

Isolation and Characterization of Collagen and Collagen Peptides with Hyaluronidase Inhibition Activity Derived from the Skin of Marlin (Istiophoridae)



Abstract
:1. Introduction
2. Results and Discussions
2.1. Collagen Characterization
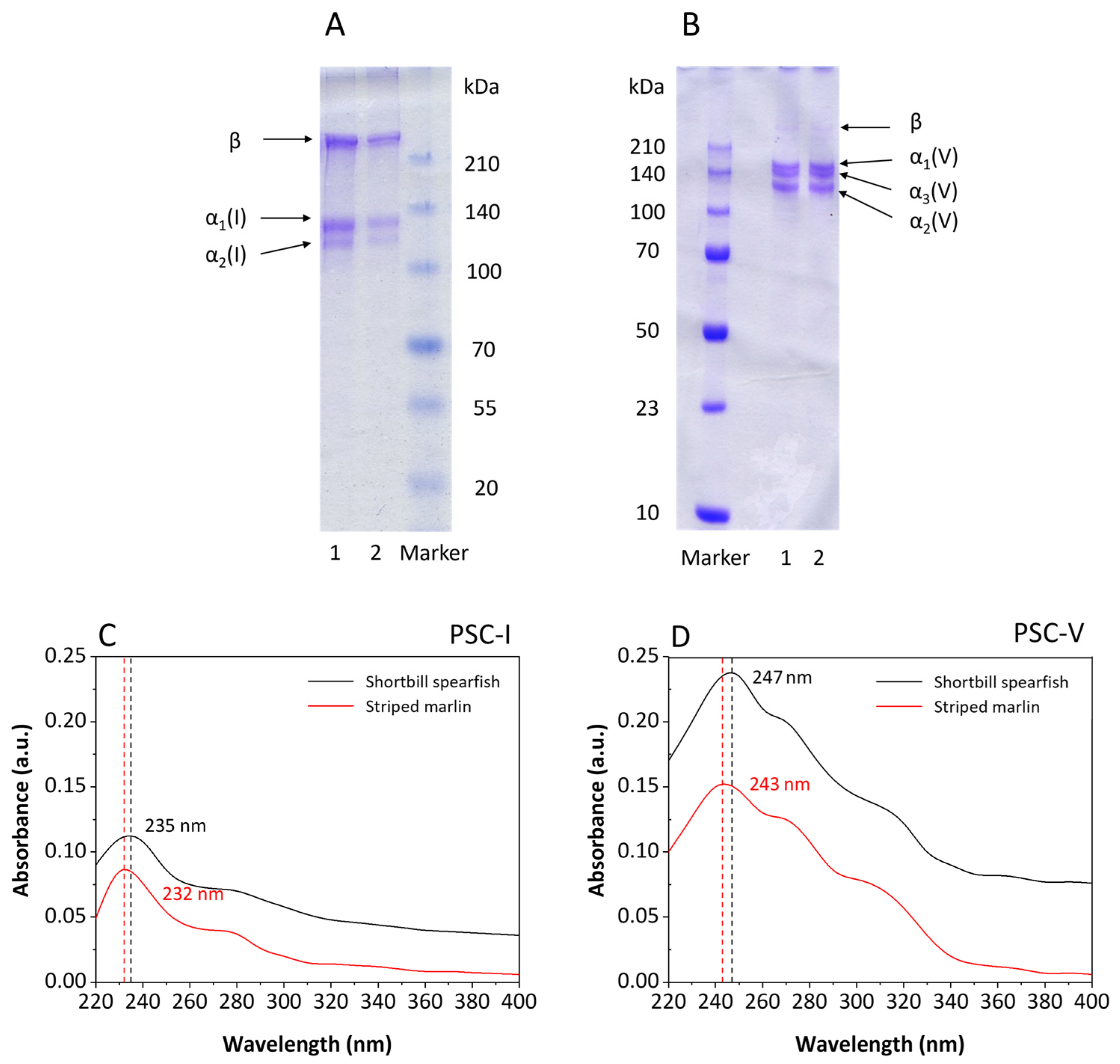
2.1.1. Sodium Dodecyl Sulfate Polyacrylamide Gel Electrophoresis (SDS-PAGE) Pattern of Collagens
2.1.2. Ultraviolet-Visible Spectroscopy (UV-Vis) Scanning
2.2. Collagen Peptide Characterization
2.2.1. Molecular Weight Distribution
2.2.2. Properties of Collagen Peptide
2.3. Peptide Purification
2.3.1. Ultrafiltration of PSC-I and PSC-V Peptides Prepared from Shortbill Spearfish
2.3.2. Purification of Bioactive Peptides by High-Performance Liquid Chromatography (RP-HPLC)
2.4. Stability of Collagen Peptide
2.4.1. Simulated Gastric Fluid (SGF) and Simulated Intestinal Fluid (SIF) Digestion
2.4.2. Effect of Temperature and pH
3. Materials and Methods
3.1. Materials
3.2. Extraction of Collagen from Marlin Skin
3.2.1. Pretreatment
3.2.2. Extraction of Type I and V Collagens
3.3. Preparation of Collagen Peptide
3.3.1. Collagen Peptide Production
3.3.2. Peptide Purification
3.4. Hyp Content
3.5. SDS-Polyacrylamide Gel Electrophoresis (SDS-PAGE)
3.6. Ultraviolet-Visible Spectroscopy (UV-Vis) Scanning
3.7. HAase-I Assay
3.8. DH Determination
3.9. Stability of Collagen Peptide
3.9.1. SGF and SIF Digestion
3.9.2. pH Condition and Temperature Influence
3.10. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Henchion, M.; McCarthy, M.; O’Callaghan, J. Transforming beef by-products into valuable ingredients: Which spell/recipe to use? Front. Nutr. 2016, 3, 53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez-Alvarez, O.; Chamorro, S.; Brenes, A. Protein hydrolysates from animal processing by-products as a source of bioactive molecules with interest in animal feeding: A review. Food Res. Int. 2015, 73, 204–212. [Google Scholar] [CrossRef] [Green Version]
- Olsen, R.L.; Toppe, J.; Karunasagar, I. Challenges and realistic opportunities in the use of by-products from processing of fish and shellfish. Trends Food Sci. Technol. 2014, 36, 144–151. [Google Scholar] [CrossRef]
- Silva, T.H.; Moreira-Silva, J.; Marques, A.L.P.; Domingues, A.; Bayon, Y.; Reis, R.L. Marine origin collagens and its potential applications. Mar. Drugs 2014, 12, 5881–5901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gómez-Guillén, M.C.; Turnay, J.; Fernandez-Diaz, M.D.; Ulmo, N.; Lizarbe, M.A.; Montero, P. Structural and physical properties of gelatin extracted from different marine species: A comparative study. Food Hydrocoll. 2002, 16, 25–34. [Google Scholar] [CrossRef] [Green Version]
- Karayannakidis, P.D.; Chatziantoniou, S.E.; Zotos, A. Effects of selected process parameters on physical and sensorial properties of yellowfin tuna (Thunnus albacares) skin gelatin. J. Food Process Eng. 2014, 37, 461–473. [Google Scholar] [CrossRef]
