorexia/cachexia: Role of megestrol acetate and medroxyprogesterone acetate on cytokine downregulation and improvement of clinical symptoms. Crit. Rev. Oncog. 1998, 9, 99–106. [Google Scholar] [CrossRef] [PubMed]
- Kurebayashi, J.; Yamamoto, S.; Otsuki, T.; Sonoo, H. Medroxyprogesterone acetate inhibits interleukin 6 secretion from KPL-4 human breast cancer cells both in vitro and in vivo: A possible mechanism of the anticachectic effect. Br. J. Cancer 1999, 79, 631–636. [Google Scholar] [CrossRef]
- Groh, L.A.; Verel, D.E.; van der Heijden, C.D.C.C.; Matzaraki, V.; Moorlag, S.J.C.F.M.; de Bree, L.C.; Koeken, V.A.C.M.; Mourits, V.P.; Keating, S.T.; van Puffelen, J.H.; et al. Immune modulatory effects of progesterone on oxLDL-induced trained immunity in monocytes. J. Leukoc. Biol. 2022, 112, 279–288. [Google Scholar] [CrossRef]
- Brundin, P.M.A.; Landgren, B.-M.; Fjällström, P.; Shamekh, M.M.; Gustafsson, J.-A.; Johansson, A.F.; Nalvarte, I. Expression of Sex Hormone Receptor and Immune Response Genes in Peripheral Blood Mononuclear Cells during the Menstrual Cycle. Front. Endocrinol. 2021, 12, 721813. [Google Scholar] [CrossRef] [PubMed]
- Fedotcheva, T.A.; Fedotcheva, N.I.; Shimanovsky, N.L. Progesterone as an Anti-Inflammatory Drug and Immunomodulator: New Aspects in Hormonal Regulation of the Inflammation. Biomolecules 2022, 12, 1299. [Google Scholar] [CrossRef]
- Koubovec, D.; Ronacher, K.; Stubsrud, E.; Louw, A.; Hapgood, J.P. Synthetic progestins used in HRT have different glucocorticoid agonist properties. Mol. Cell Endocrinol. 2005, 242, 23–32. [Google Scholar] [CrossRef]
- Ozdemir, F.; Ovali, E.; Aydin, F.; Kavgaci, H.; Büyükcelik, A.; Ucar, F.; Sönmez, M. The in-vitro effects of medroxyprogesterone acetate on acidic pH induced apoptosis of periferal blood mononuclear cells. J. Exp. Clin. Cancer Res. 2004, 23, 93–95. [Google Scholar]
- Hardy, D.B.; Janowski, B.A.; Corey, D.R.; Mendelson, C.R. Progesterone receptor plays a major antiinflammatory role in human myometrial cells by antagonism of nuclear factor-kappaB activation of cyclooxygenase 2 expression. Mol. Endocrinol. 2006, 20, 2724–2733. [Google Scholar] [CrossRef]
- Miller, L.; Hunt, J.S. Regulation of TNF-alpha production in activated mouse macrophages by progesterone. J. Immunol. 1998, 160, 5098–5104. [Google Scholar] [CrossRef]
- Africander, D.; Verhoog, N.; Hapgood, J.P. Molecular mechanisms of steroid receptor-mediated actions by synthetic progestins used in HRT and contraception. Steroids 2011, 76, 636–652. [Google Scholar] [CrossRef] [PubMed]
- Zeynalov, O.A.; Savinova, T.S.; Andryushina, V.A.; Petrosyan, M.A. Synthetic analogues of progesterone in in vitro and in vivo models. Biomedicine 2018, 3, 80–89. [Google Scholar]
- Liga, A.B.; Ukhina, T.V.; Shimanovskii, N.L. Activity lysosomal enzymes in rat skin fibroblasts after treatment with progesterone and new gestagen ABMP. Bull. Exp. Biol. Med. 2008, 145, 44–46. [Google Scholar] [CrossRef] [PubMed]
- Sergeev, P.V.; Fedotcheva, T.A.; Rzheznikov, V.M.; Grinenko, G.S.; Semeĭkin, A.V.; Vetchinkina, V.B.; Atroshkin, K.A.; Shimanovskiĭ, N.L. A new Russian gestagen with anticancer activity. Vestn. Ross. Akad. Med. Nauk 2007, 5, 27–32. [Google Scholar]
- Kareva, E.N.; Grinenko, G.S.; Gasparian, N.D.; Ovchinnikova, E.V.; Gorenkova, O.S. Influence of the structure of synthetic gestagens on their binding to progesteron receptors in the endometrium. Eksp. Klin. Farm. 2006, 69, 36–38. [Google Scholar]
- Sitruk-Ware, R. Pharmacological profile of progestins. Maturitas 2008, 61, 151–157. [Google Scholar] [CrossRef] [PubMed]
- Stanczyk, F.Z.; Hapgood, J.P.; Winer, S.; Mishell, D.R. Progestogens used in postmenopausal hormone therapy: Differences in their pharmacological properties, intracellular actions, and clinical effects. Endocr. Rev. 2013, 34, 171–208. [Google Scholar] [CrossRef]
- Schindler, A.E. Progestational effects of dydrogesterone in vitro, in vivo and on the human endometrium. Maturitas 2009, 65, S3–S11. [Google Scholar] [CrossRef]
- Zeinalov, O.A.; Andryushina, V.A.; Yaderets, V.V. New synthetic analogs of progesterone: From the search for an active molecule to clinical use (review of our own research). Pharm. Chem. J. 2022, 56, 145–154. [Google Scholar] [CrossRef]
- Africander, D.; Louw, R.; Verhoog, N.; Noeth, D.; Hapgood, J.P. Differential regulation of endogenous pro-inflammatory cytokine genes by medroxyprogesterone acetate and norethisterone acetate in cell lines of the female genital tract. Contraception 2011, 84, 423–435. [Google Scholar] [CrossRef]
- Proskurnina, E.V.; Polimova, A.M.; Sozarukova, M.M.; Vladimirov, Y.A.; Prudnikova, M.A.; Ametov, A.S. Kinetic chemiluminescence as a method for oxidative stress evaluation in examinations of patients with type 2 diabetes mellitus. Bull. Exp. Biol. Med. 2016, 161, 131–133. [Google Scholar] [CrossRef] [PubMed]
- Gaudio, E.; Bordin, S.; Lora, I.; Lora, M.; Massignani, M.; Benedictis, G.M.D. Leukocyte coping capacity chemiluminescence as an innovative tool for stress and pain assessment in calves undergoing ring castration. J. Anim. Sci. 2018, 96, 4579–4589. [Google Scholar] [PubMed]
- Nascimento, C.R.; Fernandes, N.A.R.; Maldonado, L.A.G.; Junior, C.R. Comparison of monocytic cell lines U937 and THP-1 as macrophage models for in vitro studies. Biochem. Biophys. Rep. 2022, 32, 101383. [Google Scholar] [CrossRef] [PubMed]
- Hall, O.J.; Klein, S.L. Progesterone-based compounds affect immune responses and susceptibility to infections at diverse mucosal sites. Mucosal. Immunol. 2017, 10, 1097–1107. [Google Scholar] [CrossRef] [PubMed]
- Mantovani, G.; Macciò, A.; Esu, S.; Lai, P.; Santona, M.C.; Massa, E.; Dessì, D.; Melis, G.B.; Del Giacco, G.S. Medroxyprogesterone acetate reduces the in vitro production of cytokines and serotonin involved in anorexia/cachexia and emesis by peripheral blood mononuclear cells of cancer patients. Eur. J. Cancer 1997, 33, 602–607. [Google Scholar] [CrossRef]
- Huijbregts, R.P.; Helton, E.S.; Michel, K.G.; Sabbaj, S.; Richter, H.E.; Goepfert, P.A.; Hel, Z. Hormonal contraception and HIV-1 infection: Medroxyprogesterone acetate suppresses innate and adaptive immune mechanisms. Endocrinology 2013, 154, 1282–1295. [Google Scholar] [CrossRef]
