
Garcinia mangostana L. Pericarp Extract and Its Active Compound α-Mangostin as Potential Inhibitors of Immune Checkpoint Programmed Death Ligand-1

 , , ,
, , ,
Abstract
:
1. Introduction
2. Results and Discussion
2.1. Identification and Quantitation of α-Mangostin by HPLC Analysis
2.2. Cancer Cell Line Selection
2.3. Cell Viability
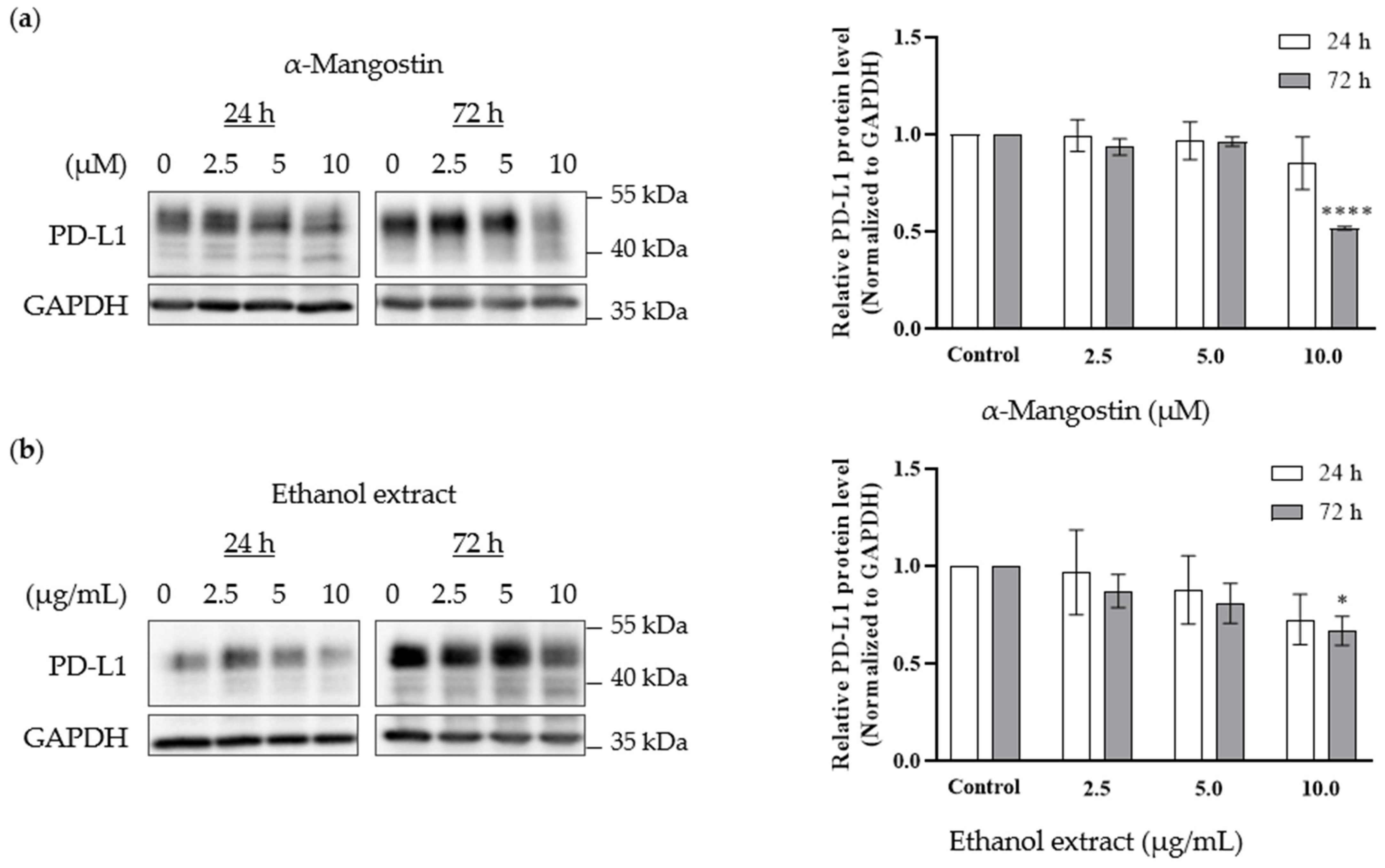
2.4. Western Blotting
2.5. Immunofluorescence Staining
2.6. ADMET Prediction
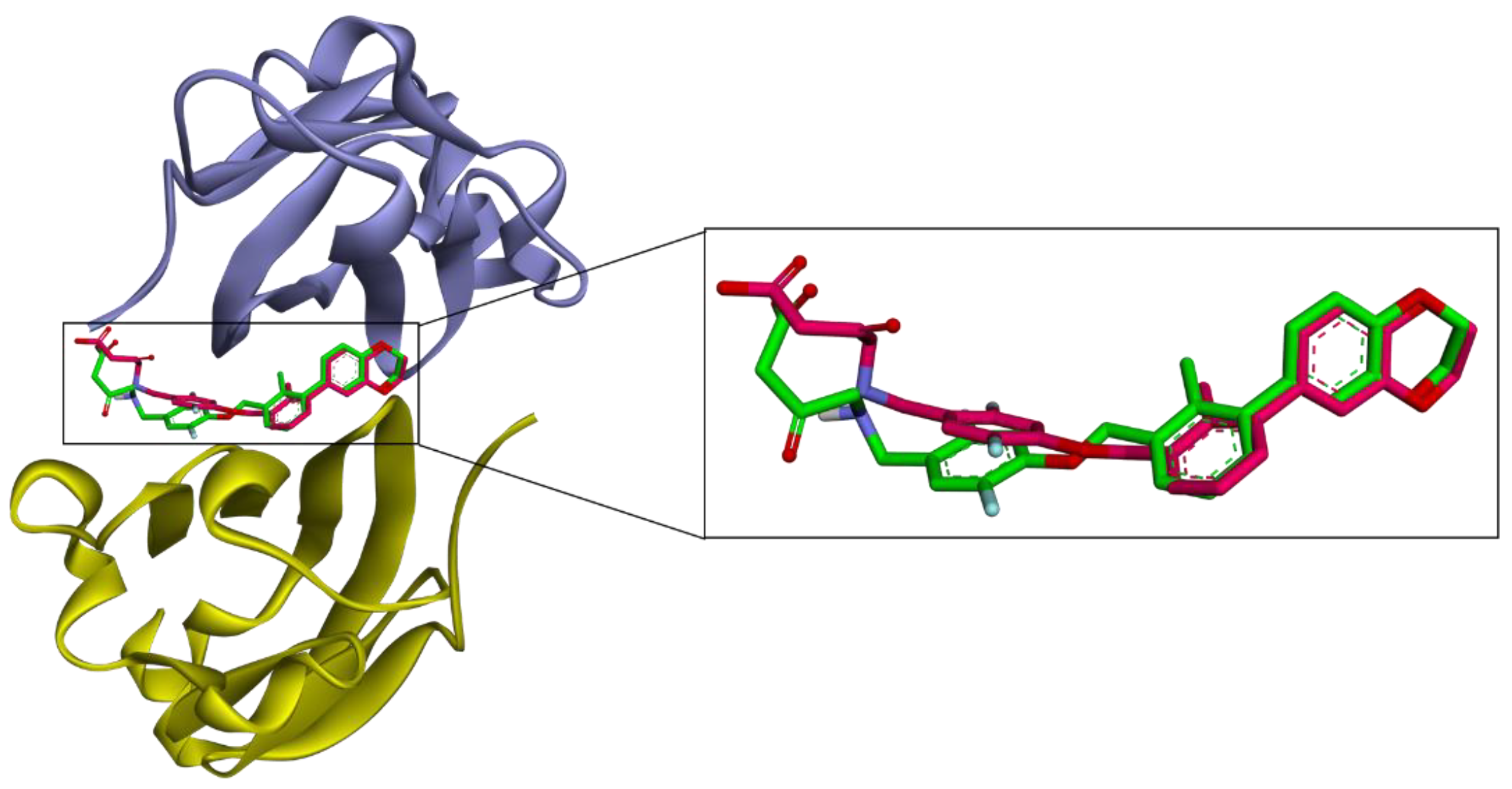
2.7. Molecular Docking
2.8. Molecular Dynamics Simulation
2.9. Binding Free Energy Calculation
3. Materials and Methods
3.1. Plant Materials
3.2. Chemicals and Reagents
3.3. Extraction of Mangosteen Pericarp
3.4. Identification and Quantification of α-Mangostin by HPLC Analysis
3.4.1. Sample Preparation
3.4.2. HPLC Analysis of α-Mangostin in Mangosteen Pericarp Extract
3.5. Cell Culture and Cancer Cell Line Selection
3.6. Cell Treatment
3.7. Cell Viability
3.8. In Vitro Analysis of the Effect of α-Mangostin and Mangosteen Pericarp Extract on PD-L1 Expression
3.8.1. Western Blotting
3.8.2. Immunofluorescence Staining
3.9. ADMET Prediction of α-Mangostin
3.10. In Silico Analysis of the Inhibitory Activity of α-Mangostin on PD-L1 Protein
3.10.1. Molecular Docking
3.10.2. Molecular Dynamics Simulation
3.10.3. Binding Free Energy Calculation
3.11. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Verma, N.; Pandit, S.; Kumar, A.; Yadav, G.; Giri, S.K.; Lahiri, D.; Kari, Z.A.; Prasad, R.; KanuPriya. Recent update on active biological molecules in generating the anticancerous therapeutic potential of Garcinia mangostana. Appl. Biochem. Biotechnol. 2022, 194, 4724–4744. [Google Scholar] [CrossRef] [PubMed]
- Pedraza-Chaverri, J.; Cárdenas-Rodríguez, N.; Orozco-Ibarra, M.; Pérez-Rojas, J.M. Medicinal properties of mangosteen (Garcinia mangostana). Food Chem. Toxicol. 2008, 46, 3227–3239. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, G.A.; Al-Abd, A.M.; El-halawany, A.M.; Abdallah, H.M.; Ibrahim, S.R.M. New xanthones and cytotoxic constituents from Garcinia mangostana fruit hulls against human hepatocellular, breast, and colorectal cancer cell lines. J. Ethnopharmacol. 2017, 198, 302–312. [Google Scholar] [CrossRef] [PubMed]
- Akao, Y.; Nakagawa, Y.; Iinuma, M.; Nozawa, Y. Anti-cancer effects of xanthones from pericarps of mangosteen. Int. J. Mol. Sci. 2008, 9, 355–370. [Google Scholar] [CrossRef]
- Shan, T.; Ma, Q.; Guo, K.; Liu, J.; Li, W.; Wang, F.; Wu, E. Xanthones from mangosteen extracts as natural chemopreventive agents: Potential anticancer drugs. Curr. Mol. Med. 2011, 11, 666–677. [Google Scholar] [CrossRef]
- Krajarng, A.; Nakamura, Y.; Suksamrarn, S.; Watanapokasin, R. α-Mangostin induces apoptosis in human chondrosarcoma cells through downregulation of ERK/JNK and Akt signaling pathway. J. Agric. Food Chem. 2011, 59, 5746–5754. [Google Scholar] [CrossRef]
- Hafeez, B.B.; Mustafa, A.; Fischer, J.W.; Singh, A.; Zhong, W.; Shekhani, M.O.; Meske, L.; Havighurst, T.; Kim, K.; Verma, A.K. α-Mangostin: A dietary antioxidant derived from the pericarp of Garcinia mangostana L. inhibits pancreatic tumor growth in xenograft mouse model. Antioxid. Redox Signal. 2014, 21, 682–699. [Google Scholar] [CrossRef]
- Xia, Y.; Liu, X.; Zou, C.; Feng, S.; Guo, H.; Yang, Y.; Lei, Y.; Zhang, J.; Lu, Y. Garcinone C exerts antitumor activity by modulating the expression of ATR/Stat3/4E-BP1 in nasopharyngeal carcinoma cells. Oncol. Rep. 2018, 39, 1485–1493. [Google Scholar] [CrossRef]
- Yang, X.; Wang, S.; Ouyang, Y.; Tu, Y.; Liu, A.; Tian, Y.; He, M.; Pi, R. Garcinone D, a natural xanthone promotes C17.2 neural stem cell proliferation: Possible involvement of STAT3/Cyclin D1 pathway and Nrf2/HO-1 pathway. Neurosci. Lett. 2016, 626, 6–12. [Google Scholar] [CrossRef]
- Vemu, B.; Nauman, M.C.; Veenstra, J.P.; Johnson, J.J. Structure activity relationship of xanthones for inhibition of cyclin dependent kinase 4 from mangosteen (Garcinia mangostana L.). Int. J. Nutr. 2019, 4, 38–45. [Google Scholar] [CrossRef]
- Aizat, W.M.; Ahmad-Hashim, F.H.; Syed Jaafar, S.N. Valorization of mangosteen, “the queen of fruits,” and new advances in postharvest and in food and engineering applications: A review. J. Adv. Res. 2019, 20, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Lomarat, P.; Moongkarndi, P.; Jaengprajak, J.; Samer, J.; Srisukh, V. Three functional foods from Garcinia mangostana L. using low-α-mangostin aqueous extract of the pericarp: Product development, bioactive compound extractions and analyses, and sensory evaluation. Thai J. Pharm. Sci. 2019, 43, 49–56. [Google Scholar]
