

Effect of Thermal Treatment on the Self-Assembly of Wheat Gluten Polypeptide

Abstract
:1. Introduction
2. Results and Discussion
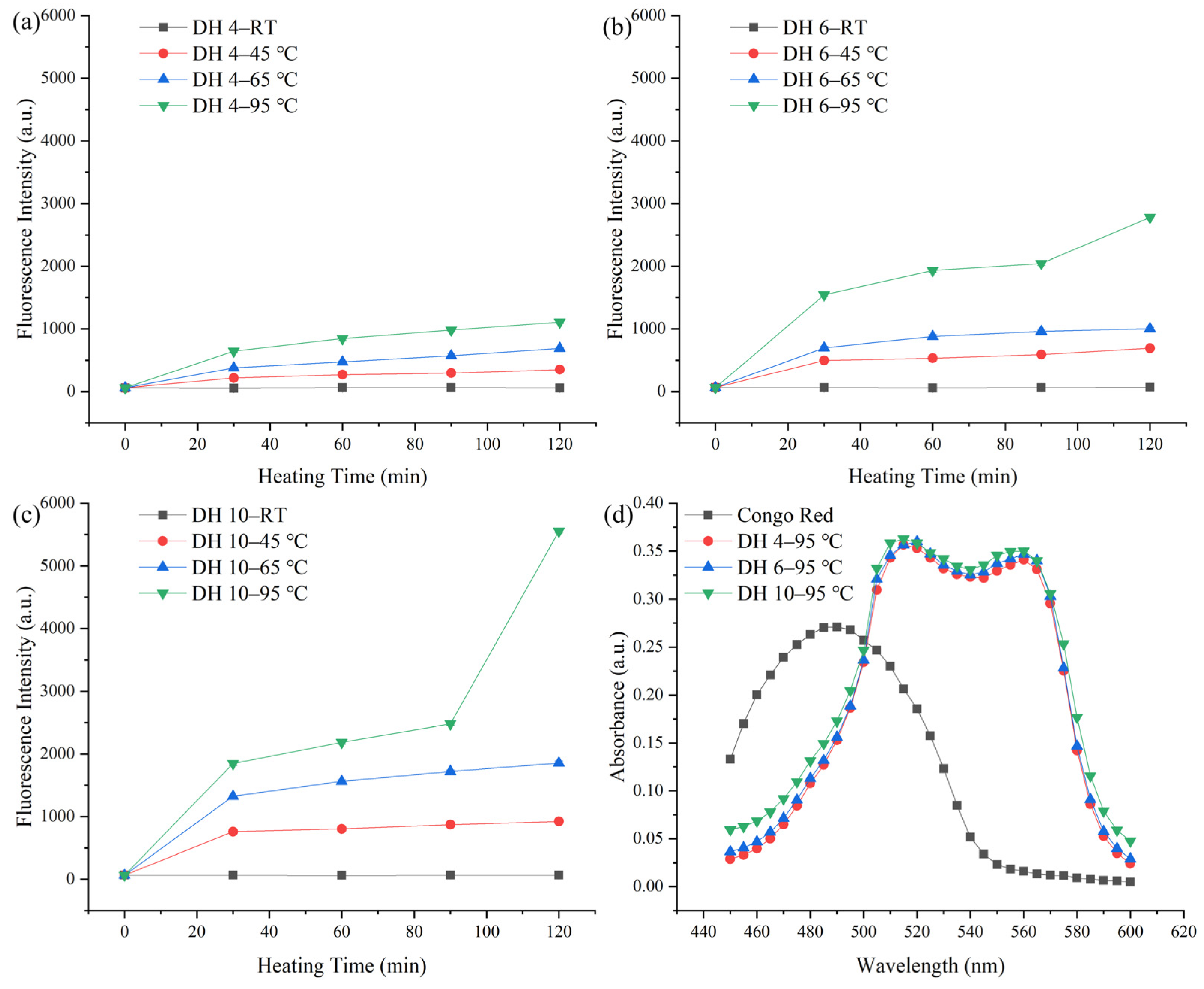
2.1. Effect of the Different Thermal Effects on the Formation of AFs from GPs
2.2. Effect of Different Thermal Effects on the Secondary Structure of GPs
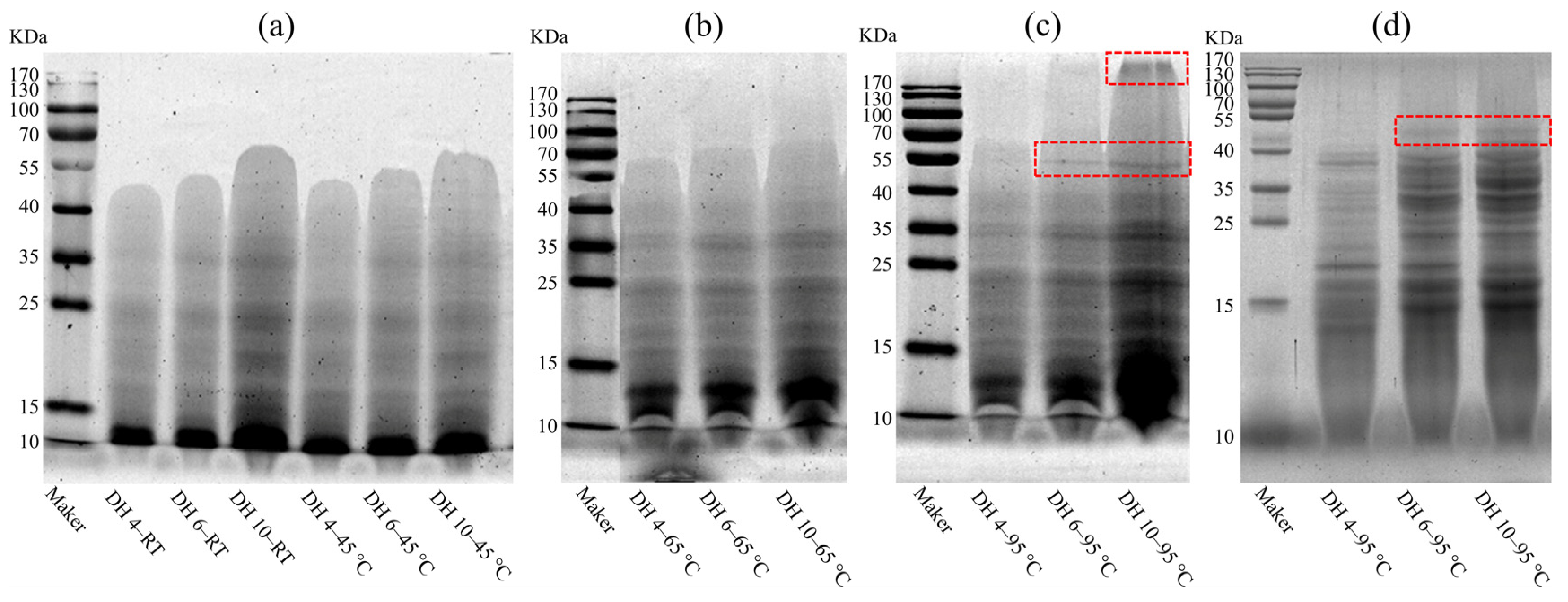
2.3. Effect of the Different Thermal Effects on the Molecular Weight Distribution of GPs
2.4. Molecular Constitution of GPs after Thermal Treatment
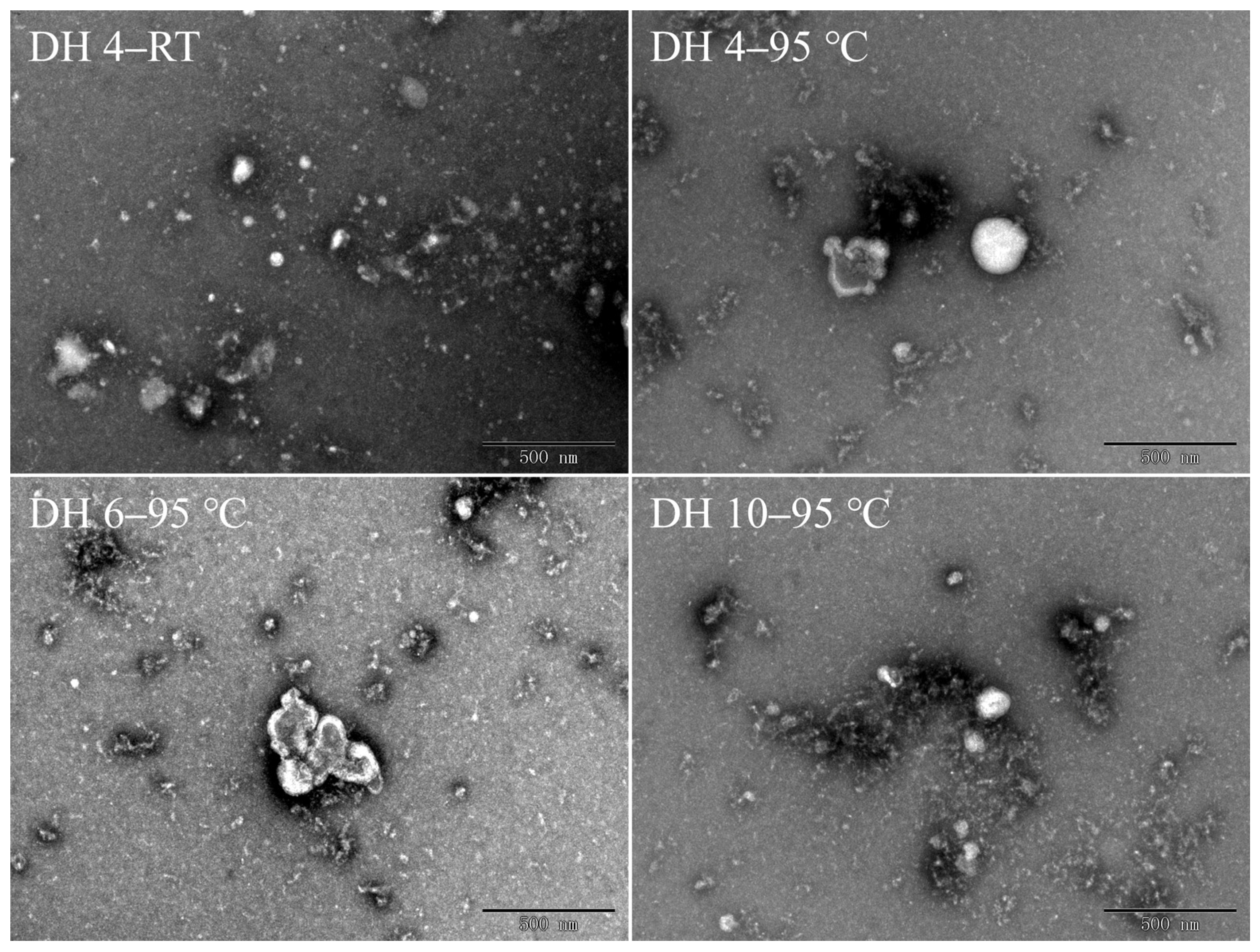
2.5. Morphology of GPs after Thermal Treatment
3. Materials and Methods
3.1. Materials
3.2. Preparation of the Wheat Gluten and Wheat Gluten Polypeptides (GPs)
3.3. Self-Assembly of Wheat Peptides
3.4. Thioflavin T Fluorescence Measurements
3.5. Congo Red Spectral Shift Analysis
3.6. Fourier Transform Infrared (FT-IR) Spectroscopy
3.7. Sodium Dodecyl Sulfate Polyacrylamide Gel Electrophoresis (SDS-PAGE)
3.8. Size-Exclusion High-Performance Liquid Chromatography (SE-HPLC)
3.9. Transmission Electron Microscopy (TEM)
3.10. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Whitesides, G.M.; Grzybowski, B. Self-assembly at all scales. Science 2002, 295, 2418–2421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mezzenga, R.; Schurtenberger, P.; Burbidge, A.; Michel, M. Understanding foods as soft materials. Nat. Mater. 2005, 4, 729–740. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Zhang, Y.; Guo, Z.; Zhang, L. Self-assembly of nanoparticles Static and dynamic. Mater. Today 2019, 25, 112–113. [Google Scholar] [CrossRef]
- Jiang, C.; Xu, G.; Gao, J. Stimuli-responsive macromolecular self-assembly. Sustainability 2022, 14, 11738. [Google Scholar] [CrossRef]
- Shi, B.; Shen, D.; Li, W.; Wang, G. Self-assembly of copolymers containing crystallizable blocks: Strategies and applications. Macromol. Rapid Commun. 2022, 43, 2200071. [Google Scholar] [CrossRef]
- Adamcik, J.; Jung, J.-M.; Flakowski, J.; De Los Rios, P.; Dietler, G.; Mezzenga, R. Understanding amyloid aggregation by statistical analysis of atomic force microscopy images. Nat. Nanotechnol. 2010, 5, 423–428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoppenreijs, L.J.G.; Fitzner, L.; Ruhmlieb, T.; Heyn, T.R.; Schild, K.; van der Goot, A.J.; Boom, R.M.; Steffen-Heins, A.; Schwarz, K.; Keppler, J.K. Engineering amyloid and amyloid-like morphologies of beta-lactoglobulin. Food Hydrocoll. 2022, 124, 107301. [Google Scholar] [CrossRef]
- Jansens, K.J.A.; Brijs, K.; Delcour, J.A.; Scanlon, M.G. Amyloid-like aggregation of ovalbumin: Effect of disulfide reduction and other egg white proteins. Food Hydrocoll. 2016, 61, 914–922. [Google Scholar] [CrossRef]
- Jansens, K.J.A.; Lambrecht, M.A.; Rombouts, I.; Monge-Morera, M.; Brijs, K.; Rousseau, F.; Schymkowitz, J.; Delcour, J.A. Conditions Governing Food Protein Amyloid Fibril Formation-Part I: Egg and Cereal Proteins. Compr. Rev. Food Sci. Food Saf. 2019, 18, 1256–1276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.; Berthelier, V.; Hamilton, J.B.; O’Nuallain, B.; Wetzel, R. Amyloid-like features of polyglutamine aggregates and their assembly kinetics. Biochemistry 2002, 41, 7391–7399. [Google Scholar] [CrossRef]
- Xu, Z.; Shan, G.; Hao, N.; Li, L.; Lan, T.; Dong, Y.; Wen, J.; Tian, R.; Zhang, Y.; Jiang, L.; et al. Structure remodeling of soy protein-derived amyloid fibrils mediated by epigallocatechin-3-gallate. Biomaterials 2022, 283, 121455. [Google Scholar] [CrossRef]
- Cao, Y.; Mezzenga, R. Food protein amyloid fibrils: Origin, structure, formation, characterization, applications and health implications. Adv. Colloid Interface Sci. 2019, 269, 334–356. [Google Scholar] [CrossRef] [PubMed]
- Lagrain, B.; Brijs, K.; Delcour, J.A. Reaction kinetics of gliadin-glutenin cross-linking in model systems and in bread making. J. Agric. Food Chem. 2008, 56, 10660–10666. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Liang, Y.; Chen, Z.; Liu, M.; Qu, Z.; He, B.; Zhang, X.; Wang, J. Effect of curdlan on the aggregation behavior of gluten protein in frozen cooked noodles during cooking. J. Cereal Sci. 2022, 103, 103395. [Google Scholar] [CrossRef]
- Wang, Z.; Ma, S.; Sun, B.; Wang, F.; Huang, J.; Wang, X.; Bao, Q. Effects of thermal properties and behavior of wheat starch and gluten on their interaction: A review. Int. J. Biol. Macromol. 2021, 177, 474–484. [Google Scholar] [CrossRef]
- Wang, P.; Zou, M.; Tian, M.Q.; Gu, Z.X.; Yang, R.Q. The impact of heating on the unfolding and polymerization process of frozen-stored gluten. Food Hydrocoll. 2018, 85, 195–203. [Google Scholar] [CrossRef]
- Wang, P.; Zhao, X.; Yang, R.; Zhou, Y.; Zhou, Q.; Gu, Z.; Jiang, D. Water-Extractable Arabinoxylan-Induced Changes in the Conformation and Polymerization Behavior of Gluten upon Thermal Treatment. J. Agric. Food Chem. 2020, 68, 4005–4016. [Google Scholar] [CrossRef]
- Lagrain, B.; Thewissen, B.G.; Brijs, K.; Delcour, J.A. Mechanism of gliadin-glutenin cross-linking during hydrothermal treatment. Food Chem. 2008, 107, 753–760. [Google Scholar] [CrossRef]
- Stanciuc, N.; Banu, I.; Bolea, C.; Patrascu, L.; Aprodu, I. Structural and antigenic properties of thermally treated gluten proteins. Food Chem. 2018, 267, 43–51. [Google Scholar] [CrossRef]
- Hudson, S.A.; Ecroyd, H.; Kee, T.W.; Carver, J.A. The thioflavin T fluorescence assay for amyloid fibril detection can be biased by the presence of exogenous compounds. FEBS J. 2009, 276, 5960–5972. [Google Scholar] [CrossRef]
- Verbauwhede, A.E.; Lambrecht, M.A.; Jekle, M.; Lucas, I.; Fierens, E.; Shegay, O.; Brijs, K.; Delcour, J.A. Microscopic investigation of the formation of a thermoset wheat gluten network in a model system relevant for bread making. Int. J. Food Sci. Technol. 2020, 55, 891–898. [Google Scholar] [CrossRef]
- Wieser, H. Chemistry of gluten proteins. Food Microbiol. 2007, 24, 115–119. [Google Scholar] [CrossRef]
- Lambrecht, M.A.; Monge-Morera, M.; Godefroidt, T.; Vluymans, N.; Deleu, L.J.; Goos, P.; Schymkowitz, J.; Rousseau, F.; Delcour, J.A. Hydrothermal treatments cause wheat gluten-derived peptides to form amyloid-like fibrils. J. Agric. Food Chem. 2021, 69, 1963–1974. [Google Scholar] [CrossRef] [PubMed]
- Klunk, W.E.; Jacob, R.F.; Mason, R.P. Quantifying amyloid by congo red spectral shift assay. Methods Enzymol. 1999, 309, 285–305. [Google Scholar]
- Ridgley, D.M.; Claunch, E.C.; Barone, J.R. Characterization of large amyloid fibers and tapes with fourier transform infrared (FT-IR) and raman spectroscopy. Appl. Spectrosc. 2013, 67, 1417–1426. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, M.; Jain, N.; Dogra, P.; Samai, S.; Mukhopadhyay, S. Nanoscopic amyloid pores formed via stepwise protein assembly. J. Phys. Chem. Lett. 2013, 4, 480–485. [Google Scholar] [CrossRef]
- Bhattacharya, M.; Dogra, P. Self-assembly of ovalbumin amyloid pores: Effects on membrane permeabilization, dipole potential, and bilayer fluidity. Langmuir 2015, 31, 8911–8922. [Google Scholar] [CrossRef] [PubMed]
- Verbauwhede, A.E.; Lambrecht, M.A.; Fierens, E.; Shegay, O.; Brijs, K.; Delcour, J.A. Impact of aqualysin 1 peptidase from Thermus aquaticus on molecular scale changes in the wheat gluten network during bread baking. Food Chem. 2019, 295, 599–606. [Google Scholar] [CrossRef] [PubMed]
- Han, C.; Ma, M.; Yang, T.; Li, M.; Sun, Q. Heat mediated physicochemical and structural changes of wheat gluten in the presence of salt and alkali. Food Hydrocoll. 2021, 120, 106971. [Google Scholar] [CrossRef]
- Eisenberg, D.S.; Sawaya, M.R. Structural studies of amyloid proteins at the molecular level. Annu. Rev. Biochem. 2017, 61, 8. [Google Scholar] [CrossRef] [Green Version]
- Ji, F.; Xu, J.; Ouyang, Y.; Mu, D.; Li, X.; Luo, S.; Shen, Y.; Zheng, Z. Effects of NaCl concentration and temperature on fibrillation, structure, and functional properties of soy protein isolate fibril dispersions. LWT-Food Sci. Technol. 2021, 149, 111862. [Google Scholar] [CrossRef]
- Wang, X.H.; Liang, Y.; Wang, Q.; Chen, Y.; Liu, H.; Wang, J.S. Low-sodium salt mediated aggregation behavior of gluten in wheat dough. Int. J. Biol. Macromol. 2022, 205, 231–239. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | β-Sheet (Fibrillation)/% | β-Sheet (Native)/% | Random Coil/% | α-Helix/% | β-Turn, Antiparallel β-Sheet/% |
|---|---|---|---|---|---|
| DH 4–RT | 4.98 ± 0.22 a | 7.63 ± 0.36 a | 22.09 ± 0.80 f | 30.98 ± 0.76 d | 35.16 ± 0.88 a |
| 45 °C | 12.19 ± 0.22 b | - | 19.12 ± 0.73 bcde | 29.28 ± 0.58 cd | 39.41 ± 0.07 b |
| 65 °C | 12.94 ± 0.37 bc | - | 18.96 ± 0.82 bcde | 26.76 ± 1.25 b | 41.33 ± 2.44 b |
