
Secondary Metabolomic Analysis and In Vitro Bioactivity Evaluation of Stems Provide a Comprehensive Comparison between Dendrobium chrysotoxum and Dendrobium thyrsiflorum


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
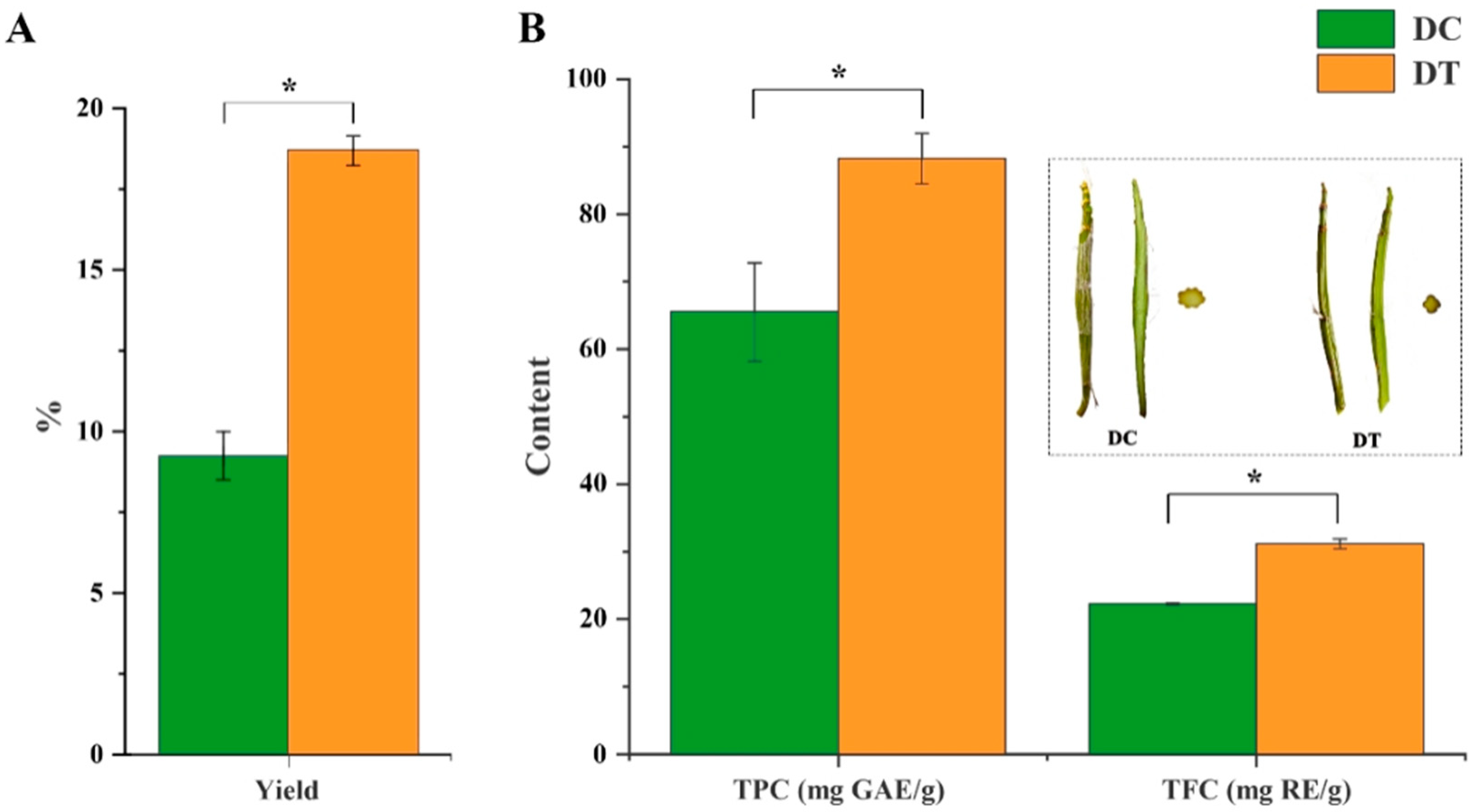
2.1. Yield, Total Phenolic, and Flavonoid Content of Stem Extracts
2.2. Secondary Metabolite Profiling of Stem Extracts
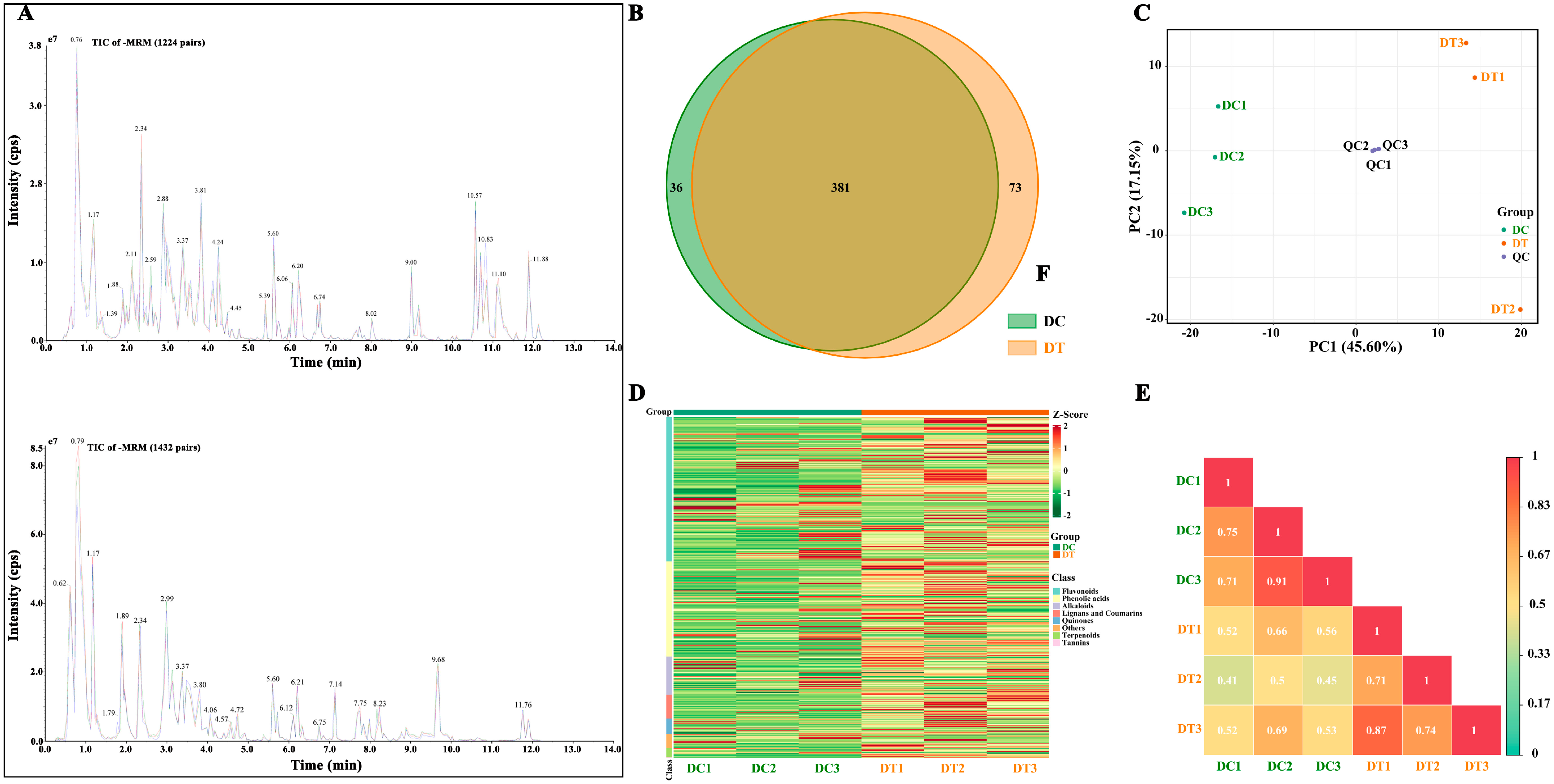
2.2.1. Overall Secondary Metabolite Profiling
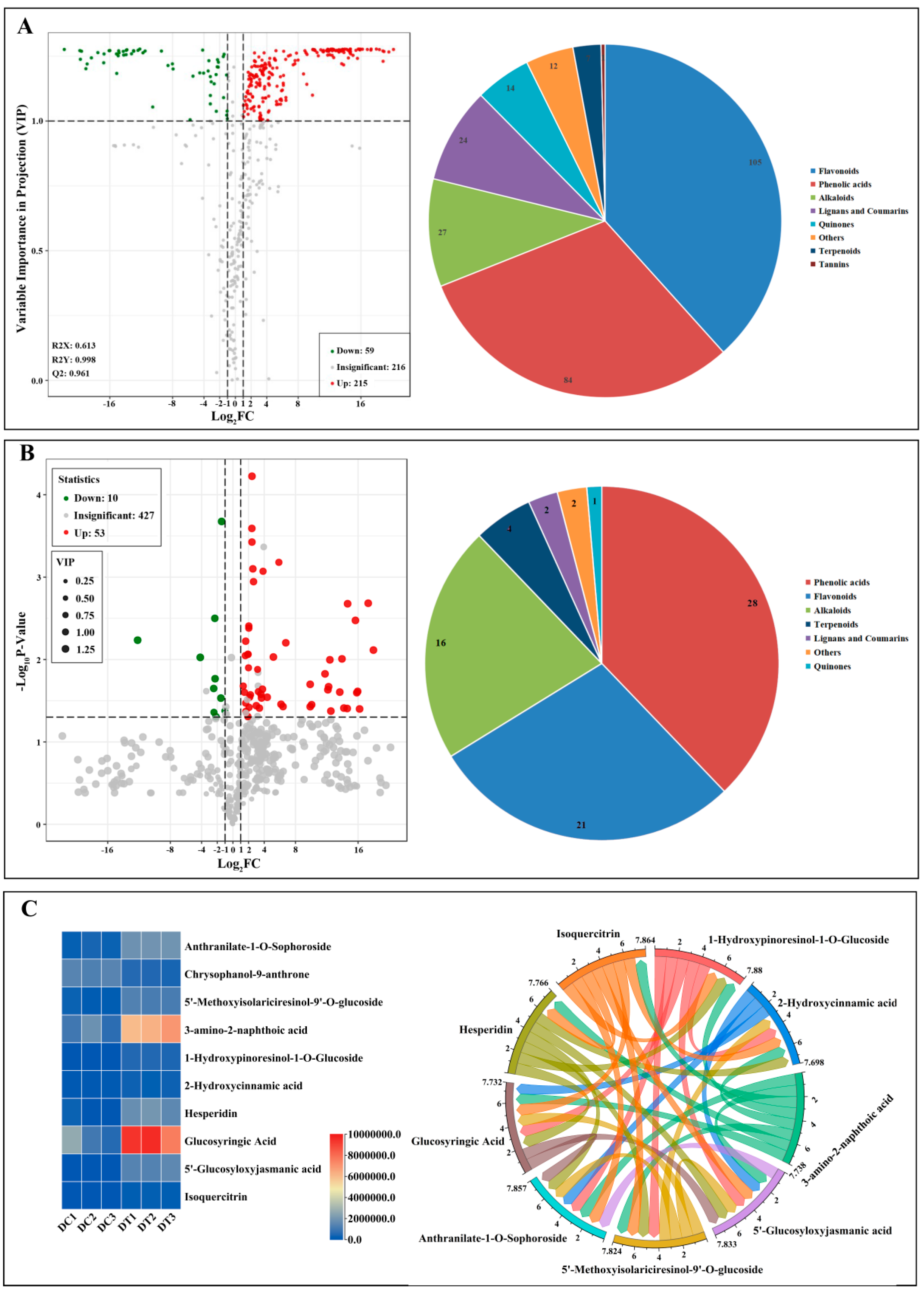
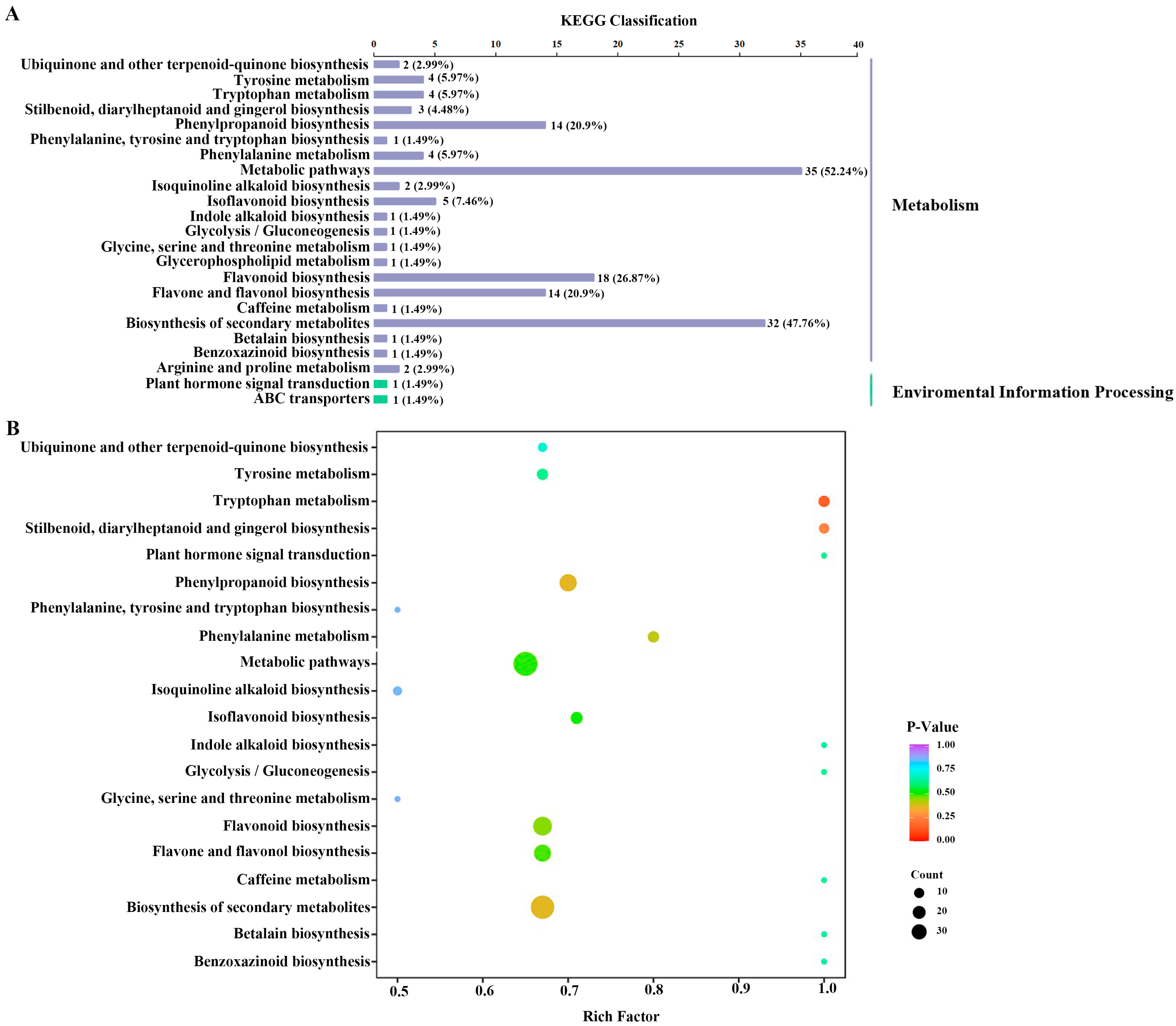
2.2.2. Determination and Metabolic Pathways of Differential Metabolites
2.2.3. Determination of the Key Health-Promoting Constituents
2.3. In Vitro Bioactivity of the Stem Extracts
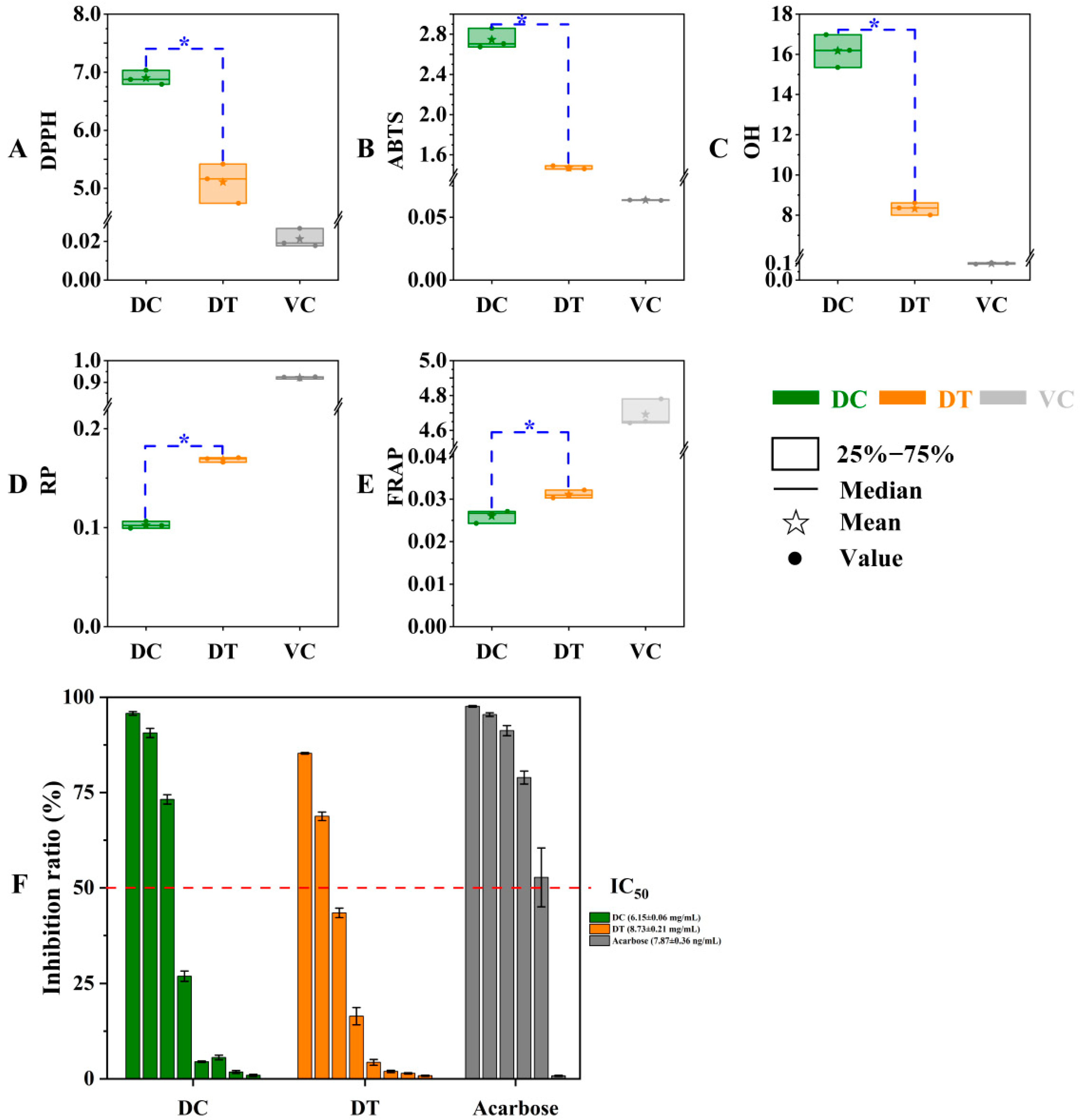
2.3.1. In Vitro Antioxidant Activity
2.3.2. α-Glucosidase Inhibition Assay
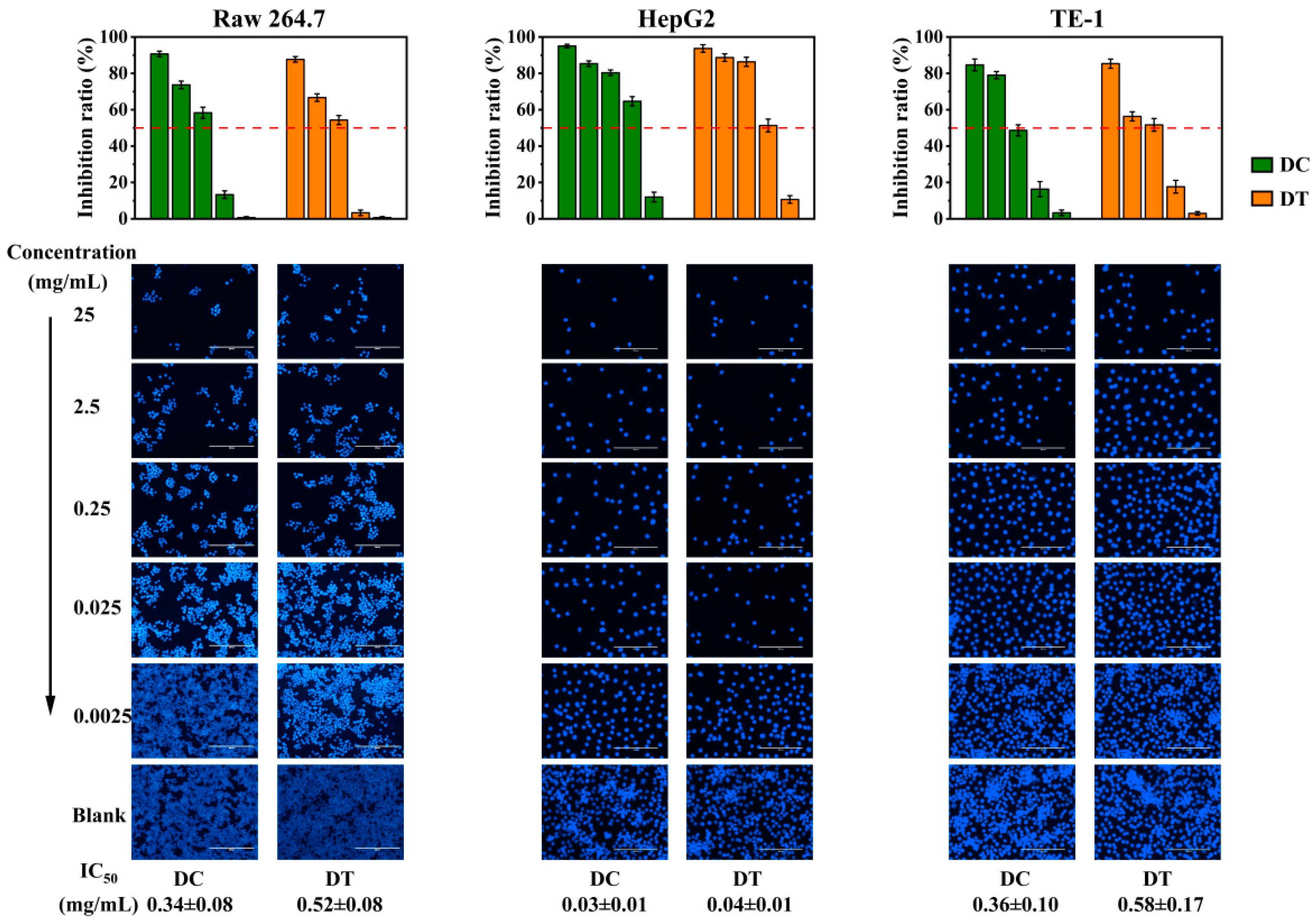
2.3.3. Cytotoxicity of Stem Extracts
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Collection of Plant Material and Sample Preparation
4.3. Determination of Total Phenolics and Total Flavonoids in the Extracts
4.4. Metabolomic Profiling of the Extracts
4.4.1. The Qualitative and Quantitative Analysis of Metabolites
4.4.2. Determination and Annotation of Differential Metabolites
4.4.3. Determination of the Key Health-Promoting Constituents
4.5. Determination of In Vitro Bioactivities for the Extracts
4.5.1. Determination of In Vitro Antioxidant Assays
4.5.2. α-Glucosidase Inhibition Assay
4.5.3. Determination of Cytotoxicity
4.5.4. Calculation of the Half Maximal Inhibitory Concentration
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Cheng, J.; Dang, P.P.; Zhao, Z.; Yuan, L.C.; Zhou, Z.H.; Wolf, D.; Luo, Y.B. An assessment of the Chinese medicinal Dendrobium industry: Supply, demand and sustainability. J. Ethnopharmacol. 2019, 229, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Kanlayavattanakul, M.; Lourith, N.; Chaikul, P. Biological activity and phytochemical profiles of Dendrobium: A new source for specialty cosmetic materials. Ind. Crops Prod. 2018, 120, 61–70. [Google Scholar] [CrossRef]
- Wang, K.-W.; Yang, C.; Yan, S.-N.; Wang, H.; Cao, X.-J.; Cheng, Y. Dendrobium hancockii polysaccharides, structure characterization, modification, antioxidant and antibacterial activity. Ind. Crops Prod. 2022, 188, 115565. [Google Scholar] [CrossRef]
- Yan, L.; Zhang, Z.; Liu, Y.; Ren, S.; Zhu, Z.; Wei, L.; Feng, J.; Duan, T.; Sun, X.; Xie, T.; et al. Anticancer activity of erianin: Cancer-specific target prediction based on network pharmacology. Front. Mol. Biosci. 2022, 9, 862932. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Li, X.; Wan, W.; Yang, Q.; Ma, W.; Chen, D.; Hu, J.; Chen, C.Y.O.; Wei, X. Gigantol from Dendrobium chrysotoxum Lindl. binds and inhibits aldose reductase gene to exert its anti-cataract activity: An in vitro mechanistic study. J. Ethnopharmacol. 2017, 198, 255–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.-p.; Wang, Y.-j.; Zhang, J. Cytotoxic phenanthrenes and fluorenones from Dendrobium thyrsiflorum. Chem. Nat. Compd. 2016, 52, 880–882. [Google Scholar] [CrossRef]