- Ruszczak, Z. Effect of collagen matrices on dermal wound healing. Adv. Drug Deliv. Rev. 2003, 55, 1595–1611. [Google Scholar] [CrossRef]
- Clare, D.; Swaisgood, H. Bioactive milk peptides: A prospectus. J. Dairy Sci. 2000, 83, 1187–1195. [Google Scholar] [CrossRef]
- Udenigwe, C.C. Bioinformatics approaches, prospects and challenges of food bioactive peptide research. Trends Food Sci. Technol. 2014, 36, 137–143. [Google Scholar] [CrossRef]
- Gómez-Guillén, M.; Giménez, B.; López-caballero, M.E.; Montero, M.P. Functional and bioactive properties of collagen and gelatin from alternative sources: A review. Food hydrocoll. 2011, 25, 1813–1827. [Google Scholar] [CrossRef]
- Xu, L.; Dong, W.; Zhao, J.; Xu, Y. Effect of marine collagen peptides on physiological and neurobehavioral development of male rats with perinatal asphyxia. Mar. Drugs 2015, 13, 3653–3671. [Google Scholar] [CrossRef] [Green Version]
- Alemán, A.; Gómez-Guillén, M.; Montero, P. Identification of ace-inhibitory peptides from squid skin collagen after in vitro gastrointestinal digestion. Food Res. Int. 2013, 54, 790–795. [Google Scholar] [CrossRef]
- Kim, S.K.; Kim, Y.T.; Byun, H.G.; Nam, K.S.; Joo, D.S.; Shahidi, F. Isolation and characterization of antioxidative peptides from gelatin hydrolysate of Alaska pollack skin. J. Agric. Food Chem. 2001, 49, 1984–1989. [Google Scholar] [CrossRef]
- Alemán, A.; Martínez-Alvarez, O. Marine collagen as a source of bioactive molecules: A review. Nat. Prod. J. 2013, 3, 105–114. [Google Scholar] [CrossRef] [Green Version]
- Pati, F.; Adhikari, B.; Dhara, S. Isolation and characterization of fish scale collagen of higher thermal stability. Bioresour. Technol. 2010, 101, 3737–3742. [Google Scholar] [CrossRef]
- Safandowska, M.; Pietrucha, K. Effect of fish collagen modification on its thermal and rheological properties. Int. J. Biol. Macromol. 2013, 53, 32–37. [Google Scholar] [CrossRef]
- Burgeson, R.E.; El Adli, F.A.; Kaitila, I.I.; Hollister, D.W. Fetal membrane collagens: Identification of two new collagen alpha chains. Proc. Natl. Acad. Sci. USA 1976, 73, 2579–2583. [Google Scholar] [CrossRef] [Green Version]
- Chung, E.; Rhodes, R.K.; Miller, E.J. Isolation of three collagenous components of probable basement membrane origin from several tissues. Biochem. Biophys. Res. Commun. 1976, 71, 1167–1174. [Google Scholar] [CrossRef]
- Mayne, R. Structure and Function of Collagen Types; Mayne, R., Burgeson, R.E., Eds.; Academic Press: Cambridge, MA, USA, 2012. [Google Scholar]
- Adachi, E.; Hayashi, T. In vitro formation of hybrid fibrils of type V collagen and type I collagen limited growth of type I collagen into thick fibrils by type V collagen. Connect. Tissue Res. 1986, 14, 257–266. [Google Scholar] [CrossRef]
- Luparello, C.; Schillaci, R.; Pucci-Minafra, I.; Minafra, S. Adhesion, growth and cytoskeletal characteristics of 8701-BC breast carcinoma cells cultured in the presence of type V collagen. Eur. J. Cancer Clin. Oncol. 1990, 26, 231–240. [Google Scholar] [CrossRef]
- