


Selenium Protects ARPE-19 and ACBRI 181 Cells against High Glucose-Induced Oxidative Stress

 ,
,
Abstract
:1. Introduction
2. Results
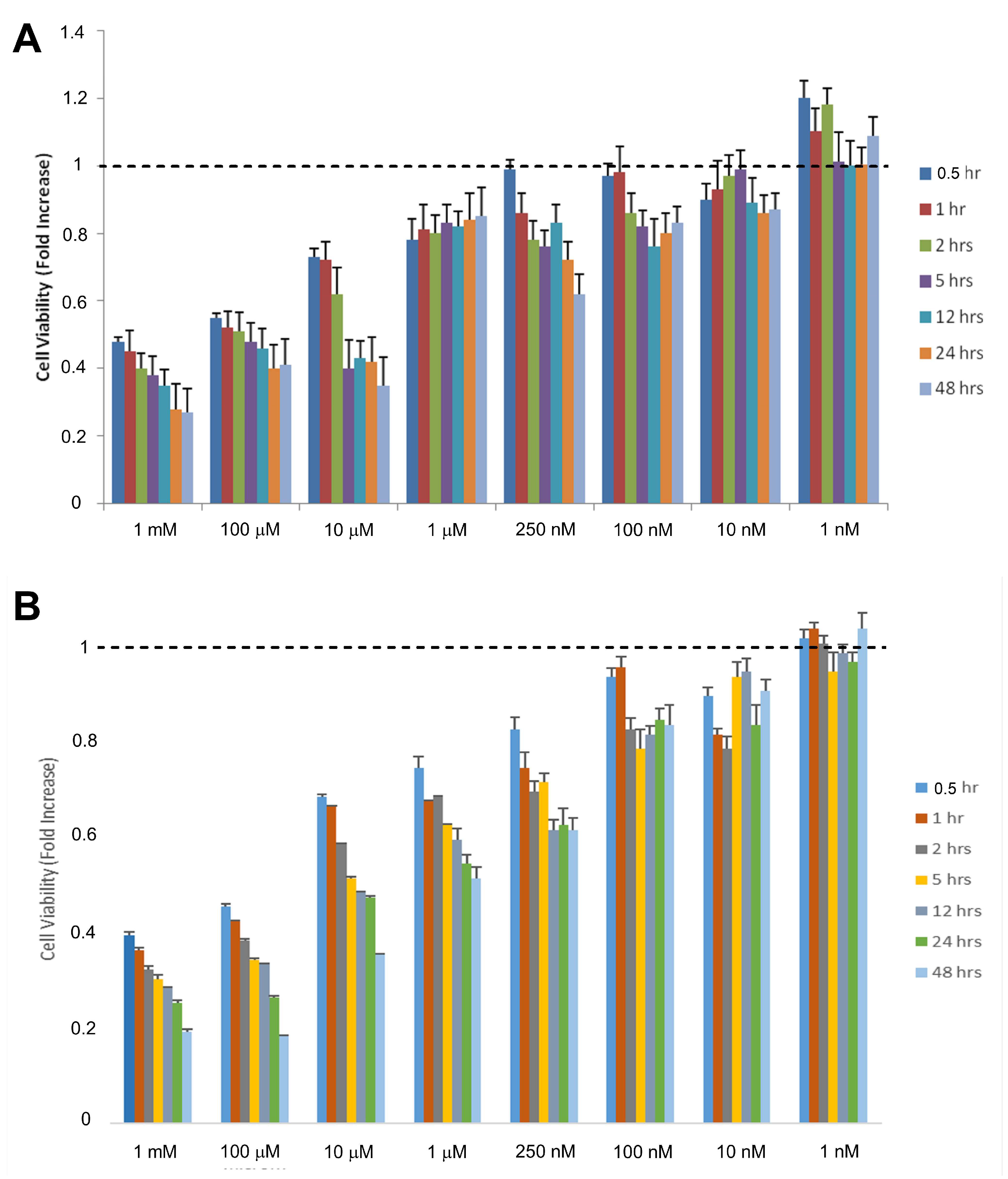
2.1. Toxic and Therapeutic Doses of Se on Cell Viability
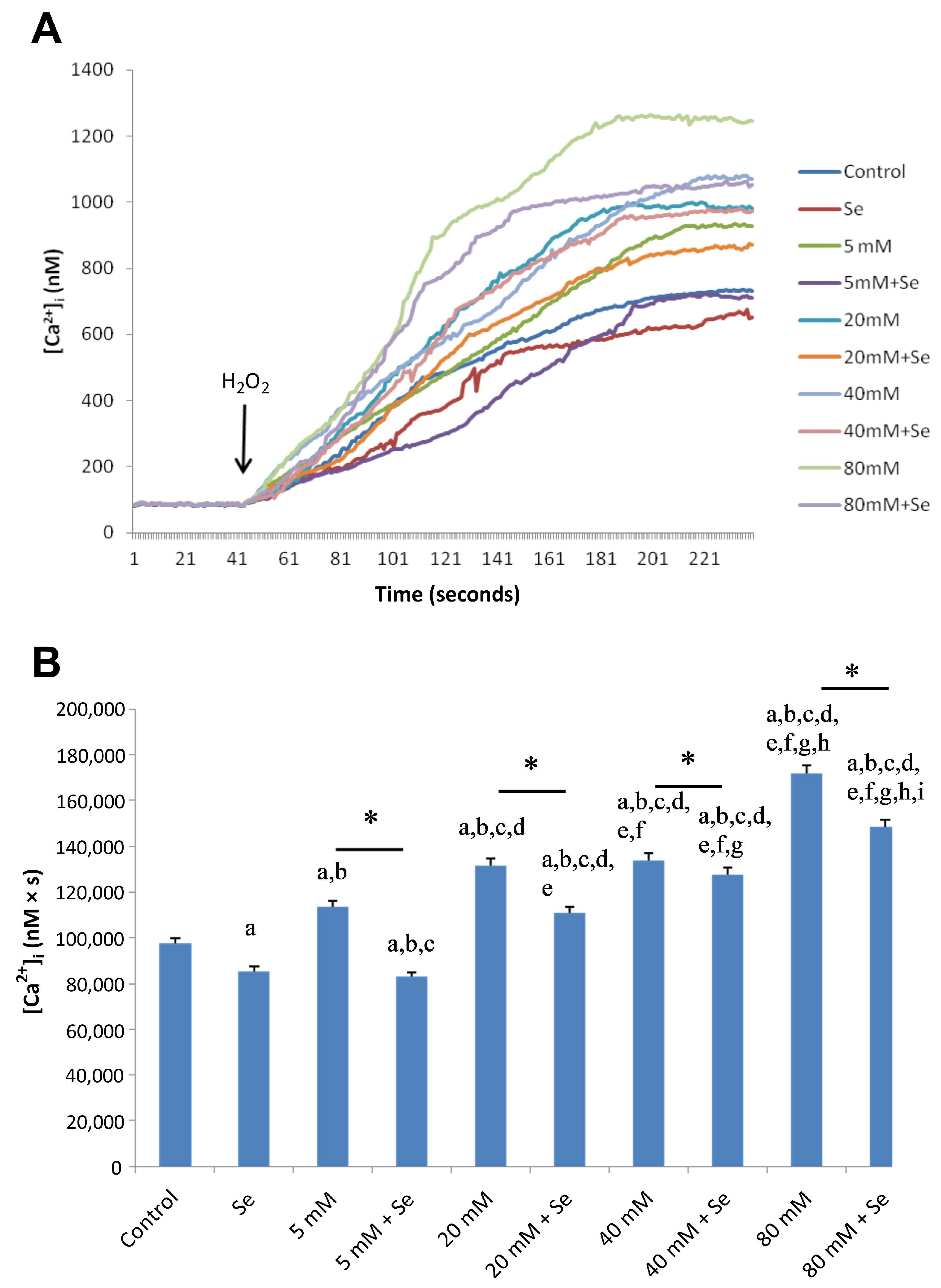
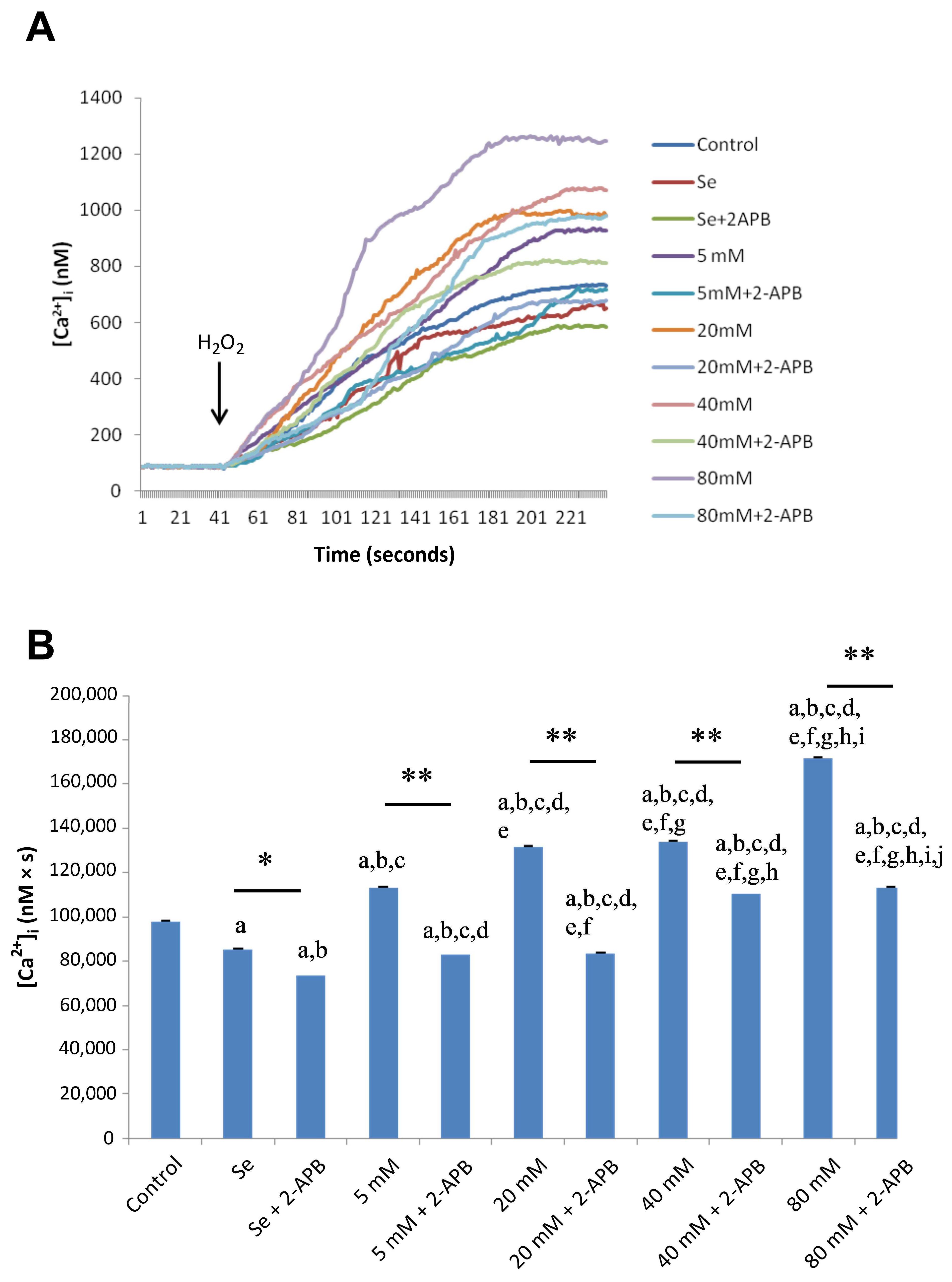
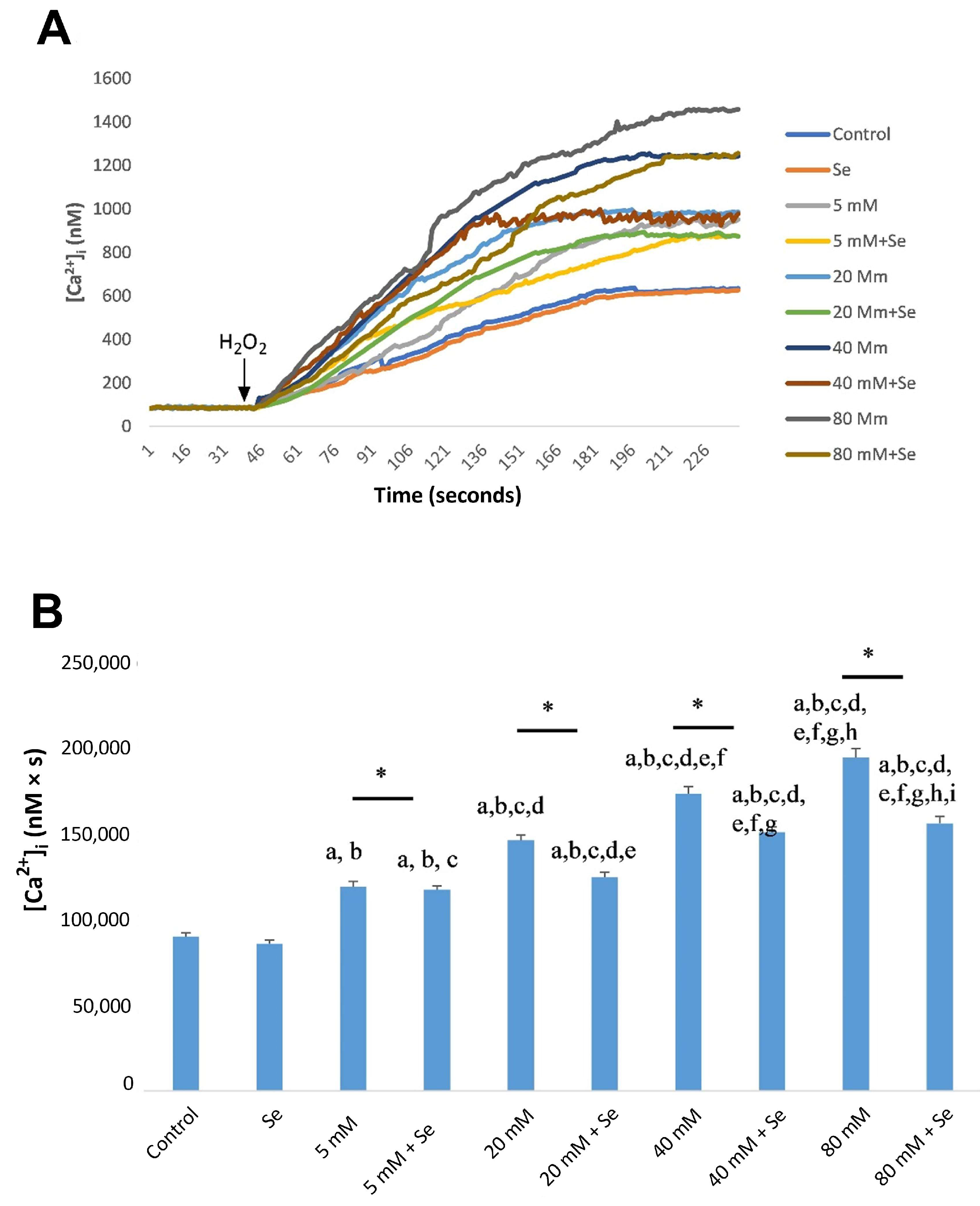
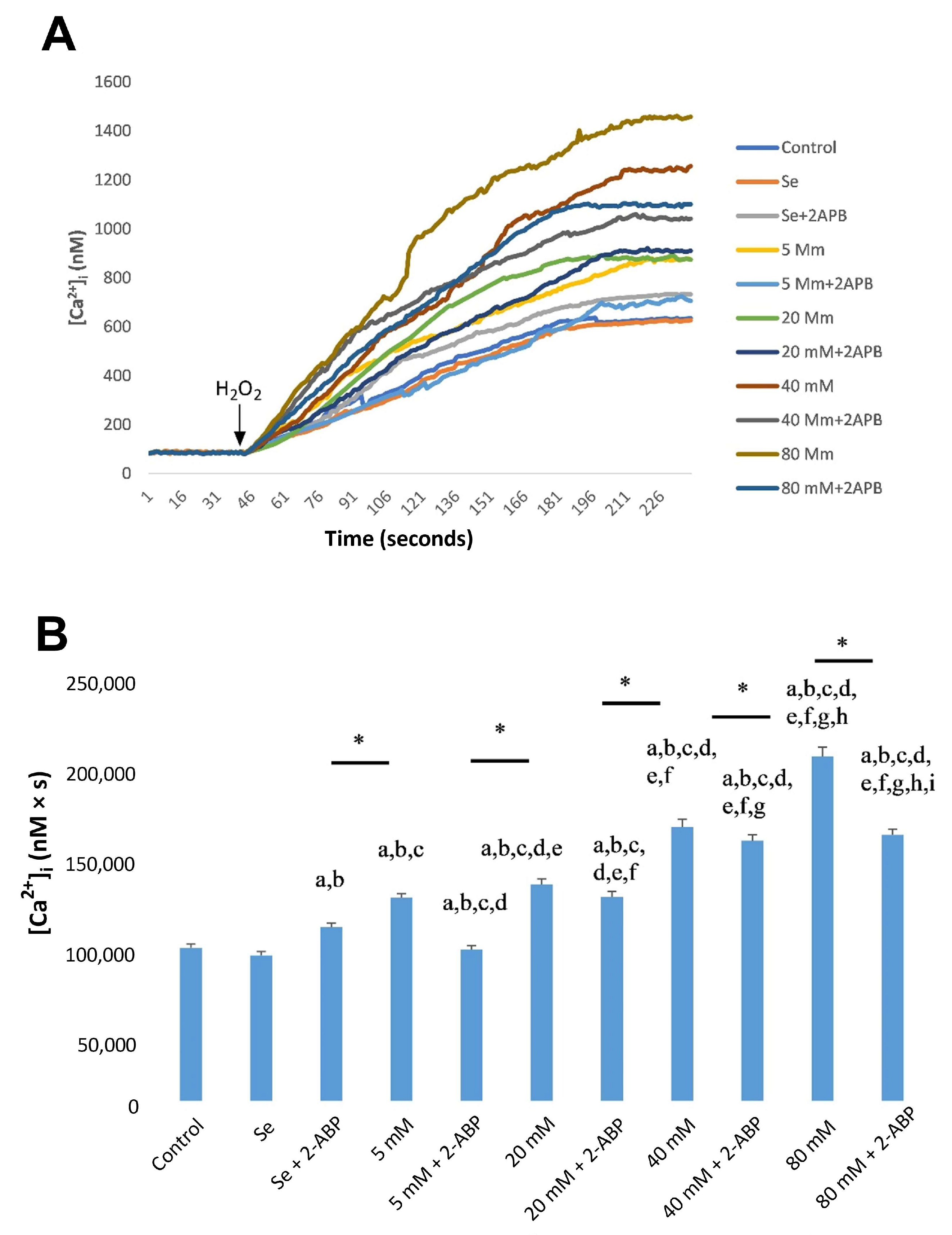
2.2. Effects of Se and HG on Intracellular Ca2+ Concentrations
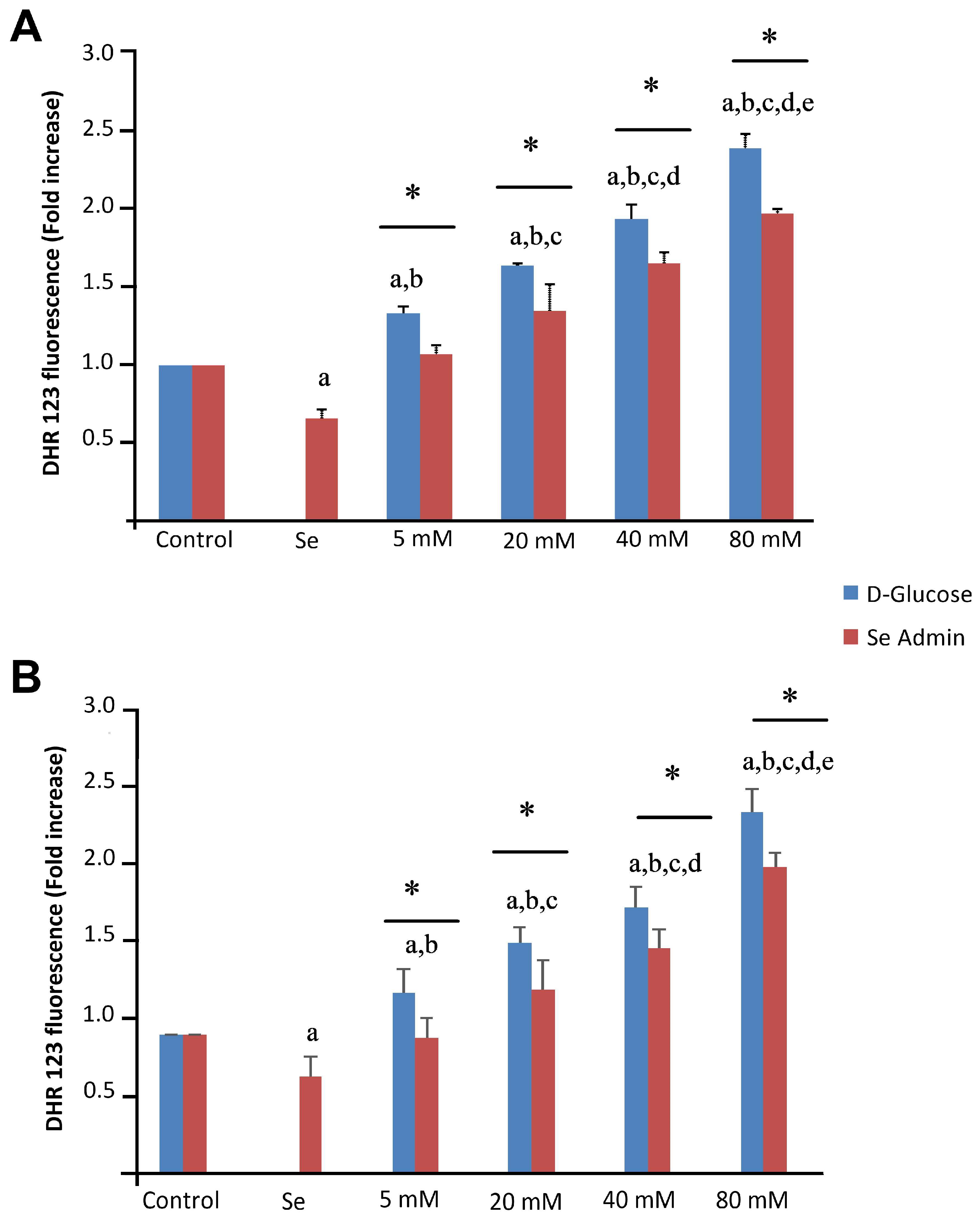
2.3. Effects of Se and HG on Intracellular ROS Production
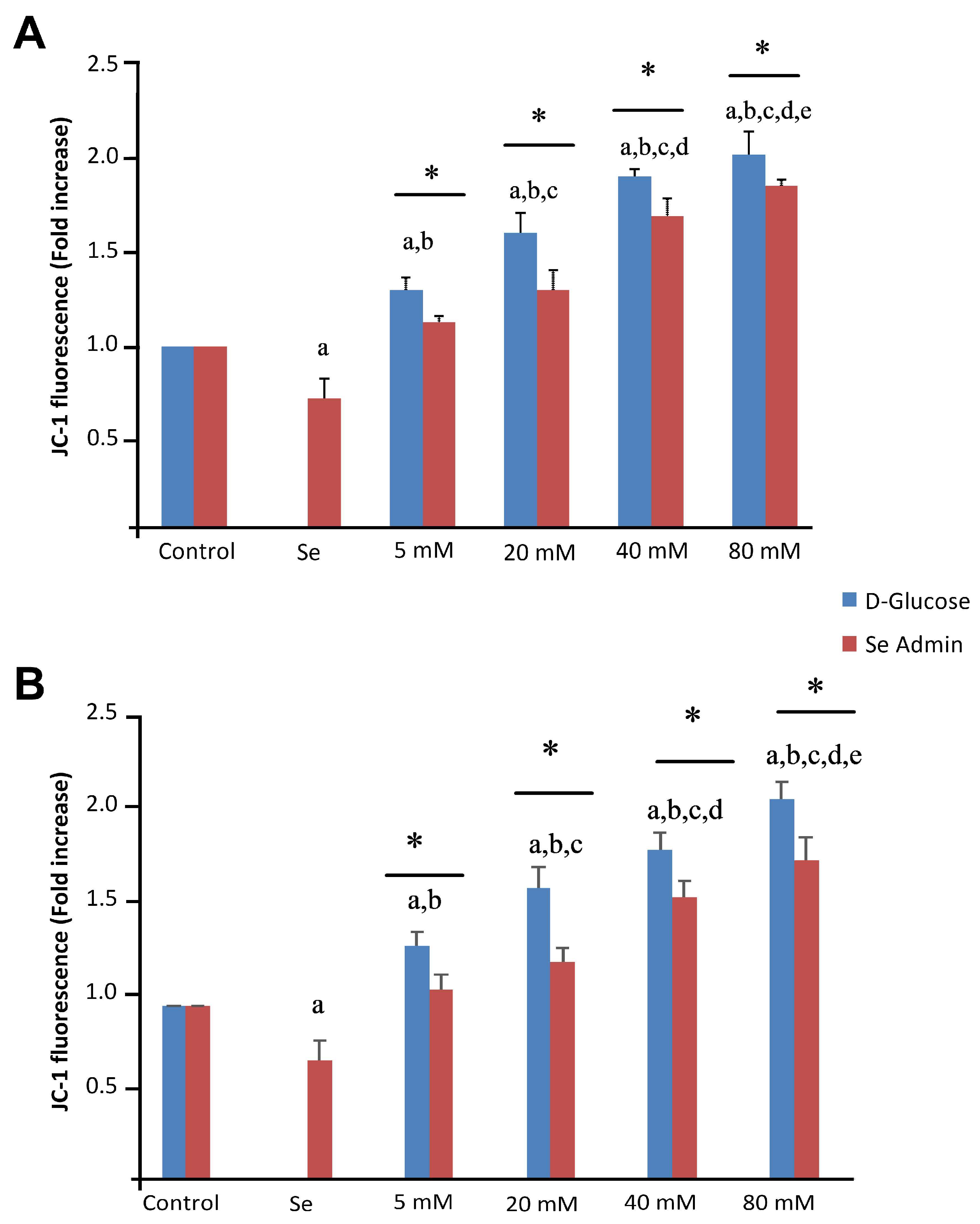
2.4. Effects of Se and HG on Mitochondrial Membrane Depolarization Levels
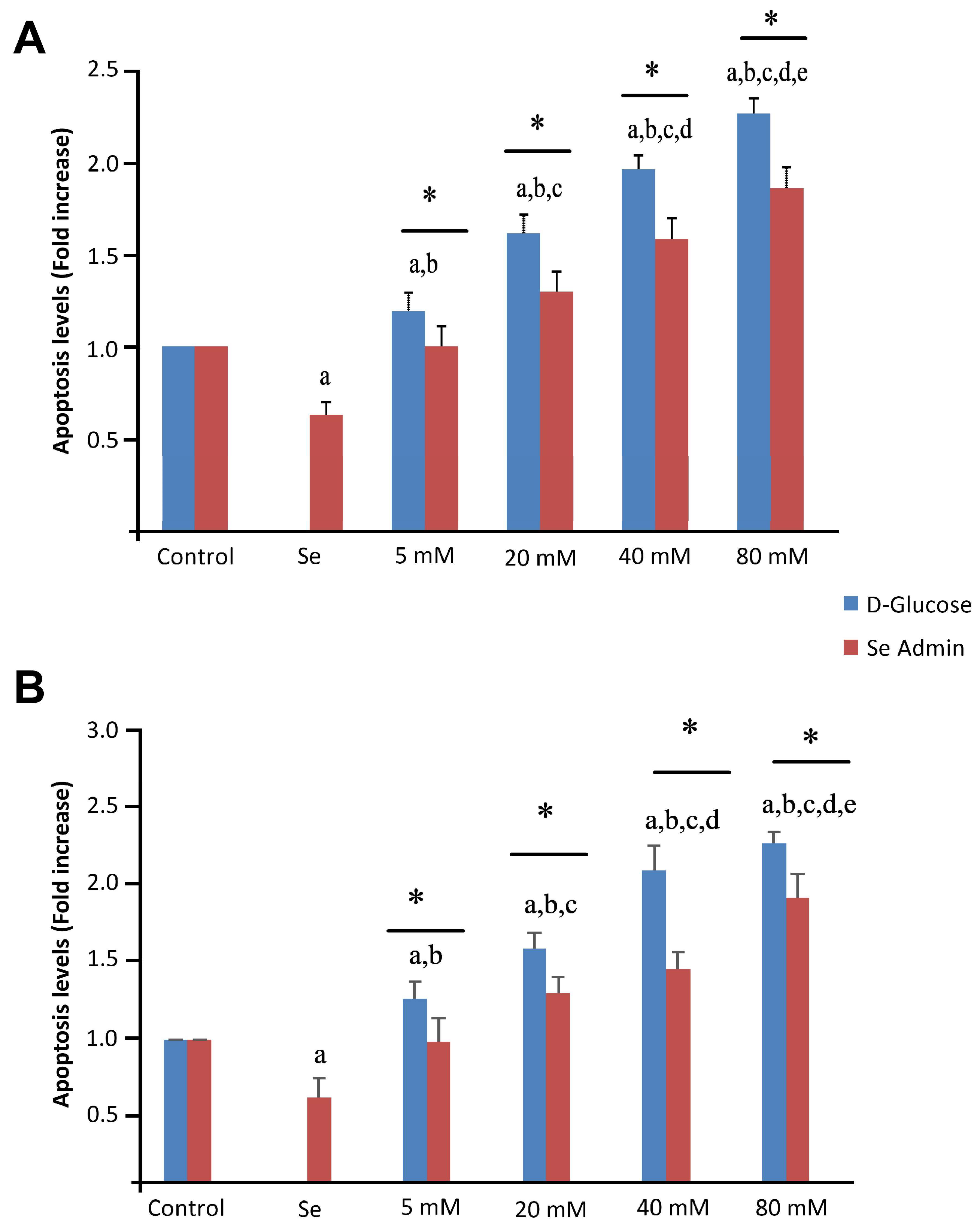
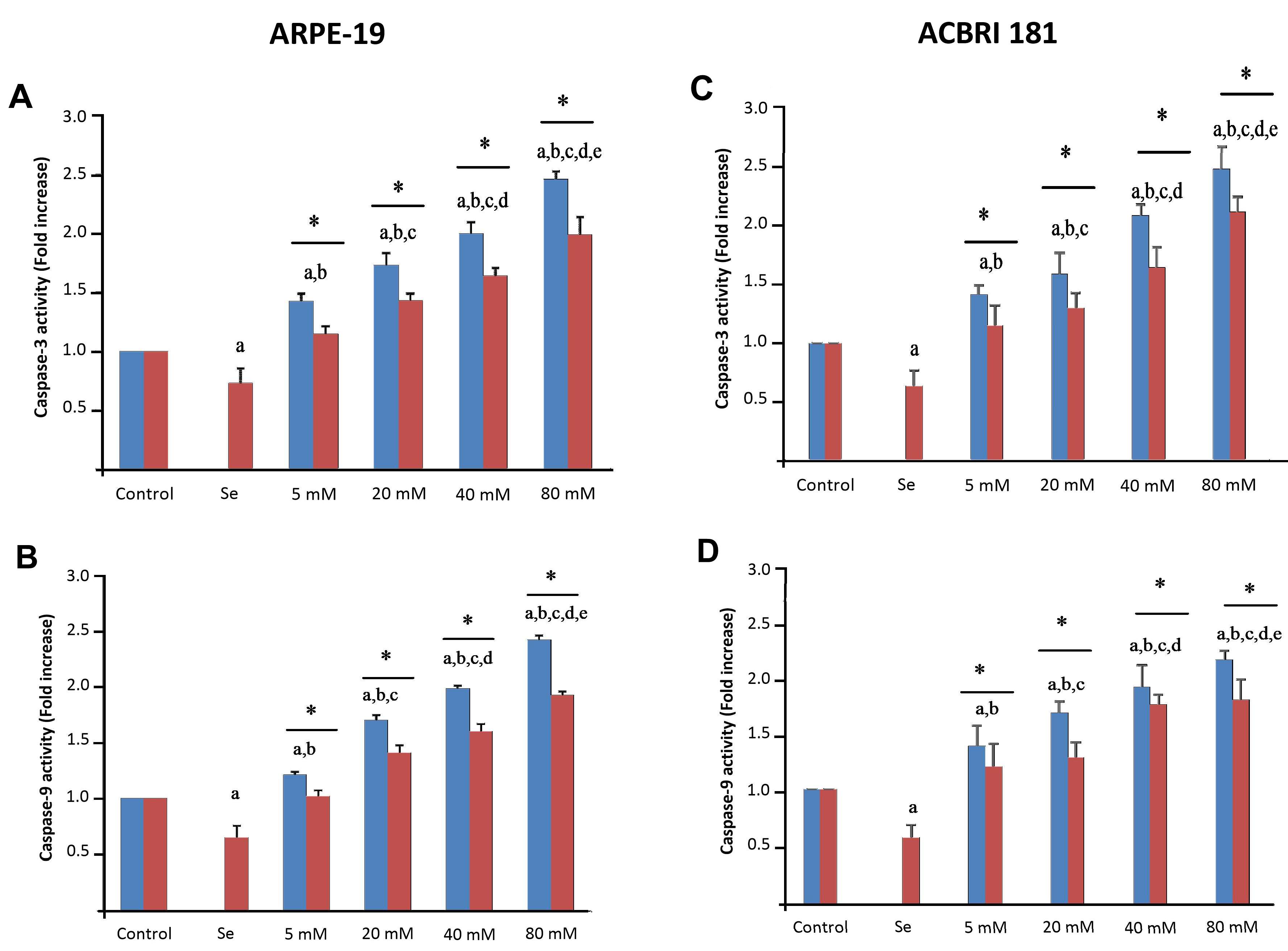
2.5. Effects of Se and HG on Apoptosis and Caspase-3/-9 Activation
2.6. Effects of Se and HG on LP, GSH, GSH-Px, and VEGF Levels
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Cell Culture
4.3. Determination of Selenium Doses by Cell Viability (MTT Assay)
4.4. Determination of Calcium ([Ca2+]i) by Fluorescent Dye
4.5. Measurement of ROS-Sensitive Fluorescence
4.6. Measurement of Lipid Peroxidation (LP) Level
4.7. Measurement of Reduced Glutathione (GSH), Glutathione Peroxidase (GSH-Px), and Protein Assay
4.8. Measurement of Mitochondrial Membrane Potential
4.9. Apoptosis Assay
4.10. Assay for Determination of Caspase Activities
4.11. Measurement of VEGF Levels
4.12. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Flaxman, S.R.; Bourne, R.R.A.; Resnikoff, S.; Ackland, P.; Braithwaite, T.; Cicinelli, M.V.; Das, A.; Jonas, J.B.; Keeffe, J.; Kempen, J.H.; et al. Global causes of blindness and distance vision impairment 1990–2020: A systematic review and meta-analysis. Lancet Glob. Health 2017, 5, e1221–e1234. [Google Scholar] [CrossRef] [Green Version]
