


Evaluation of Antioxidant Activities from a Sustainable Source of Okara Protein Hydrolysate Using Enzymatic Reaction

 , , ,
, , ,
Abstract
:1. Introduction
2. Results and Discussion
2.1. The Selection of Optimum Protease Based on In Silico Analysis
2.2. Peptide Ranking
2.3. Physiochemical Properties of Peptides
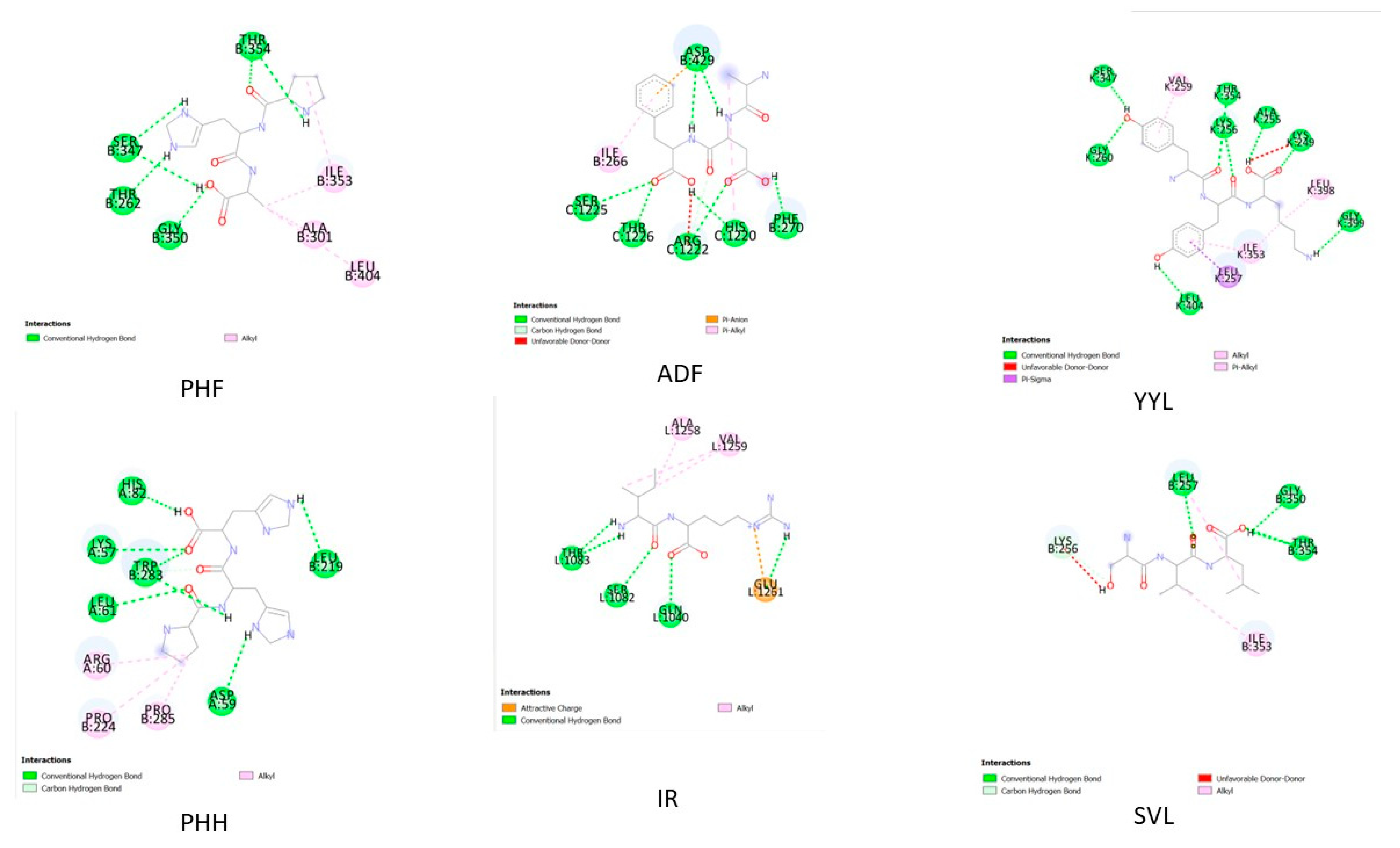
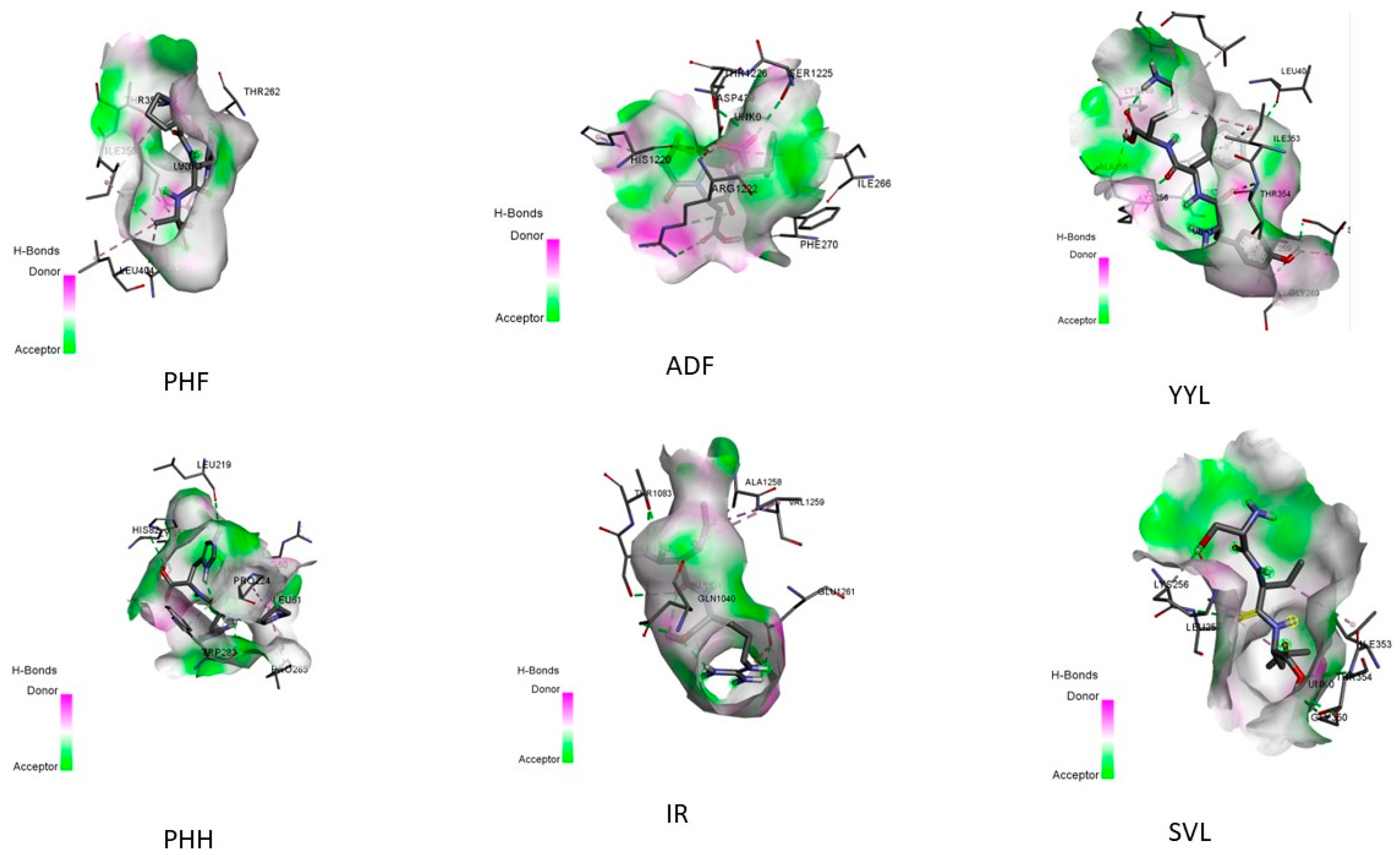
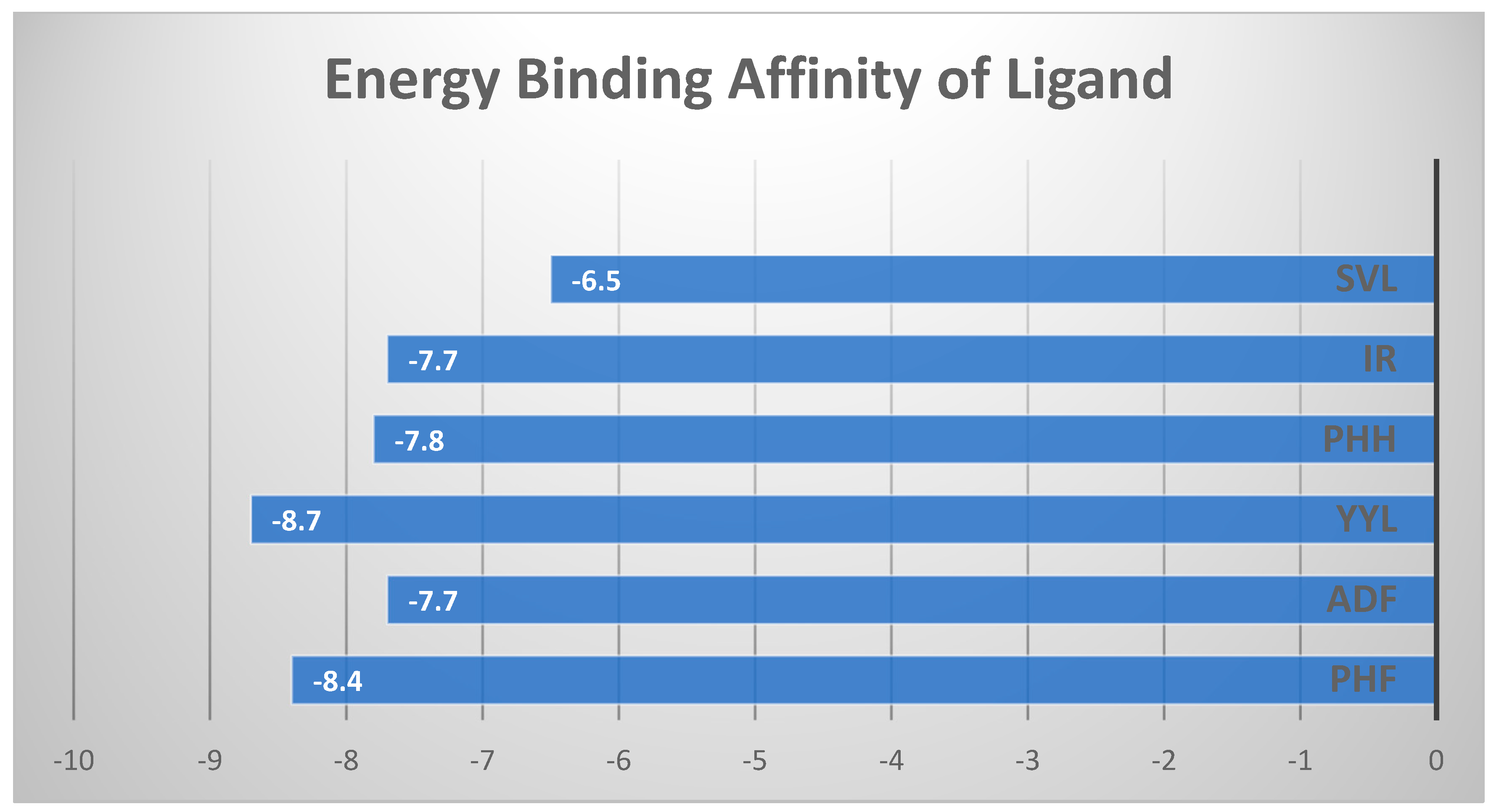
2.4. In Silico Peptide Docking with an Antioxidant Enzyme (Binding Affinity and Interaction)
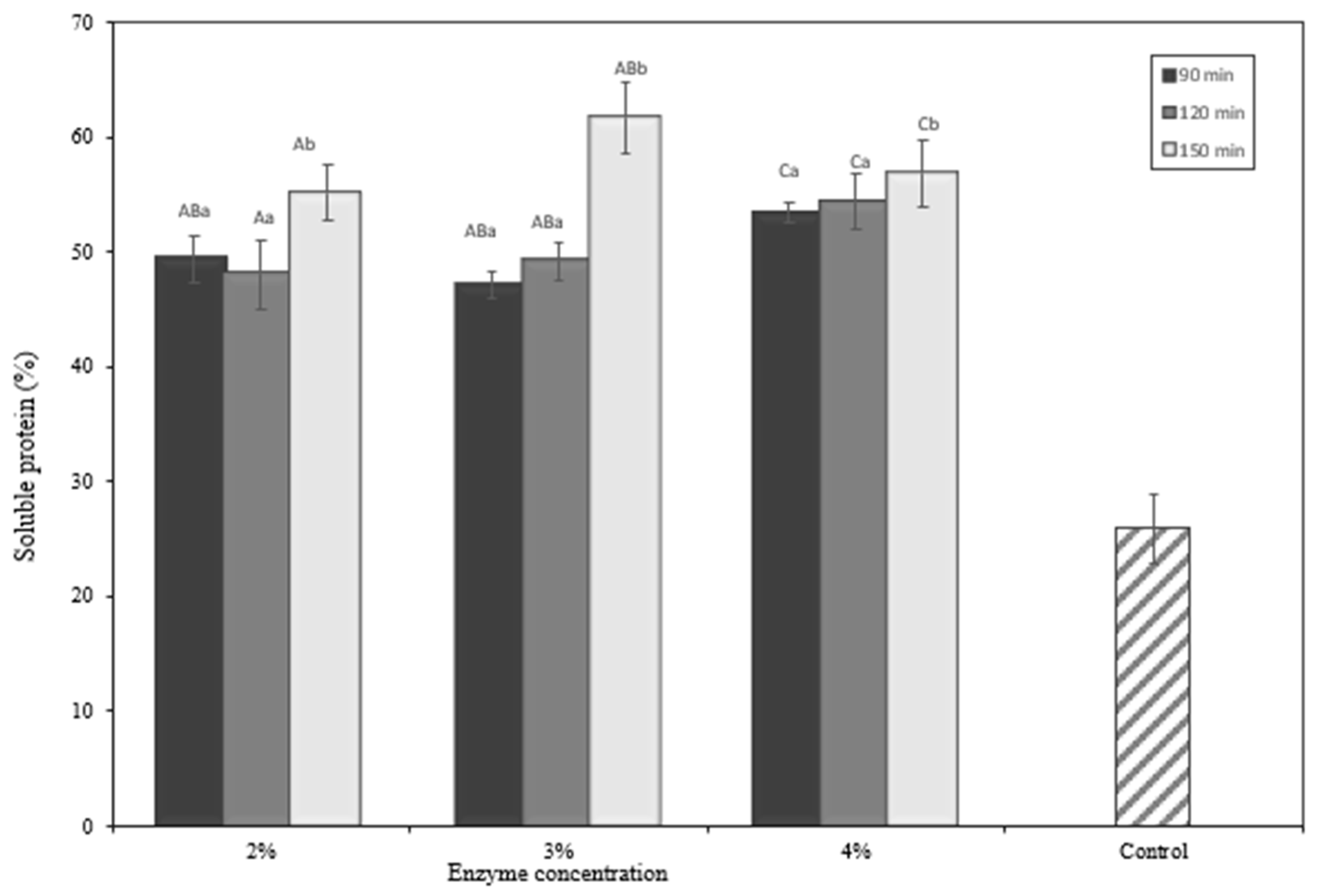
2.5. Proximate Analysis
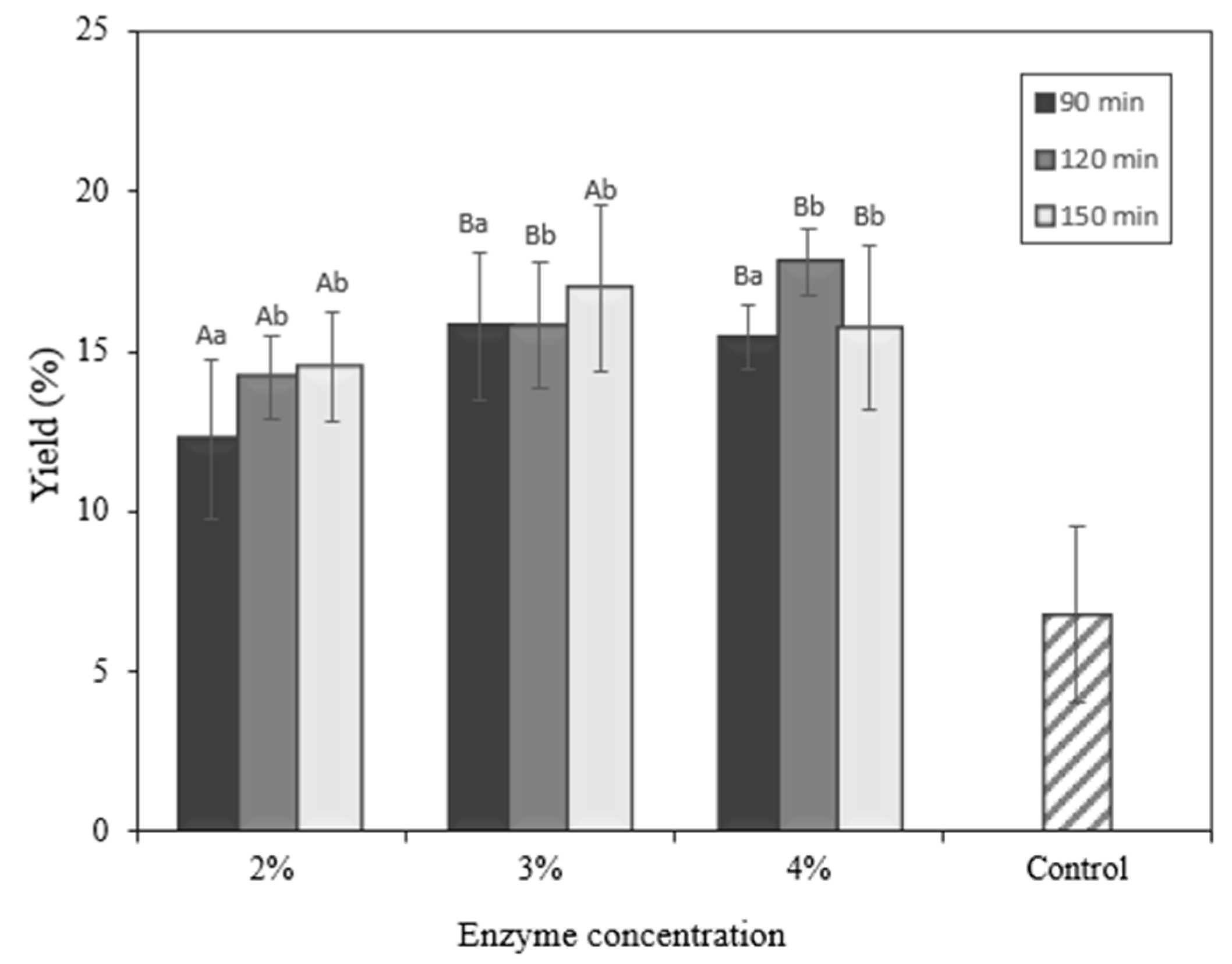
2.6. Yield
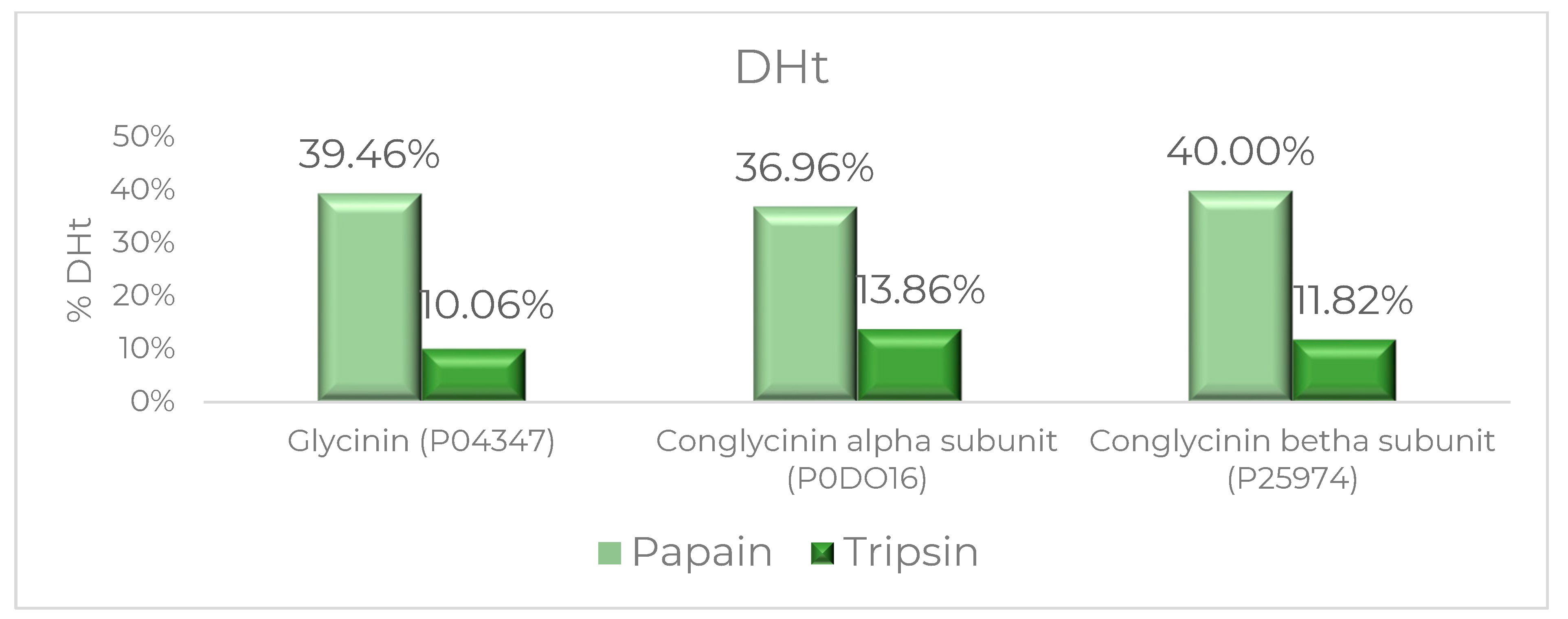
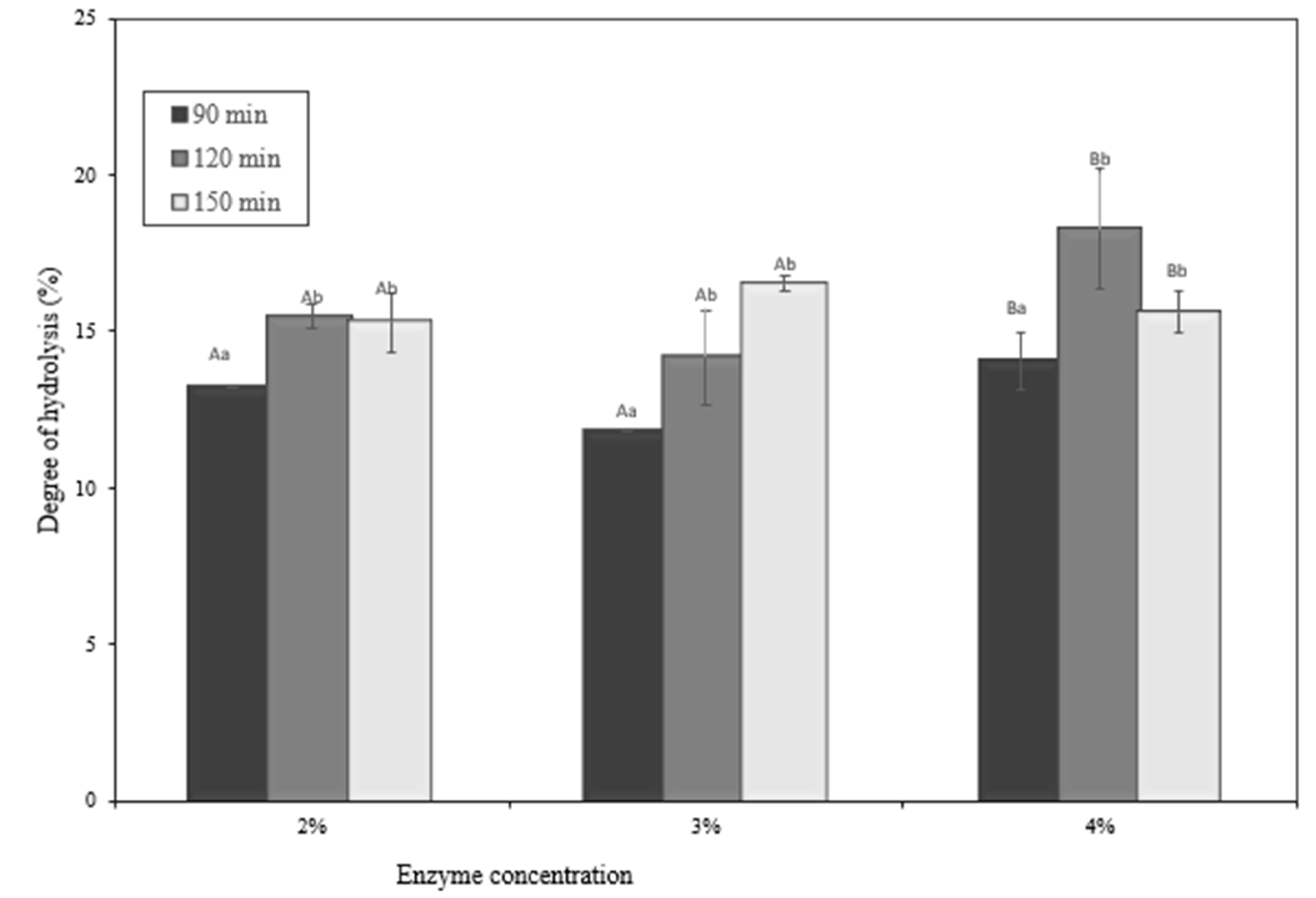
2.7. Degree of Hydrolysis
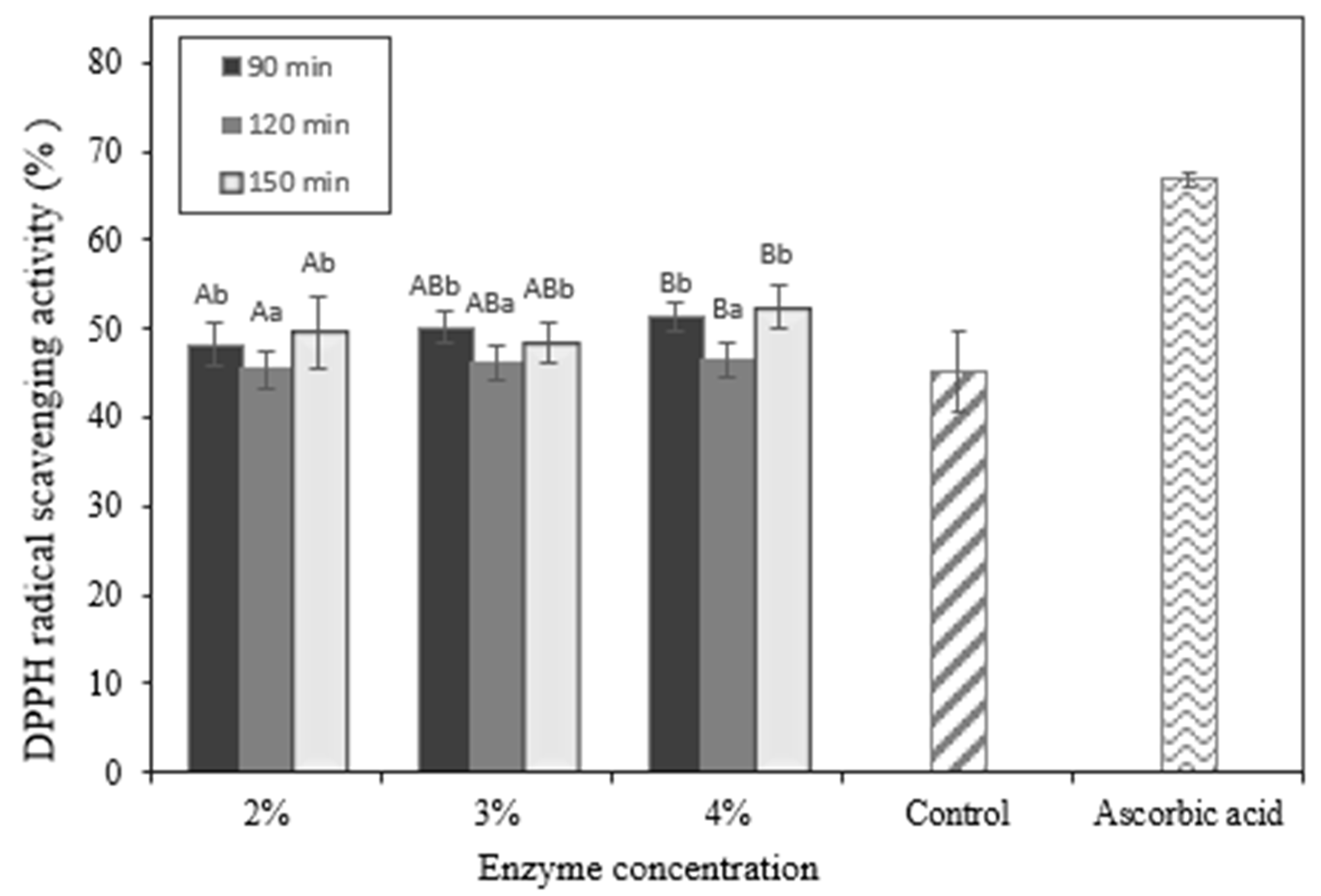
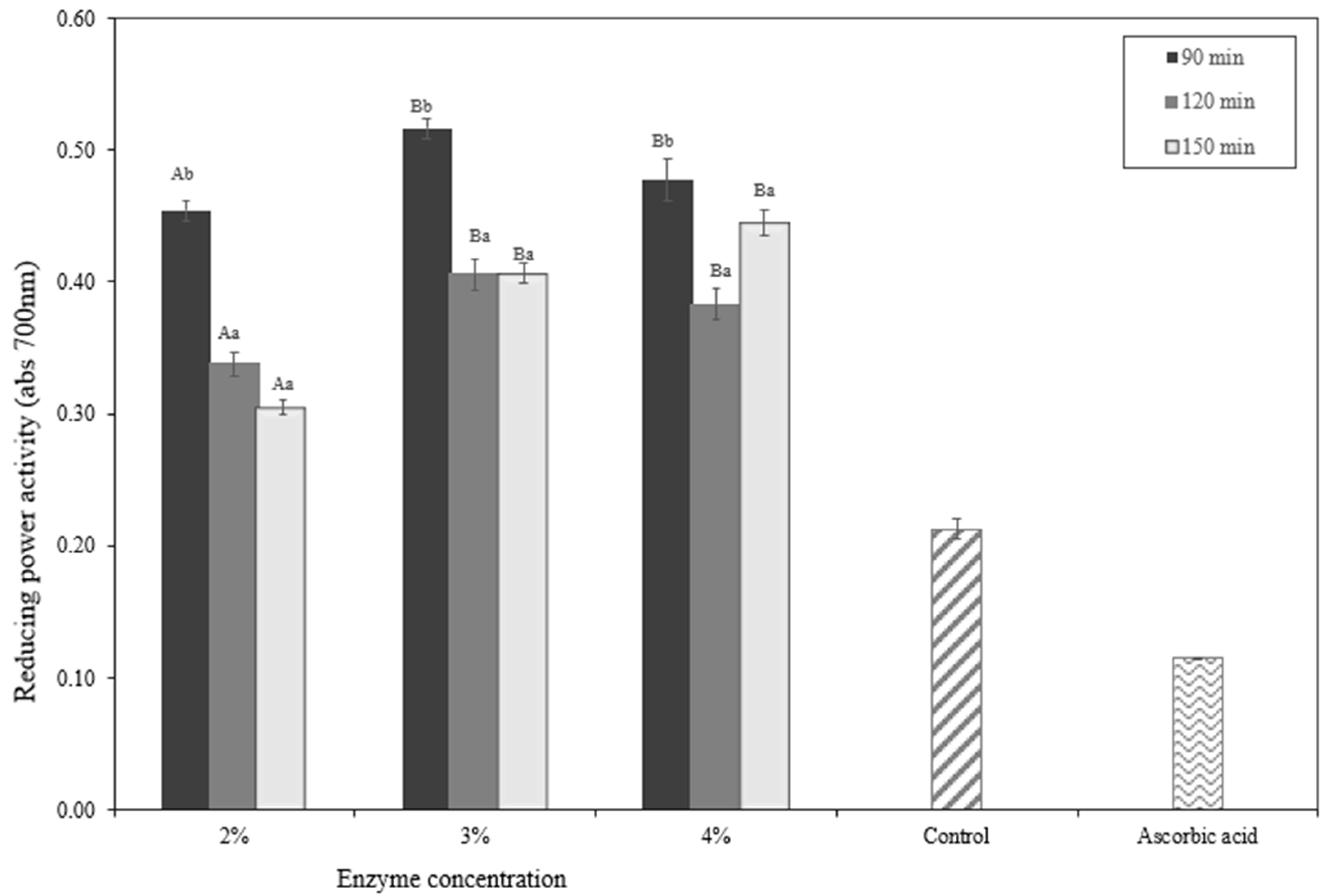
2.8. In Vitro Evaluation of Antioxidant Activity in Okara Hydrolysates
3. Materials and Methods
3.1. Material
3.2. Enzyme and Chemicals
3.3. In Silico Analysis
3.3.1. The Selection of Optimum Protease Based on In Silico Analysis
3.3.2. Peptide Ranking
3.3.3. Toxicity and Sensory Prediction
3.3.4. Molecular Docking against Peptides
3.4. Preparation of Okara Hydrolysates Using Enzymatic Hydrolysis
3.5. Proximate Analysis
3.6. Yield
3.7. Degree of Hydrolysis
3.8. Antioxidant Assays
3.8.1. DPPH Radical Scavenging Activity
3.8.2. Antioxidant Analysis (FRAP Method)
3.9. Determination of the Sequence of the Peptide
3.10. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Vong, W.C.; Liu, S.Q. Biovalorisation of okara (soybean residue) for food and nutrition. Trends Food Sci. Technol. 2016, 52, 139–147. [Google Scholar] [CrossRef]
- Samaranayaka, A.G.P.; Li-Chan, E.C.Y. Food-derived peptidic antioxidants: A review of their production, assessment, and potential applications. J. Funct. Foods 2011, 3, 229–254. [Google Scholar] [CrossRef]
- Umaraw, P.; Munekata, P.E.S.; Verma, A.K.; Barba, F.J.; Singh, V.P.; Kumar, P.; Lorenzo, J.M. Edible films/coating with tailored properties for active packaging of meat, fish and derived products. Trends Food Sci. Technol. 2020, 98, 10–24. [Google Scholar] [CrossRef]
- Gopinatth, V.; Mendez, R.L.; Ballinger, E.; Kwon, J.Y. Therapeutic potential of tuna backbone peptide and its analogs: An in vitro and in silico study. Molecules 2021, 26, 64. [Google Scholar] [CrossRef]
- Kang, N.; Kim, E.A.; Kim, J.; Lee, S.H.; Heo, S.J. Identifying potential antioxidant properties from the viscera of sea snails (turbo cornutus). Mar. Drugs 2021, 19, 67. [Google Scholar] [CrossRef]
- Bhardwaj, V.K.; Oakley, A.; Purohit, R. Mechanistic behavior and subtle key events during DNA clamp opening and closing in T4 bacteriophage. Int. J. Biol. Macromol. 2022, 208, 11–19. [Google Scholar] [CrossRef]
- Kumar, A.; Rajendran, V.; Sethumadhavan, R.; Purohit, R. In silico prediction of a disease-associated STIL mutant and its affect on the recruitment of centromere protein J (CENPJ). FEBS Open Bio 2012, 2, 285–293. [Google Scholar] [CrossRef] [Green Version]
- Purohit, R.; Rajasekaran, R.; Sudandiradoss, C.; George Priya Doss, C.; Ramanathan, K.; Rao, S. Studies on flexibility and binding affinity of Asp25 of HIV-1 protease mutants. Int. J. Biol. Macromol. 2008, 42, 386–391. [Google Scholar] [CrossRef]
- Han, R.; Hernández Álvarez, A.J.; Maycock, J.; Murray, B.S.; Boesch, C. Comparison of alcalase- and pepsin-treated oilseed protein hydrolysates–Experimental validation of predicted antioxidant, antihypertensive and antidiabetic properties. Curr. Res. Food Sci. 2021, 4, 141–149. [Google Scholar] [CrossRef]
- Gupta, S.; Sharma, A.K.; Shastri, V.; Madhu, M.K.; Sharma, V.K. Prediction of anti-inflammatory proteins/peptides: An insilico approach. J. Transl. Med. 2017, 15, 7. [Google Scholar] [CrossRef] [Green Version]
- Zheng, J.; Zheng, X.; Zhao, L.; Yi, J.; Cai, S. Effects and interaction mechanism of soybean 7S and 11S globulins on anthocyanin stability and antioxidant activity during in vitro simulated digestion. Curr. Res. Food Sci. 2021, 4, 543–550. [Google Scholar] [CrossRef]
- Wahyu, D.; Ningrum, A.; Vanidia, N.; Siti, H.; Munawaroh, H.; Susanto, E.; Show, P. Food Hydrocolloids for Health In silico and in vitro assessment of yellowfin tuna skin (Thunnus albacares) hydrolysate antioxidation effect. Food Hydrocoll. Health 2023, 3, 100126. [Google Scholar] [CrossRef]