- Zhu, B.; Luo, X.; Gao, Z.; Feng, Q.; Hu, L.; Weng, Q. The complete chloroplast genome sequence of Dendrobium thyrsiflorum (Orchidaceae) and its phylogenetic analysis. Mitochondrial DNA Part B Resour. 2019, 4, 4085–4086. [Google Scholar] [CrossRef] [Green Version]
- Ning, S.; Ying, L.U.; Minghua, Q.I.U. Studies on immunomodulation of polysaccharide from Dendrobium thyrsiflorum Rchb.f. Nat. Prod. Res. Dev. 2006, 18, 445–448. [Google Scholar]
- Deng, H.; Wu, G.; Zhang, R.; Yin, Q.; Xu, B.; Zhou, L.; Chen, Z. Comparative nutritional and metabolic analysis reveals the taste variations during yellow rambutan fruit maturation. Food Chem. X 2023, 17, 100580. [Google Scholar] [CrossRef]
- Wei, J.; Li, S.; Su, T.; Zhao, J.; Jiang, Y.; Zubarev, Y.A.; Bi, Y. Phenolic compositions and antioxidant activities of Hippophae tibetana and H. rhamnoides ssp. sinensis berries produced in Qinghai-Tibet Plateau. Food Chem. X 2022, 15, 100397. [Google Scholar] [CrossRef]
- Yin, Q.-C.; Ji, J.-B.; Zhang, R.-H.; Duan, Z.-W.; Xie, H.; Chen, Z.; Hu, F.-C.; Deng, H. Identification and verification of key taste components in wampee using widely targeted metabolomics. Food Chem. X 2022, 13, 100261. [Google Scholar] [CrossRef] [PubMed]
- Zou, S.; Wu, J.; Shahid, M.Q.; He, Y.; Lin, S.; Liu, Z.; Yang, X. Identification of key taste components in loquat using widely targeted metabolomics. Food Chem. 2020, 323, 126822. [Google Scholar] [CrossRef] [PubMed]
- Hong, Q.; Chen, G.; Wang, Z.; Chen, X.; Kan, J. Effects of different thermal processing methods on bioactive components, phenolic compounds, and antioxidant activities of Qingke (highland hull-less barley). Food Sci. Hum. Wellness 2023, 12, 119–129. [Google Scholar] [CrossRef]
- Liu, Y.; Huang, W.; Zhang, C.; Li, C.; Fang, Z.; Zeng, Z.; Hu, B.; Chen, H.; Wu, W.; Wang, T.; et al. Targeted and untargeted metabolomic analyses and biological activity of Tibetan tea. Food Chem. 2022, 384, 132517. [Google Scholar] [CrossRef]
- Ma, Y.; Li, X.; Hou, L.-X.; Wei, A.-Z. Extraction solvent affects the antioxidant, antimicrobial, cholinesterase and HepG2 human hepatocellular carcinoma cell inhibitory activities of Zanthoxylum bungeanum pericarps and the major chemical components. Ind. Crops Prod. 2019, 142, 111872. [Google Scholar] [CrossRef]
- Ali, I.B.E.; Tajini, F.; Boulila, A.; Jebri, M.-A.; Boussaid, M.; Messaoud, C.; Sebai, H. Bioactive compounds from Tunisian Pelargonium graveolens (L’Her.) essential oils and extracts: Alpha-amylase and acethylcholinesterase inhibitory and antioxidant, antibacterial and phytotoxic activities. Ind. Crops Prod. 2020, 158, 112951. [Google Scholar]
- Pintac, D.; Majkic, T.; Torovic, L.; Orcic, D.; Beara, I.; Simin, N.; Mimica-Dukic, N.; Lesjak, M. Solvent selection for efficient extraction of bioactive compounds from grape pomace. Ind. Crops Prod. 2018, 111, 379–390. [Google Scholar] [CrossRef]
- Zhan, X.; Qian, Y.; Mao, B. Metabolic profiling of terpene diversity and the response of prenylsynthase-terpene synthase genes during biotic and abiotic stresses in Dendrobium catenatum. Int. J. Mol. Sci. 2022, 23, 6398. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, Q.; Wei, F.; Liu, N. Progressive study of effects of erianin on anticancer activity. Oncotargets Ther. 2019, 12, 5457–5465. [Google Scholar] [CrossRef] [Green Version]
- Jarocka-Karpowicz, I.; Markowska, A. Therapeutic potential of jasmonic acid and its derivatives. Int. J. Mol. Sci. 2021, 22, 8437. [Google Scholar] [CrossRef]
- Valentova, K.; Vrba, J.; Bancirova, M.; Ulrichova, J.; Kren, V. Isoquercitrin: Pharmacology, toxicology, and metabolism. Food Chem. Toxicol. 2014, 68, 267–282. [Google Scholar] [CrossRef] [PubMed]
- Song, C.; Jiao, C.; Jin, Q.; Chen, C.; Cai, Y.; Lin, Y. Metabolomics analysis of nitrogen-containing metabolites between two Dendrobium plants. Physiol. Mol. Biol. Plants 2020, 26, 1425–1435. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.; Tang, H.; Song, J.; Long, J.; Zhang, L.; Li, X. Chrysophanol: A review of its pharmacology, toxicity and pharmacokinetics. J. Pharm. Pharmacol. 2019, 71, 1475–1487. [Google Scholar] [CrossRef] [Green Version]
- Cao, X.; Yang, H.; Shang, C.; Ma, S.; Liu, L.; Cheng, J. The roles of auxin biosynthesis yucca gene family in plants. Int. J. Mol. Sci. 2019, 20, 6343. [Google Scholar] [CrossRef] [Green Version]
- Zengin, G.; Atasagun, B.; Aumeeruddy, M.Z.; Saleem, H.; Mollica, A.; Bahadori, M.B.; Mahomoodally, M.F. Phenolic profiling and in vitro biological properties of two Lamiaceae species (Salvia modesta and Thymus argaeus): A comprehensive evaluation. Ind. Crops Prod. 2019, 128, 308–314. [Google Scholar] [CrossRef]