- Shivarudrappa, A.H.; Ponesakki, G. Lutein reverses hyperglycemia-mediated blockage of Nrf2 translocation by modulating the activation of intracellular protein kinases in retinal pigment epithelial (ARPE-19) cells. J. Cell Commun. Signal. 2020, 14, 207–221. [Google Scholar] [CrossRef] [PubMed]
- Tenconi, P.E.; Bermúdez, V.; Oresti, G.M.; Giusto, N.M.; Salvador, G.A.; Mateos, M.V. High glucose-induced phospholipase D activity in retinal pigment epithelium cells: New insights into the molecular mechanisms of diabetic retinopathy. Exp. Eye Res. 2019, 184, 243–257. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.; Zhang, G.; Zhang, L.; Hou, Y.P.; Liu, X.L.; Zhang, L. High admission glucose levels increase Fas apoptosis and mortality in patients with acute ST-elevation myocardial infarction: A prospective cohort study. Cardiovasc. Diabetol. 2013, 12, 171. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Xi, X.; Mei, Y.; Zhao, X.; Zhou, L.; Ma, M.; Liu, S.; Zha, X.; Yang, Y. High-glucose induces retinal pigment epithelium mitochondrial pathways of apoptosis and inhibits mitophagy by regulating ROS/PINK1/Parkin signal pathway. Biomed. Pharmacother. 2019, 111, 1315–1325. [Google Scholar] [CrossRef] [PubMed]
- Berridge, M.J. Calcium microdomains: Organization and function. Cell Calcium 2006, 40, 405–412. [Google Scholar] [CrossRef]
- Nazıroğlu, M.; Dikici, D.M.; Dursun, S. Role of oxidative stress and Ca2+ signaling on molecular pathways of neuropathic pain in diabetes: Focus on TRP channels. Neurochem. Res. 2012, 37, 2065–2075. [Google Scholar] [CrossRef]
- Wimmers, S.; Strauss, O. Basal calcium entry in retinal pigment epithelial cells is mediated by TRPC channels. Investig. Ophthalmol. Vis. Sci. 2007, 48, 5767–5772. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Barile, G.; Chang, S.; Hays, A.; Pachydaki, S.; Schiff, W.; Sparrow, J. Apoptosis and cell proliferation in proliferative retinal disorders: PCNA, Ki-67, caspase-3, and PARP expression. Curr. Eye Res. 2005, 30, 395–403. [Google Scholar] [CrossRef]
- Terao, R.; Kaneko, H. Lipid Signaling in Ocular Neovascularization. Int. J. Mol. Sci. 2020, 21, 4758. [Google Scholar] [CrossRef]
- Kahya, M.C.; Nazıroğlu, M.; Övey, İ.S. Modulation of Diabetes-Induced Oxidative Stress, Apoptosis, and Ca2+ Entry Through TRPM2 and TRPV1 Channels in Dorsal Root Ganglion and Hippocampus of Diabetic Rats by Melatonin and Selenium. Mol. Neurobiol. 2017, 54, 2345–2360. [Google Scholar] [CrossRef]
- Sachdeva, R.; Schlotterer, A.; Schumacher, D.; Matka, C.; Mathar, I.; Dietrich, N.; Medert, R.; Kriebs, U.; Lin, J.; Nawroth, P.; et al. TRPC proteins contribute to development of diabetic retinopathy and regulate glyoxalase 1 activity and methylglyoxal accumulation. Mol. Metab. 2018, 9, 156–167. [Google Scholar] [CrossRef]
- Xu, S.Z.; Sukumar, P.; Zeng, F.; Li, J.; Jairaman, A.; English, A.; Naylor, J.; Ciurtin, C.; Majeed, Y.; Milligan, C.J.; et al. TRPC channel activation by extracellular thioredoxin. Nature 2008, 451, 69–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, C.; Seo, H.; Kwak, M.; Jeon, J.; Jang, J.; Jeong, E.M.; Myeong, J.; Hwang, Y.J.; Ha, K.; Kang, M.J.; et al. Increased TRPC5 glutathionylation contributes to striatal neuron loss in Huntington’s disease. Brain J. Neurol. 2015, 138, 3030–3047. [Google Scholar] [CrossRef] [Green Version]
- Ma, R.; Chaudhari, S.; Li, W. Canonical Transient Receptor Potential 6 Channel: A New Target of Reactive Oxygen Species in Renal Physiology and Pathology. Antioxid. Redox Signal. 2016, 25, 732–748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, W.; Zhu, J.; Li, S.; Tang, C.; Zhao, Q.; Zhang, J. Selenium supplementation protects against oxidative stress-induced cardiomyocyte cell cycle arrest through activation of PI3K/AKT. Met. Integr. Biomet. Sci. 2020, 12, 1965–1978. [Google Scholar] [CrossRef]
- Watson, M.; van Leer, L.; Vanderlelie, J.J.; Perkins, A.V. Selenium supplementation protects trophoblast cells from oxidative stress. Placenta 2012, 33, 1012–1019. [Google Scholar] [CrossRef] [Green Version]