- García-Juárez, M.; Camacho-Morales, A. Defining the Role of Anti- and Pro-inflammatory Outcomes of Interleukin-6 in Mental Health. Neuroscience 2022, 492, 32–46. [Google Scholar] [CrossRef]
- Askari, V.R.; Rahimi, V.B.; Zargarani, R.; Ghodsi, R.; Boskabady, M.; Boskabady, M.H. Anti-oxidant and anti-inflammatory effects of auraptene on phytohemagglutinin (PHA)-induced inflammation in human lymphocytes. Pharmacol. Rep. 2021, 73, 154–162. [Google Scholar] [CrossRef]
- Komane, M.; Avenant, C.; Louw-du Toit, R.; Africander, D.J.; Hapgood, J.P. Differential off-target glucocorticoid activity of progestins used in endocrine therapy. Steroids 2022, 182, 108998. [Google Scholar] [CrossRef]
- Preciado-Martínez, E.; García-Ruíz, G.; Flores-Espinosa, P.; Bermejo-Martínez, L.; Espejel-Nuñez, A.; Estrada-Gutiérrez, G.; Razo-Aguilera, G.; Granados-Cepeda, M.; Helguera-Repetto, A.C.; Irles, C.; et al. Progesterone suppresses the lipopolysaccharide-induced pro-inflammatory response in primary mononuclear cells isolated from human placental blood. Immunol. Investig. 2018, 47, 181–195. [Google Scholar] [CrossRef]
- Garcia-Ruíz, G.; Flores-Espinosa, P.; Preciado-Martínez, E.; Bermejo-Martínez, L.; Espejel-Nuñez, A.; Estrada-Gutierrez, G.; Maida-Claros, R.; Flores-Pliego, A.; Zaga-Clavellina, V. In vitro progesterone modulation on bacterial endotoxin-induced production of IL-1β, TNF-α, IL-6, IL-8, IL-10, MIP-1α, and MMP-9 in pre-labor human term placenta. Reprod. Biol. Endocrinol. 2015, 13, 115. [Google Scholar] [CrossRef] [PubMed]
- Bouman, A.; Schipper, M.; Heineman, M.J.; Faas, M. 17beta-estradiol and progesterone do not influence the production of cytokines from lipopolysaccharide-stimulated monocytes in humans. Fertil. Steril. 2004, 82 (Suppl. S3), 1212–1219. [Google Scholar] [CrossRef] [PubMed]
- Jain, S.K.; Kannan, K.; Prouty, L.; Jain, S.K. Progesterone, but not 17beta-estradiol, increases TNF-alpha secretion in U937 monocytes. Cytokine 2004, 26, 102–105. [Google Scholar] [CrossRef] [PubMed]
- Campe, K.-N.J.; Redlich, A.; Zenclussen, A.C.; Busse, M. An increased proportion of progesterone receptor A in peripheral B cells from women who ultimately underwent spontaneous preterm birth. J. Reprod. Immunol. 2022, 154, 103756. [Google Scholar] [CrossRef]
- Hudić, I.; Szekeres-Bartho, J.; Fatušić, Z.; Stray-Pedersen, B.; Dizdarević-Hudić, L.; Latifagić, A.; Hotić, N.; Kamerić, L.; Mandžić, A. Dydrogesterone supplementation in women with threatened preterm delivery--the impact on cytokine profile, hormone profile, and progesterone-induced blocking factor. J. Reprod. Immunol. 2011, 92, 103–107. [Google Scholar] [CrossRef]
- Konecna, L.; Yan, M.S.; Miller, L.E.; Schölmerich, J.; Falk, W.; Straub, R.H. Modulation of IL-6 production during the menstrual cycle in vivo and in vitro. Brain Behav. Immun. 2000, 14, 49–61. [Google Scholar] [CrossRef]
- Sciarra, F.; Campolo, F.; Franceschini, E.; Carlomagno, F.; Venneri, M.A. Gender-Specific Impact of Sex Hormones on the Immune System. Int. J. Mol. Sci. 2023, 24, 6302. [Google Scholar] [CrossRef]