- Chitchumroonchokchai, C.; Riedl, K.M.; Suksumrarn, S.; Clinton, S.K.; Kinghorn, A.D.; Failla, M.L. Xanthones in mangosteen juice are absorbed and partially conjugated by healthy adults. J. Nutr. 2012, 142, 675–680. [Google Scholar] [CrossRef] [PubMed]
- Pardoll, D.M. The blockade of immune checkpoints in cancer immunotherapy. Nat. Rev. Cancer 2012, 12, 252–264. [Google Scholar] [CrossRef]
- Ventola, C.L. Cancer immunotherapy, part 1: Current strategies and agents. Pharm. Ther. 2017, 42, 375–383. [Google Scholar]
- Akinleye, A.; Rasool, Z. Immune checkpoint inhibitors of PD-L1 as cancer therapeutics. J. Hematol. Oncol. 2019, 12, 92. [Google Scholar] [CrossRef]
- Han, Y.; Liu, D.; Li, L. PD-1/PD-L1 pathway: Current researches in cancer. Am. J. Cancer Res. 2020, 10, 727–742. [Google Scholar]
- Wu, Y.; Chen, W.; Xu, Z.P.; Gu, W. PD-L1 distribution and perspective for cancer immunotherapy-blockade, knockdown, or inhibition. Front. Immunol. 2019, 10, 2022. [Google Scholar] [CrossRef]
- Daassi, D.; Mahoney, K.M.; Freeman, G.J. The importance of exosomal PDL1 in tumour immune evasion. Nat. Rev. Immunol. 2020, 20, 209–215. [Google Scholar] [CrossRef]
- Guzik, K.; Zak, K.M.; Grudnik, P.; Magiera, K.; Musielak, B.; Törner, R.; Skalniak, L.; Dömling, A.; Dubin, G.; Holak, T.A. Small-molecule inhibitors of the programmed cell death-1/programmed death-ligand 1 (PD-1/PD-L1) interaction via transiently induced protein states and dimerization of PD-L1. J. Med. Chem. 2017, 60, 5857–5867. [Google Scholar] [CrossRef]
- Park, J.J.; Thi, E.P.; Carpio, V.H.; Bi, Y.; Cole, A.G.; Dorsey, B.D.; Fan, K.; Harasym, T.; Iott, C.L.; Kadhim, S.; et al. Checkpoint inhibition through small molecule-induced internalization of programmed death-ligand 1. Nat. Commun. 2021, 12, 1222. [Google Scholar] [CrossRef]
- Wang, T.; Cai, S.; Cheng, Y.; Zhang, W.; Wang, M.; Sun, H.; Guo, B.; Li, Z.; Xiao, Y.; Jiang, S. Discovery of small-molecule inhibitors of the PD-1/PD-L1 axis that promote PD-L1 internalization and degradation. J. Med. Chem. 2022, 65, 3879–3893. [Google Scholar] [CrossRef]
- Wang, T.; Wu, X.; Guo, C.; Zhang, K.; Xu, J.; Li, Z.; Jiang, S. Development of inhibitors of the programmed cell death-1/programmed cell death-ligand 1 signaling pathway. J. Med. Chem. 2019, 62, 1715–1730. [Google Scholar] [CrossRef]
- Validation of Analytical Procedures: Text and Methodology Q2(R1); International Conference on Harmonisation: Geneva, Switzerland, 2005.
- Lee, H.H.; Wang, Y.N.; Xia, W.; Chen, C.H.; Rau, K.M.; Ye, L.; Wei, Y.; Chou, C.K.; Wang, S.C.; Yan, M.; et al. Removal of N-linked glycosylation enhances PD-L1 detection and predicts anti-PD-1/PD-L1 therapeutic efficacy. Cancer Cell 2019, 36, 168–178. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Fang, Y.C.; Li, J. PD-L1 expression levels on tumor cells affect their immunosuppressive activity. Oncol. Lett. 2019, 18, 5399–5407. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.N.; Lee, H.H.; Hsu, J.L.; Yu, D.; Hung, M.C. The impact of PD-L1 N-linked glycosylation on cancer therapy and clinical diagnosis. J. Biomed. Sci. 2020, 27, 77. [Google Scholar] [CrossRef] [PubMed]
- Lipinski, C.A. Lead- and drug-like compounds: The rule-of-five revolution. Drug Discov. Today Technol. 2004, 1, 337–341. [Google Scholar] [CrossRef]
- Gao, Y.; Gesenberg, C.; Zheng, W. Oral formulations for preclinical studies: Principle, design, and development considerations. In Developing Solid Oral Dosage Forms, 2nd ed.; Qiu, Y., Chen, Y., Zhang, G.G.Z., Yu, L., Mantri, R.V., Eds.; Academic Press: Boston, MA, USA, 2017; pp. 455–495. [Google Scholar] [CrossRef]
- Egan, W.J.; Merz, K.M.; Baldwin, J.J. Prediction of drug absorption using multivariate statistics. J. Med. Chem. 2000, 43, 3867–3877. [Google Scholar] [CrossRef]
- Arnott, J.A.; Planey, S.L. The influence of lipophilicity in drug discovery and design. Expert Opin. Drug Discov. 2012, 7, 863–875. [Google Scholar] [CrossRef]
- Bolton, E.E.; Chen, J.; Kim, S.; Han, L.; He, S.; Shi, W.; Simonyan, V.; Sun, Y.; Thiessen, P.A.; Wang, J.; et al. PubChem3D: A new resource for scientists. J. Cheminform. 2011, 3, 32. [Google Scholar] [CrossRef]