- Uguz, A.C.; Cig, B.; Espino, J.; Bejarano, I.; Naziroglu, M.; Rodriguez, A.B.; Pariente, J.A. Melatonin potentiates chemotherapy-induced cytotoxicity and apoptosis in rat pancreatic tumor cells. J. Pineal Res. 2012, 53, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Argun, M.; Tök, L.; Uğuz, A.C.; Çelik, Ö.; Tök, Ö.Y.; Naziroğlu, M. Melatonin and amfenac modulate calcium entry, apoptosis, and oxidative stress in ARPE-19 cell culture exposed to blue light irradiation (405 nm). Eye 2014, 28, 752–760. [Google Scholar] [CrossRef] [Green Version]
- Simó, R.; Villarroel, M.; Corraliza, L.; Hernández, C.; Garcia-Ramírez, M. The retinal pigment epithelium: Something more than a constituent of the blood-retinal barrier--implications for the pathogenesis of diabetic retinopathy. J. Biomed. Biotechnol. 2010, 2010, 190724. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.Z.; Le, Y.Z. Significance of outer blood-retina barrier breakdown in diabetes and ischemia. Investig. Ophthalmol. Vis. Sci. 2011, 52, 2160–2164. [Google Scholar] [CrossRef] [PubMed]
- Maedler, K.; Spinas, G.A.; Lehmann, R.; Sergeev, P.; Weber, M.; Fontana, A.; Kaiser, N.; Donath, M.Y. Glucose induces beta-cell apoptosis via upregulation of the Fas receptor in human islets. Diabetes 2001, 50, 1683–1690. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.; Wang, W.; Ma, J.; Ye, P.; Wang, K. High glucose induces mitochondrial dysfunction and apoptosis in human retinal pigment epithelium cells via promoting SOCS1 and Fas/FasL signaling. Cytokine 2016, 78, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Brini, M.; Carafoli, E. The plasma membrane Ca2+ ATPase and the plasma membrane sodium calcium exchanger cooperate in the regulation of cell calcium. Cold Spring Harb. Perspect. Biol. 2011, 3, a004168. [Google Scholar] [CrossRef]
- Kim, J.H.; Kim, J.H.; Jun, H.O.; Yu, Y.S.; Min, B.H.; Park, K.H.; Kim, K.W. Protective effect of clusterin from oxidative stress-induced apoptosis in human retinal pigment epithelial cells. Investig. Ophthalmol. Vis. Sci. 2010, 51, 561–566. [Google Scholar] [CrossRef] [PubMed]
- Zou, X.; Feng, Z.; Li, Y.; Wang, Y.; Wertz, K.; Weber, P.; Fu, Y.; Liu, J. Stimulation of GSH synthesis to prevent oxidative stress-induced apoptosis by hydroxytyrosol in human retinal pigment epithelial cells: Activation of Nrf2 and JNK-p62/SQSTM1 pathways. J. Nutr. Biochem. 2012, 23, 994–1006. [Google Scholar] [CrossRef]
- Hanus, J.; Zhang, H.; Wang, Z.; Liu, Q.; Zhou, Q.; Wang, S. Induction of necrotic cell death by oxidative stress in retinal pigment epithelial cells. Cell Death Dis. 2013, 4, e965. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.; Zhang, Q.; Steinle, J.J. Beta-adrenergic receptor agonist decreases VEGF levels through altered eNOS and PKC signaling in diabetic retina. Growth Factors 2015, 33, 192–199. [Google Scholar] [CrossRef] [Green Version]
- Song, H.B.; Jun, H.O.; Kim, J.H.; Yu, Y.S.; Kim, K.W.; Kim, J.H. Suppression of protein kinase C-ζ attenuates vascular leakage via prevention of tight junction protein decrease in diabetic retinopathy. Biochem. Biophys. Res. Commun. 2014, 444, 63–68. [Google Scholar] [CrossRef]
- Titchenell, P.M.; Lin, C.M.; Keil, J.M.; Sundstrom, J.M.; Smith, C.D.; Antonetti, D.A. Novel atypical PKC inhibitors prevent vascular endothelial growth factor-induced blood-retinal barrier dysfunction. Biochem. J. 2012, 446, 455–467. [Google Scholar] [CrossRef] [Green Version]
- Wilson, C.H.; Ali, E.S.; Scrimgeour, N.; Martin, A.M.; Hua, J.; Tallis, G.A.; Rychkov, G.Y.; Barritt, G.J. Steatosis inhibits liver cell store-operated Ca2+ entry and reduces ER Ca2+ through a protein kinase C-dependent mechanism. Biochem. J. 2015, 466, 379–390. [Google Scholar] [CrossRef] [PubMed]
- González de Vega, R.; García, M.; Fernández-Sánchez, M.L.; González-Iglesias, H.; Sanz-Medel, A. Protective effect of selenium supplementation following oxidative stress mediated by glucose on retinal pigment epithelium. Met. Integr. Biometal Sci. 2018, 10, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Öztürk Kurt, H.P.; Karagöz Özen, D.S.; Genç, İ.; Erdem, M.; Demirdağ, M.D. Comparison of selenium levels between diabetic patients with and without retinopathy: Selenium levels and diabetic retinopathy. J. Surg. Med. 2023, 7, 58–62. [Google Scholar] [CrossRef]
- Wang, C.; Ran, R.; Jin, X.; Zhu, X. Plasma and vitreous selenium concentrations in patients with type 2 diabetes and diabetic retinopathy. Medicine 2022, 101, e30877. [Google Scholar] [CrossRef] [PubMed]
- Hoque, B.; Shi, Z. Association between selenium intake, diabetes and mortality in adults: Findings from National Health and Nutrition Examination Survey (NHANES) 2003-2014. Br. J. Nutr. 2022, 127, 1098–1105. [Google Scholar] [CrossRef]
- Dunn, K.C.; Aotaki-Keen, A.E.; Putkey, F.R.; Hjelmeland, L.M. ARPE-19, a human retinal pigment epithelial cell line with differentiated properties. Exp. Eye Res. 1996, 62, 155–169. [Google Scholar] [CrossRef]
- Grynkiewicz, G.; Poenie, M.; Tsien, R.Y. A new generation of Ca2+ indicators with greatly improved fluorescence properties. J. Biol. Chem. 1985, 260, 3440–3450. [Google Scholar] [CrossRef]
- Altinkiliç, S.; Naziroğlu, M.; Uğuz, A.C.; Ozcankaya, R. Fish oil and antipsychotic drug risperidone modulate oxidative stress in PC12 cell membranes through regulation of cytosolic calcium ion release and antioxidant system. J. Membr. Biol. 2010, 235, 211–218. [Google Scholar] [CrossRef]
- Uğuz, A.C.; Naziroğlu, M.; Espino, J.; Bejarano, I.; González, D.; Rodríguez, A.B.; Pariente, J.A. Selenium modulates oxidative stress-induced cell apoptosis in human myeloid HL-60 cells through regulation of calcium release and caspase-3 and -9 activities. J. Membr. Biol. 2009, 232, 15–23. [Google Scholar] [CrossRef]
- Placer, Z.A.; Cushman, L.L.; Johnson, B.C. Estimation of product of lipid peroxidation (malonyl dialdehyde) in biochemical systems. Anal. Biochem. 1966, 16, 359–364. [Google Scholar] [CrossRef]