- Ge, X.; Jing, L.; Zhao, K.; Su, C.; Zhang, B.; Zhang, Q.; Han, L.; Yu, X.; Li, W. The phenolic compounds profile, quantitative analysis and antioxidant activity of four naked barley grains with different color. Food Chem. 2021, 335, 127655. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Gu, X.; Shi, Y. A review on lignin antioxidants: Their sources, isolations, antioxidant activities and various applications. Int. J. Biol. Macromol. 2022, 210, 716–741. [Google Scholar] [CrossRef] [PubMed]
- Xie, G.; Jin, S.; Li, H.; Ai, M.; Han, F.; Dai, Y.; Tao, W.; Zhu, Y.; Zhao, Y.; Qin, M. Chemical constituents and antioxidative, anti-inflammatory and anti-proliferative activities of wild and cultivated Corydalis saxicola. Ind. Crops Prod. 2021, 169, 113647. [Google Scholar] [CrossRef]
- Han, B.; Sun, H.; Yang, X.; Wang, T.; He, C.; Zhao, K.; An, S.; Guo, C.; Niu, D.; Xue, X.; et al. Ultrasound-assisted enzymatic extraction of Corni Fructus alpha-glucosidase inhibitors improves insulin resistance in HepG2 cells. Food Funct. 2021, 12, 9808–9819. [Google Scholar] [CrossRef]
- Wang, X.; Deng, Y.; Xie, P.; Liu, L.; Zhang, C.; Cheng, J.; Zhang, Y.; Liu, Y.; Huang, L.; Jiang, J. Novel bioactive peptides from ginkgo biloba seed protein and evaluation of their alpha-glucosidase inhibition activity. Food Chem. 2023, 404, 134481. [Google Scholar] [CrossRef]
- Hong, Y.; Liao, X.; Chen, Z. Screening and characterization of potential alpha-glucosidase inhibitors from Cercis chinensis Bunge fruits using ultrafiltration coupled with HPLC-ESI-MS/MS. Food Chem. 2022, 372, 131316. [Google Scholar] [CrossRef] [PubMed]
- Ye, C.; Zhang, R.; Dong, L.; Chi, J.; Huang, F.; Dong, L.; Zhang, M.; Jia, X. alpha-Glucosidase inhibitors from brown rice bound phenolics extracts (BRBPE): Identification and mechanism. Food Chem. 2022, 372, 131306. [Google Scholar] [CrossRef] [PubMed]
- Zhai, D.; Lv, X.; Chen, J.; Peng, M.; Cai, J. Recent research progress on natural stilbenes in Dendrobium species. Molecules 2022, 27, 7233. [Google Scholar] [CrossRef]
- Sharma, R.; Kumar, S.; Kapoor, S.; Padwad, Y.; Kumar, D. Nuclear magnetic resonance-based metabolomics and cytotoxicity (HT-29 and HCT-116 cell lines) studies insight the potential of less utilized parts of Camellia sinensis (Kangra tea). Food Chem. 2022, 373, 131561. [Google Scholar] [CrossRef]
- Li, C.; Sun, X.; Song, Z.; Sun, J.; Li, Y.; Wang, N.; Zhang, D.; Ye, F.; Dai, J. Chemical constituents from the stems of Dendrobium gratiosissimum and their biological activities. Phytochemistry 2022, 201, 113260. [Google Scholar] [CrossRef] [PubMed]
- Lima, G.S.; Lima, N.M.; Roque, J.V.; de Aguiar, D.V.A.; Oliveira, J.V.A.; Dos Santos, G.F.; Chaves, A.R.; Vaz, B.G. LC-HRMS/MS-Based metabolomics approaches applied to the detection of antifungal compounds and a metabolic dynamic assessment of Orchidaceae. Molecules 2022, 27, 7937. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Lv, Q.; Liu, A.; Wang, J.; Sun, X.; Deng, J.; Chen, Q.; Wu, Q. Comparative metabolomics study of Tartary (Fagopyrum tataricum (L.) Gaertn) and common (Fagopyrum esculentum Moench) buckwheat seeds. Food Chem. 2022, 371, 131125. [Google Scholar] [CrossRef]
- Zaharudin, N.; Staerk, D.; Dragsted, L.O. Inhibition of alpha-glucosidase activity by selected edible seaweeds and fucoxanthin. Food Chem. 2019, 270, 481–486. [Google Scholar] [CrossRef]
- Chen, F.; Deng, Z.-Y.; Zhang, B.; Xiong, Z.-X.; Zheng, S.-L.; Tan, C.-L.; Hu, J.-N. Esterification of ginsenoside rh2 enhanced its cellular uptake and antitumor activity in human Hepg2 cells. J. Agric. Food Chem. 2016, 64, 253–261. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xie, L.; Huang, J.; Xiong, T.; Ma, Y. Secondary Metabolomic Analysis and In Vitro Bioactivity Evaluation of Stems Provide a Comprehensive Comparison between Dendrobium chrysotoxum and Dendrobium thyrsiflorum. Molecules 2023, 28, 6039. https://doi.org/10.3390/molecules28166039
Xie L, Huang J, Xiong T, Ma Y. Secondary Metabolomic Analysis and In Vitro Bioactivity Evaluation of Stems Provide a Comprehensive Comparison between Dendrobium chrysotoxum and Dendrobium thyrsiflorum. Molecules. 2023; 28(16):6039. https://doi.org/10.3390/molecules28166039
Chicago/Turabian StyleXie, Lihang, Jinyong Huang, Tingjian Xiong, and Yao Ma. 2023. "Secondary Metabolomic Analysis and In Vitro Bioactivity Evaluation of Stems Provide a Comprehensive Comparison between Dendrobium chrysotoxum and Dendrobium thyrsiflorum" Molecules 28, no. 16: 6039. https://doi.org/10.3390/molecules28166039
APA StyleXie, L., Huang, J., Xiong, T., & Ma, Y. (2023). Secondary Metabolomic Analysis and In Vitro Bioactivity Evaluation of Stems Provide a Comprehensive Comparison between Dendrobium chrysotoxum and Dendrobium thyrsiflorum. Molecules, 28(16), 6039. https://doi.org/10.3390/molecules28166039