- Ramaiya, A.; Li, G.; Petiwala, S.M.; Johnson, J.J. Single dose oral pharmacokinetic profile of α-mangostin in mice. Curr. Drug Targets 2012, 13, 1698–1704. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.H.; Han, S.Y.; Kim, Y.J.; Kim, Y.M.; Chin, Y.W. Absorption, tissue distribution, tissue metabolism and safety of α-mangostin in mangosteen extract using mouse models. Food Chem. Toxicol. 2014, 66, 140–146. [Google Scholar] [CrossRef]
- Hevener, K.E.; Zhao, W.; Ball, D.M.; Babaoglu, K.; Qi, J.; White, S.W.; Lee, R.E. Validation of molecular docking programs for virtual screening against dihydropteroate synthase. J. Chem. Inf. Model. 2009, 49, 444–460. [Google Scholar] [CrossRef] [PubMed]
- Kuang, Z.; Heng, Y.; Huang, S.; Shi, T.; Chen, L.; Xu, L.; Mei, H. Partial least-squares discriminant analysis and ensemble-based flexible docking of PD-1/PD-L1 inhibitors: A pilot study. ACS Omega 2020, 5, 26914–26923. [Google Scholar] [CrossRef] [PubMed]
- Almahmoud, S.; Zhong, H.A. Molecular modeling studies on the binding mode of the PD-1/PD-L1 complex inhibitors. Int. J. Mol. Sci. 2019, 20, 4654. [Google Scholar] [CrossRef]
- Wu, X.; Wang, N.; Liang, J.; Wang, B.; Jin, Y.; Liu, B.; Yang, Y. Is the triggering of PD-L1 dimerization a potential mechanism for food-derived small molecules in cancer immunotherapy? A study by molecular dynamics. Int. J. Mol. Sci. 2023, 24, 1413. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Liang, J.; Liu, B.; Jin, Y. Molecular mechanism of food-derived polyphenols on PD-L1 dimerization: A molecular dynamics simulation study. Int. J. Mol. Sci. 2021, 22, 10924. [Google Scholar] [CrossRef]
- Moongkarndi, P.; Jaisupa, N.; Kosem, N.; Konlata, J.; Samer, J.; Pattanapanyasat, K.; Rodpai, E. Effect of purified α-mangostin from mangosteen pericarp on cytotoxicity, cell cycle arrest and apoptotic gene expression in human cancer cells. World J. Pharm. Sci. 2015, 3, 1473–1484. [Google Scholar]
- Moongkarndi, P.; Jaisupa, N.; Samer, J.; Kosem, N.; Konlata, J.; Rodpai, E.; Pongpan, N. Comparison of the biological activity of two different isolates from mangosteen. J. Pharm. Pharmacol. 2014, 66, 1171–1179. [Google Scholar] [CrossRef]