- Sedlak, J.; Lindsay, R.H. Estimation of total, protein-bound, and nonprotein sulfhydryl groups in tissue with Ellman’s reagent. Anal. Biochem. 1968, 25, 192–205. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, R.A.; Burk, R.F. Glutathione peroxidase activity in selenium-deficient rat liver. Biochem. Biophys. Res. Commun. 1976, 71, 952–958. [Google Scholar] [CrossRef] [PubMed]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters/Groups | Control | Se | 5 mM | 5 mM + Se | 20 mM | 20 mM + Se | 40 mM | 40 mM + Se | 80 mM | 80 mM + Se |
|---|---|---|---|---|---|---|---|---|---|---|
| LP (µMol/g protein) | 13.26 ± 1.24 | 10.76 ± 1.34 a | 15.89 ± 1.23 a,b | 13.87 ± 1.38 b,c | 25.93 ± 2.35 a,b,c,d | 20.16 ± 2.41 a,b,c,d,e | 26.18 ± 1.76 a,b,c,d,e,f | 24.22 ± 2.43 a,b,c,d,e,f,g | 28.53 ± 2.47 a,b,c,d,e,f,g,h | 27.11 ± 2.46 a,b,c,d,e,f,g,h,i |
| GSH (µMol/g protein) | 6.16 ± 0.44 | 14.12 ± 0.44 a | 5.48 ± 1.41 a,b | 7.47 ± 1.64 a,b,c | 4.33 ± 1.26 a,b,c,d | 5.78 ± 1.36 a,b,d,e | 4.18 ± 1.27 a,b,c,d,f | 5.92 ± 1.24 a,b,d,e,g | 3.78 ± 0.64 a,b,c,d,e,f,g,h | 5.17 ± 1.57 a,b,d,e,g,i |
| GSH-Px (IU/g protein) | 15.62 ± 2.14 | 19.43 ± 2.34 a | 14.42 ± 2.32 b | 16.01 ± 2.78 b,c | 18.65 ± 1.84 a,c,d | 17.71 ± 2.36 a,b,c,d | 20.82 ± 2.36 a,c,d,e,f | 18.76 ± 1.77 a,c,d,f | 22.52 ± 2.18 a,b,c,d,e,f,h | 20.43 ± 4.41 a,c,d,e,f,h,i |
| VEGF (pg/mL) | 41.18 ± 2.48 | 33.25 ± 3.21 a | 54.17 ± 5.16 a,b | 50.16 ± 5.24 a,b,c | 63.45 ± 4.28 a,b,c,d | 57.13 ± 3.47 a,b,c,d,e | 67.28 ± 5.18 a,b,c,d,e,f | 58.14 ± 4.27 a,b,c,d,e,g | 69.38 ± 4.37 a,b,c,d,e,f,h | 61.21 ± 3.18 a,b,c,d,e,f,g,h,i |
| Parameters/Groups | Control | Se | 5 mM | 5 mM + Se | 20 mM | 20 mM + Se | 40 mM | 40 mM + Se | 80 mM | 80 mM + Se |
|---|---|---|---|---|---|---|---|---|---|---|
| LP (µMol/g protein) | 14.37 ± 1.19 | 9.81 ± 1.28 a | 16.74 ± 1.14 a,b | 14.87 ± 1.38 b,c | 27.41 ± 1.21 a,b,c,d | 21.13 ± 1.85 a,b,c,d,e | 28.41 ± 1.24 a,b,c,d,e,f | 25.28 ± 2.14 a,b,c,d,e,f,g | 27.13 ± 2.47 a,b,c,d,e,f,g,h | 29.14 ± 2.13 a,b,c,d,e,f,g,h,i |
| GSH (µMol/g protein) | 5.18 ± 0.38 | 13.23 ± 0.52 a | 5.52 ± 1.24 a,b | 7.56 ± 1.28 a,b,c | 5.13 ± 1.17 a,b,c,d | 5.59 ± 1.24 a,b,d,e | 4.23 ± 1,34 a,b,c,d,f | 6.13 ± 1.41 a,b,d,e,g | 3.41 ± 0.27 a,b,c,d,e,f,g,h | 5.39 ± 1.21 a,b,d,e,g,i |
| GSH-Px (IU/g protein) | 14.27 ± 2.11 | 20.37 ± 1.87 a | 14.93 ± 2.15 b | 17.13 ± 2.14 b,c | 10.48 ± 1.23 a,c,d | 17.51 ± 2.36 a,b,c,d | 11.67 ± 2.56 a,c,d,e,f | 17.83 ± 1.41 a,c,d,f | 12.41 ± 2.07 a,b,c,d,e,f,h | 20.43 ± 2.12 a,c,d,e,f,h,i |
| VEGF (pg/mL) | 39.22 ± 2.13 | 31.17 ± 2.17 a | 52.29 ± 4.24 a,b | 48.13 ± 4.16 a,b,c | 60.17 ± 3.17 a,b,c,d | 55.17 ± 4.41 a,b,c,d,e | 65.17 ± 4.24 a,b,c,d,e,f | 55.43 ± 4.19 a,b,c,d,e,g | 67.51 ± 3.81 a,b,c,d,e,f,h | 57.21 ± 4.15 a,b,c,d,e,f,g,h,i |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bardak, H.; Uğuz, A.C.; Bardak, Y.; Rocha-Pimienta, J.; Delgado-Adámez, J.; Espino, J. Selenium Protects ARPE-19 and ACBRI 181 Cells against High Glucose-Induced Oxidative Stress. Molecules 2023, 28, 5961. https://doi.org/10.3390/molecules28165961
Bardak H, Uğuz AC, Bardak Y, Rocha-Pimienta J, Delgado-Adámez J, Espino J. Selenium Protects ARPE-19 and ACBRI 181 Cells against High Glucose-Induced Oxidative Stress. Molecules. 2023; 28(16):5961. https://doi.org/10.3390/molecules28165961
Chicago/Turabian StyleBardak, Handan, Abdülhadi Cihangir Uğuz, Yavuz Bardak, Javier Rocha-Pimienta, Jonathan Delgado-Adámez, and Javier Espino. 2023. "Selenium Protects ARPE-19 and ACBRI 181 Cells against High Glucose-Induced Oxidative Stress" Molecules 28, no. 16: 5961. https://doi.org/10.3390/molecules28165961
APA StyleBardak, H., Uğuz, A. C., Bardak, Y., Rocha-Pimienta, J., Delgado-Adámez, J., & Espino, J. (2023). Selenium Protects ARPE-19 and ACBRI 181 Cells against High Glucose-Induced Oxidative Stress. Molecules, 28(16), 5961. https://doi.org/10.3390/molecules28165961