- Quan, W.; Zhang, C.; Wang, Z.; Zeng, M.; Qin, F.; He, Z.; Chen, J. Assessment antioxidant properties of Torreya grandis protein enzymatic hydrolysates: Utilization of industrial by-products. Food Biosci. 2021, 43, 101325. [Google Scholar] [CrossRef]
- Liu, J.H.; Tian, Y.G.; Wang, Y.; Nie, S.P.; Xie, M.Y.; Zhu, S.; Wang, C.Y.; Zhang, P. Characterization and in vitro antioxidation of papain hydrolysate from black-bone silky fowl (Gallus gallus domesticus Brisson) muscle and its fractions. Food Res. Int. 2011, 44, 133–138. [Google Scholar] [CrossRef]
- Lear, S.; Cobb, S.L. Pep-Calc.com: A set of web utilities for the calculation of peptide and peptoid properties and automatic mass spectral peak assignment. J. Comput. Aided. Mol. Des. 2016, 30, 271–277. [Google Scholar] [CrossRef] [Green Version]
- Munawaroh, H.S.H.; Gumilar, G.G.; Berliana, J.D.; Aisyah, S.; Nuraini, V.A.; Ningrum, A.; Susanto, E.; Martha, L.; Kurniawan, I.; Hidayati, N.A.; et al. In silico proteolysis and molecular interaction of tilapia (Oreochromis niloticus) skin collagen-derived peptides for environmental remediation. Environ. Res. 2022, 212, 113002. [Google Scholar] [CrossRef] [PubMed]
- Ong, J.H.; Koh, J.A.; Cao, H.; Tan, S.A.; Manan, F.A.; Wong, F.C.; Chai, T.T. Purification, identification and characterization of antioxidant peptides from corn silk tryptic hydrolysate: An integrated in vitro-in silico approach. Antioxidants 2021, 10, 1822. [Google Scholar] [CrossRef]
- Basak, T.; Gómez-García, C.J.; Gomila, R.M.; Frontera, A.; Chattopadhyay, S. Hydrogen bond mediated intermolecular magnetic coupling in mononuclear high spin iron(iii) Schiff base complexes: Synthesis, structure and magnetic study with theoretical insight. RSC Adv. 2021, 11, 3315–3323. [Google Scholar] [CrossRef] [PubMed]
- Zhou, P.; Jin, B.; Li, H.; Huang, S.Y. HPEPDOCK: A web server for blind peptide-protein docking based on a hierarchical algorithm. Nucleic Acids Res. 2018, 46, W443–W450. [Google Scholar] [CrossRef] [PubMed]
- Sui, X.; Zhang, Y.; Zhou, W. In vitro and in silico studies of the inhibition activity of anthocyanins against porcine pancreatic α-amylase. J. Funct. Foods 2016, 21, 50–57. [Google Scholar] [CrossRef]
- Bogari, H.A.; Rashied, R.M.H.; Abdelfattah, M.A.O.; Malatani, R.T.; Khinkar, R.M.; Hareeri, R.H.; Wink, M.; Sobeh, M. Euclea divinorum Hiern: Chemical Profiling of the Leaf Extract and Its Antioxidant Activity In Silico, In Vitro and in Caenorhabditis elegans Model. Metabolites 2022, 12, 1031. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Li, T.; Chen, D.; Gu, H.; Mao, X. Identification and molecular docking of antioxidant peptides from hemp seed protein hydrolysates. Lwt 2021, 147, 111453. [Google Scholar] [CrossRef]
- Herowati, R.; Widodo, G.P. Molecular Docking Studies of Chemical Constituents of Tinospora cordifolia on Glycogen Phosphorylase. Procedia Chem. 2014, 13, 63–68. [Google Scholar] [CrossRef] [Green Version]
- Nongonierma, A.B.; FitzGerald, R.J. Enhancing bioactive peptide release and identification using targeted enzymatic hydrolysis of milk proteins. Anal. Bioanal. Chem. 2018, 410, 3407–3423. [Google Scholar] [CrossRef] [PubMed]
- Kallenberg, C.G.M. Antineutrophil Cytoplasmic Autoantibodies with Specificity for Myeloperoxidase, 3rd ed.; Elsevier: Amsterdam, The Netherlands, 2013; ISBN 9780444563781. [Google Scholar]
- Firmansyah, M.; Abduh, M.Y. Production of protein hydrolysate containing antioxidant activity from Hermetia illucens. Heliyon 2019, 5, e02005. [Google Scholar] [CrossRef] [Green Version]
- Shahi, Z.; Sayyed-Alangi, S.Z.; Najafian, L. Effects of enzyme type and process time on hydrolysis degree, electrophoresis bands and antioxidant properties of hydrolyzed proteins derived from defatted Bunium persicum Bioss. press cake. Heliyon 2020, 6, e03365. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.Y.; Hur, S.J. Antihypertensive peptides from animal products, marine organisms, and plants. Food Chem. 2017, 228, 506–517. [Google Scholar] [CrossRef]
- Unnikrishnan, P.; Kizhakkethil, B.P.; George, J.C.; Aliyamveetil Abubacker, Z.; Ninan, G.; Chandragiri Nagarajarao, R. Antioxidant Peptides from Dark Meat of Yellowfin Tuna (Thunnus albacares): Process Optimization and Characterization. Waste Biomass Valorization 2021, 12, 1845–1860. [Google Scholar] [CrossRef]
- Tadesse, S.A.; Emire, S.A. Production and processing of antioxidant bioactive peptides: A driving force for the functional food market. Heliyon 2020, 6, e04765. [Google Scholar] [CrossRef]
- Chaipoot, S.; Punfa, W.; Ounjaijean, S.; Phongphisutthinant, R.; Kulprachakarn, K.; Parklak, W.; Phaworn, L.; Rotphet, P.; Boonyapranai, K. Antioxidant, Anti-Diabetic, Anti-Obesity, and Antihypertensive Properties of Protein Hydrolysate and Peptide Fractions from Black Sesame Cake. Molecules 2023, 28, 211. [Google Scholar] [CrossRef]
- Haddar, A.; Sellimi, S.; Ghannouchi, R.; Alvarez, O.M.; Nasri, M.; Bougatef, A. Functional, antioxidant and film-forming properties of tuna-skin gelatin with a brown algae extract. Int. J. Biol. Macromol. 2012, 51, 477–483. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, C.; Gleddie, S.; Xiao, C.W. Soybean bioactive peptides and their functional properties. Nutrients 2018, 10, 8–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harnedy, P.A.; O’Keeffe, M.B.; FitzGerald, R.J. Fractionation and identification of antioxidant peptides from an enzymatically hydrolysed Palmaria palmata protein isolate. Food Res. Int. 2017, 100, 416–422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chakraborty, P.; Nath, D.; Hoque, M.; Sarkar, P.; Hati, S.; Mishra, B.K. Biopolymer-based antimicrobial coatings for aquatic food products: A review. J. Food Process. Preserv. 2022, 46, e16465. [Google Scholar] [CrossRef]
- Díaz-Calderón, P.; Flores, E.; González-Muñoz, A.; Pepczynska, M.; Quero, F.; Enrione, J. Influence of extraction variables on the structure and physical properties of salmon gelatin. Food Hydrocoll. 2017, 71, 118–128. [Google Scholar] [CrossRef]
- Arshad, M.S.; Hina, G.; Anjum, F.M.; Suleria, H.A.R. Effect of milk-derived bioactive peptides on the lipid stability and functional properties of beef nuggets. Sci. Rep. 2022, 12, 1242. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Protein Precursor | Peptide ID | Sequence | Location | Monoisotop Mass | Chemical Mass |
|---|---|---|---|---|---|---|
| 1 | Glycinin (P04347) | |||||
| 7868 | ADF | (365–367) | 260.1367 | 260.2861 | ||
| 9879 | SVL | (227–229) | 317.1944 | 317.3802 | ||
| 2 | Beta-conglycinin alpha subunit (P0DO16) | |||||
| 8025 | PHF | (453–455) | 399.1901 | 399.4426 | ||
| 8215 | IR | (214–215) | 287.1952 | 287.3576 | ||
| 3 | Beta-conglycinin beta subunit (P25974) | |||||
| 3300 | PHH | (87–89) | 389.1807 | 389.4081 | ||
| 7868 | ADF | (92–94) | 351.1425 | 351.3536 | ||
| 7941 | YYL | (132–134) | 457.2205 | 457.5182 | ||
| 8025 | PHF | (287–289) | 399.1901 | 399.4426 | ||
| 8215 | IR | (53–54) | 287.1952 | 287.3576 | ||
| Amino Acids | Percentage | ||
|---|---|---|---|
| Glycinin(P04347) | Conglycinin Alpha Subunit (P0DO16) | Conglycinin Beta Subunit(P25974) | |
| Alanin (Ala) | 3.9 | 4.6 | 5.2 |
| Arginin (Arg) | 6.4 | 7.9 | 7.1 |
| Asparagin (Asn) | 6.4 | 6.8 | 7.5 |
| Aspartic Acid (Asp) | 4.7 | 4.6 | 4.8 |
| Cysteine (Cys) | 1.6 | 0.8 | 0.2 |
| Glutamin (Gln) | 8.7 | 7.9 | 7.5 |
| Glutamic Acid (Glu) | 8.1 | 13.1 | 8.4 |
| Glycine (Gly) | 7.9 | 4.3 | 4.6 |
| Histidin (His) | 2.9 | 1.3 | 1.8 |
| Isoleusin (Ile) | 3.3 | 5.1 | 5.9 |
| Leusin (Leu) | 7.9 | 8.9 | 10.9 |
| Lysin (Lys) | 3.7 | 6.0 | 4.8 |
| Methionin (Met) | 1.0 | 0.5 | 0.5 |
| Phenylalanin (Phe) | 3.5 | 5.0 | 6.6 |
| Prolin (Pro) | 7.4 | 6.6 | 5.0 |
| Serin (Ser) | 8.3 | 7.4 | 7.5 |
| Threonin (Thr) | 4.1 | 1.8 | 2.5 |
| Tryptofan (Trp) | 0.8 | 0.3 | 0 |
| Tyrosin (Tyr) | 2.9 | 2.5 | 2.7 |
| Valin (Val) | 6.6 | 4.5 | 6.4 |
| No. | Sequence | Mass (g/mol) | Score | Activity | Sensory Evaluation | Toxicity |
|---|---|---|---|---|---|---|
| 1 | PHF | 399.44 | 0.94 | Antioxidant | ND | Non Toxic |
| 2 | ADF | 351.35 | 0.81 | Antioxidant | ND | Non Toxic |
| 3 | YYL | 457.52 | 0.60 | Antioxidant | ND | Non Toxic |
| 4 | PHH | 389.41 | 0.44 | Antioxidant | ND | Non Toxic |
| 5 | IR | 287.36 | 0.33 | Antioxidant | ND | Non Toxic |
| 6 | SVL | 317.38 | 0.13 | Antioxidant | ND | Non Toxic |
| No. | Sequence | Solubility | pI | Peptide Charge (pH 7) | Extinction Coefficient (M−1·cm−1) |
|---|---|---|---|---|---|
| 1 | PHF | Poor | 8.26 | 0 | 0 |
| 2 | ADF | Good | 0.76 | −1 | 0 |
| 3 | YYL | Poor | 3.2 | 0 | 2560 |
| 4 | PHH | Poor | 8.41 | 0.2 | 0 |
| 5 | IR | Good | 10.85 | 1 | 0 |
| 6 | SVL | Poor | 3.37 | 0 | 0 |