- Chaivisuthangkura, A.; Malaikaew, Y.; Chaovanalikit, A.; Jaratrungtawee, A.; Panseeta, P.; Ratananukul, P.; Suksamrarn, S. Prenylated xanthone composition of Garcinia mangostana (mangosteen) fruit hull. Chromatographia 2009, 69, 315–318. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Morris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. AutoDock4 and AutoDockTools4: Automated docking with selective receptor flexibility. J. Comput. Chem. 2009, 30, 2785–2791. [Google Scholar] [CrossRef]
- Phillips, J.C.; Hardy, D.J.; Maia, J.D.C.; Stone, J.E.; Ribeiro, J.V.; Bernardi, R.C.; Buch, R.; Fiorin, G.; Hénin, J.; Jiang, W.; et al. Scalable molecular dynamics on CPU and GPU architectures with NAMD. J Chem Phys. 2020, 153, 044130. [Google Scholar] [CrossRef]
- Lee, J.; Cheng, X.; Swails, J.M.; Yeom, M.S.; Eastman, P.K.; Lemkul, J.A.; Wei, S.; Buckner, J.; Jeong, J.C.; Qi, Y.; et al. CHARMM-GUI input Generator for NAMD, GROMACS, AMBER, OpenMM, and CHARMM/OpenMM simulations using the CHARMM36 additive force field. J. Chem. Theory Comput. 2016, 12, 405–413. [Google Scholar] [CrossRef] [PubMed]
- Humphrey, W.; Dalke, A.; Schulten, K. VMD: Visual molecular dynamics. J. Mol. Graph. 1996, 14, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Hou, T. CaFE: A tool for binding affinity prediction using end-point free energy methods. Bioinformatics 2016, 32, 2216–2218. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Results |
|---|---|
| Linearity range (R2 > 0.99) | 0.9993 |
| Accuracy (recovery 80–120%) | 90–101% |
| Precision (%RSD ≤ 2) | ≤0.72 |
| Limit of detection (LOD) | 0.23 µg/mL |
| Limit of quantitation (LOQ) | 0.7 µg/mL |
| Parameters | Results |
|---|---|
| Molecular weight (≤500 Da) | 410.46 Da |
| ALogP (≤5.88) | 5.935 |
| Number of hydrogen acceptors (≤10) | 6 |
| Number of hydrogen donors (≤5) | 3 |
| Rotatable bonds | 5 |
| Polar surface area (131.6 Å2) | 96.22 Å2 |
| Parameters | Results |
|---|---|
| ADMET solubility level a | 1 |
| ADMET absorption level b | 2 |
| CYP2D6 | Non-inhibitor |
| Hepatotoxicity | Toxic |
| ADMET BBB level c | 4 |