| 95 °C | 13.61 ± 0.11 cd | - | 18.46 ± 0.06 bcd | 26.42 ± 0.14 b | 41.51 ± 0.19 b |
| DH 6–RT | 5.08 ± 0.53 a | 8.18 ± 0.72 a | 20.08 ± 0.82 de | 30.42 ± 0.48 d | 36.24 ± 0.75 a |
| 45 °C | 13.23 ± 0.00 bcd | - | 18.46 ± 0.10 bcd | 27.18 ± 0.11 bc | 41.12 ± 0.21 b |
| 65 °C | 14.22 ± 0.67 cd | - | 17.67 ± 0.15 bc | 27.40 ± 1.80 bc | 40.70 ± 0.98 b |
| 95 °C | 17.81 ± 0.90 e | - | 17.40 ± 0.30 b | 25.38 ± 0.60 b | 39.41 ± 0.01 b |
| DH 10–RT | 4.60 ± 0.42 a | 8.23 ± 0.23 a | 20.43 ± 0.96 e | 30.14 ± 1.17 d | 35.76 ± 0.83 a |
| 45 °C | 13.42 ± 0.00 bcd | - | 19.28 ± 0.49 de | 27.50 ± 0.14 bc | 39.80 ± 0.63 b |
| 65 °C | 14.55 ± 0.53 d | - | 17.52 ± 0.65 bc | 26.67 ± 0.30 b | 40.26 ± 1.60 b |
| 95 °C | 21.42 ± 0.96 f | - | 15.55 ± 0.52 a | 23.21 ± 0.34 a | 39.81 ± 0.78 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, H.; Wang, J.; Liu, M.; Zhang, X.; Liang, Y.; Wang, J. Effect of Thermal Treatment on the Self-Assembly of Wheat Gluten Polypeptide. Molecules 2023, 28, 834. https://doi.org/10.3390/molecules28020834
Liu H, Wang J, Liu M, Zhang X, Liang Y, Wang J. Effect of Thermal Treatment on the Self-Assembly of Wheat Gluten Polypeptide. Molecules. 2023; 28(2):834. https://doi.org/10.3390/molecules28020834
Chicago/Turabian StyleLiu, Hao, Jingxuan Wang, Mei Liu, Xia Zhang, Ying Liang, and Jinshui Wang. 2023. "Effect of Thermal Treatment on the Self-Assembly of Wheat Gluten Polypeptide" Molecules 28, no. 2: 834. https://doi.org/10.3390/molecules28020834
APA StyleLiu, H., Wang, J., Liu, M., Zhang, X., Liang, Y., & Wang, J. (2023). Effect of Thermal Treatment on the Self-Assembly of Wheat Gluten Polypeptide. Molecules, 28(2), 834. https://doi.org/10.3390/molecules28020834