| No. | Sequence | Hydrogen Bonding | Hydrophobic Interactions | Electrostatic Interaction |
|---|---|---|---|---|
| 1 | PHF | THR354, SER347, THR262, GLY350 | ALA301,ILE353,LEU404 | |
| 2 | ADF | PHE270, HIS1220, ARG1222, THR1226, SER1225 | ILE266 | ASP 429 |
| 3 | YYL | GLY260, SER347, LEU404, THR354, LYS256, ALA255, LYS249, GLY399 | VAL259, LEU398,ILE353,LEU257 | |
| 4 | PHH | HIS82, LYS57, TRP283, LEU61, ASP59, LEU219 | ARG60,PRO224,PRO285 | |
| 5 | IR | THR1083, THR1082, GLN1040 | ALA1258,VAL1259 | GLU1261 |
| 6 | SVL | LEU257, GLY350, THR354 | ILE353,LYS256 |
| No | Protein Precursors | Sequence | Mw [Da] |
|---|---|---|---|
| 1 | Beta-conglycinin alpha subunit 1 | GSEEEDEDEDEEQDERQFPFPRPPHQK | 3299.4 |
| 2 | Beta-conglycinin alpha subunit 1 | EEDEDEQPRPIPFPRPQPR | 2332.132 |
| 4 | Beta-conglycinin alpha subunit 1 | GEKGSEEEDEDEDEEQDERQFPFPRPPHQK | 3613.55858 |
| 8 | Beta-conglycinin alpha subunit 1 | QFPFPRPPHQK | 1378.73786 |
| 10 | Beta-conglycinin alpha subunit 1 | ESEESEDSELR | 1309.53901 |
| 12 | Beta-conglycinin alpha subunit 1 | ESEESEDSELRR | 1465.64012 |
| 18 | Beta-conglycinin alpha subunit 1 | QFPFPRPPHQKEER | 1792.92416 |
| 19 | Beta-conglycinin alpha subunit 1 | KQEEDEDEEQQRESEESEDSELR | 2853.17767 |
| 29 | Glycinin G4 | WQEQQDEDEDEDEDDEDEQIPSHPPR | 3210.25262 |
| 33 | Glycinin G4 | ADFYNPK | 854.40429 |
| 34 | Kunitz-type trypsin inhibitor | IGENKDAMDGWFR | 1554.70055 |
| 35 | Kunitz-type trypsin inhibitor | IGENKDAMDGWFR | 1538.70564 |
| 39 | AAI domain-containing protein | IMENQSEELEEKQK | 1734.82146 |
| 40 | AAI domain-containing protein | IMENQSEELEEKQKK | 1878.91133 |
| 41 | AAI domain-containing protein | IMENQSEELEEKQKK | 1862.91642 |
| 42 | AAI domain-containing protein OS=Glycine max | IMENQSEELEEKQK | 1750.81637 |
| 43 | AAI domain-containing protein OS=Glycine max | IMENQSEELEEK | 1494.66283 |
| 44 | Sucrose-binding protein | EEEQQEQHEEQDENPYIFEEDKDFETR | 3470.44149 |
| 45 | Sucrose-binding protein | EREEEQQEQHEEQDENPYIFEEDKDFETR | 3755.58519 |
| 47 | Beta-conglycinin beta subunit | NFLAGEKDNVVR | 1361.71719 |
| 51 | Beta-conglycinin beta subunit | ESYFVDAQPQQKEEGSKGR | 2183.03635 |
| 53 | Beta-conglycinin beta subunit | NFLAGEKDNVVR | 1361.71719 |
| 56 | Beta-conglycinin beta subunit | ESYFVDAQPQQKEEGSKGR | 2183.03635 |
| 59 | Lipoxygenase | EIFRTDGEQALK | 1406.72742 |
| 60 | Lipoxygenase | SAWMTDEEFAR | 1342.57323 |
| 61 | Lipoxygenase | ISPIPVLK | 866.57097 |
| 62 | Lipoxygenase | EEELHNLR | 1039.5167 |
| 63 | Lipoxygenase | YREEELHNLRGDGTGER | 2030.96385 |
| 64 | Lipoxygenase | YREEELHNLR | 1358.68114 |
| 67 | Protein disulfide isomerase | EADGIVDYLKK | 1250.66269 |
| 72 | Glycinin G1 | FEELNNDLFR | 1296.62189 |
| 75 | Late embryogenesis abundant protein LEA | KLEIDDDLKLR | 1357.76856 |
| 77 | Lipoxygenase | SAWMTDEEFAR | 1342.57323 |
| 78 | Lipoxygenase | ELFRTDGEQVLK | 1434.75872 |
| 83 | Glucose and ribitol dehydrogenase-like | GHEDRDKDDTLK | 1428.671 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ningrum, A.; Wardani, D.W.; Vanidia, N.; Manikharda; Sarifudin, A.; Kumalasari, R.; Ekafitri, R.; Kristanti, D.; Setiaboma, W.; Munawaroh, H.S.H. Evaluation of Antioxidant Activities from a Sustainable Source of Okara Protein Hydrolysate Using Enzymatic Reaction. Molecules 2023, 28, 4974. https://doi.org/10.3390/molecules28134974
Ningrum A, Wardani DW, Vanidia N, Manikharda, Sarifudin A, Kumalasari R, Ekafitri R, Kristanti D, Setiaboma W, Munawaroh HSH. Evaluation of Antioxidant Activities from a Sustainable Source of Okara Protein Hydrolysate Using Enzymatic Reaction. Molecules. 2023; 28(13):4974. https://doi.org/10.3390/molecules28134974
Chicago/Turabian StyleNingrum, Andriati, Dian Wahyu Wardani, Nurul Vanidia, Manikharda, Achmat Sarifudin, Rima Kumalasari, Riyanti Ekafitri, Dita Kristanti, Woro Setiaboma, and Heli Siti Helimatul Munawaroh. 2023. "Evaluation of Antioxidant Activities from a Sustainable Source of Okara Protein Hydrolysate Using Enzymatic Reaction" Molecules 28, no. 13: 4974. https://doi.org/10.3390/molecules28134974
APA StyleNingrum, A., Wardani, D. W., Vanidia, N., Manikharda, Sarifudin, A., Kumalasari, R., Ekafitri, R., Kristanti, D., Setiaboma, W., & Munawaroh, H. S. H. (2023). Evaluation of Antioxidant Activities from a Sustainable Source of Okara Protein Hydrolysate Using Enzymatic Reaction. Molecules, 28(13), 4974. https://doi.org/10.3390/molecules28134974