| ADMET PPB d | True |
| Parameters | Results |
|---|---|
| Aerobic biodegradability | Non-degradable |
| Ames mutagenicity | Non-mutagen |
| Ocular irritation | Mild irritant |
| Skin irritation | Non-irritant |
| Rat female NTP | Non-carcinogen |
| Rat male NTP | Carcinogen |
| Rat oral LD50 (g/kg) | 0.168 |
| Rodent carcinogenicity | Non-carcinogen |
| Contribution Energy | kcal/mol |
|---|---|
| Electrostatics energy (ΔEelectrostatics) | −6.13 |
| Van der Waals (ΔEvdW) | −51.5655 |
| Polar solvation energy (ΔGPB) | 46.8995 |
| Non-polar solvation (ΔGSA) | −5.4695 |
| Gas phase molecular mechanics (ΔEgas) | −57.6956 |
| Solvation free energy (ΔGsol) | 41.43 |
| Polar binding free energy (ΔGpolar) | 40.7695 |
| Non-polar binding free energy (ΔGnon-polar) | −57.0351 |
| Free binding energy (ΔGbind) | −16.2656 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Naing, S.; Sandech, N.; Maiuthed, A.; Chongruchiroj, S.; Pratuangdejkul, J.; Lomarat, P. Garcinia mangostana L. Pericarp Extract and Its Active Compound α-Mangostin as Potential Inhibitors of Immune Checkpoint Programmed Death Ligand-1. Molecules 2023, 28, 6991. https://doi.org/10.3390/molecules28196991
Naing S, Sandech N, Maiuthed A, Chongruchiroj S, Pratuangdejkul J, Lomarat P. Garcinia mangostana L. Pericarp Extract and Its Active Compound α-Mangostin as Potential Inhibitors of Immune Checkpoint Programmed Death Ligand-1. Molecules. 2023; 28(19):6991. https://doi.org/10.3390/molecules28196991
Chicago/Turabian StyleNaing, Sandar, Nichawadee Sandech, Arnatchai Maiuthed, Sumet Chongruchiroj, Jaturong Pratuangdejkul, and Pattamapan Lomarat. 2023. "Garcinia mangostana L. Pericarp Extract and Its Active Compound α-Mangostin as Potential Inhibitors of Immune Checkpoint Programmed Death Ligand-1" Molecules 28, no. 19: 6991. https://doi.org/10.3390/molecules28196991
APA StyleNaing, S., Sandech, N., Maiuthed, A., Chongruchiroj, S., Pratuangdejkul, J., & Lomarat, P. (2023). Garcinia mangostana L. Pericarp Extract and Its Active Compound α-Mangostin as Potential Inhibitors of Immune Checkpoint Programmed Death Ligand-1. Molecules, 28(19), 6991. https://doi.org/10.3390/molecules28196991