- Stefanaki, C.; Bacopoulou, F.; Chrousos, G.P. Gut Microsex/Genderome, Immunity and the Stress Response in the Sexes: An Updated Review. Sexes 2022, 3, 533–545. [Google Scholar] [CrossRef]
- Lissauer, D.; Eldershaw, S.A.; Inman, C.F.; Coomarasamy, A.; Moss, P.A.; Kilby, M.D. Progesterone promotes maternal-fetal tolerance by reducing human maternal T-cell polyfunctionality and inducing a specific cytokine profile. Eur. J. Immunol. 2015, 45, 2858–2872. [Google Scholar] [CrossRef]
- Omu, A.E.; Al-Azemi, M.K.; Makhseed, M.; Al-Oattan, F.; Ismail, A.A.; Al-Tahir, S.; Al-Busiri, N. Differential expression of T-helper cytokines in the peritoneal fluid of women with normal ovarian cycle compared with women with chronic anovulation. Acta Obstet. Gynecol. Scand. 2003, 82, 603–609. [Google Scholar] [CrossRef]
- Baysal, B. Comparison of the resorbable barrier interceed (TC7) and preoperative use of medroxyprogesterone acetate in postoperative adhesion prevention. Clin. Exp. Obstet. Gynecol. 2001, 28, 126–127. [Google Scholar] [PubMed]
- Böyum, A. Isolation of mononuclear cells and granulocytes from human blood. Isolation of mononuclear cells by one centrifugation, and of granulocytes by combining centrifugation and sedimentation at 1 g. Scand. J. Clin. Lab. Investig. Suppl. 1968, 97, 77–89. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Progestin | RBA, % | Reference and Comment |
|---|---|---|
| P4 | 100 * | [38,39] |
| MPA | 298 * | [38,39] |
| Levonorgestrel | 323 * | [38,39] |
| Dienogest | 10 * | [38,39] |
| Drospirenon | 19 * | [38,39] |
| Didrogesterone | 85 * | [40] |
| Gestobutanoyl | 4.5 ** | [34], rabbit uterus tissue |
| 0.91 ** | [34], rat uterus tissue | |
| 59 ** | [37], human endometrium tissue | |
| 1700 ** | [36], HeLa cancer cell line |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pavlik, T.I.; Shimanovsky, N.L.; Zemlyanaya, O.A.; Fedotcheva, T.A. The Effect of Progestins on Cytokine Production in the Peripheral Blood Mononuclear Cells of Menopausal Women and Their Luminol-Dependent Chemiluminescence. Molecules 2023, 28, 4354. https://doi.org/10.3390/molecules28114354
Pavlik TI, Shimanovsky NL, Zemlyanaya OA, Fedotcheva TA. The Effect of Progestins on Cytokine Production in the Peripheral Blood Mononuclear Cells of Menopausal Women and Their Luminol-Dependent Chemiluminescence. Molecules. 2023; 28(11):4354. https://doi.org/10.3390/molecules28114354
Chicago/Turabian StylePavlik, Tatiana I., Nikolay L. Shimanovsky, Olga A. Zemlyanaya, and Tatiana A. Fedotcheva. 2023. "The Effect of Progestins on Cytokine Production in the Peripheral Blood Mononuclear Cells of Menopausal Women and Their Luminol-Dependent Chemiluminescence" Molecules 28, no. 11: 4354. https://doi.org/10.3390/molecules28114354
APA StylePavlik, T. I., Shimanovsky, N. L., Zemlyanaya, O. A., & Fedotcheva, T. A. (2023). The Effect of Progestins on Cytokine Production in the Peripheral Blood Mononuclear Cells of Menopausal Women and Their Luminol-Dependent Chemiluminescence. Molecules, 28(11), 4354. https://doi.org/10.3390/molecules28114354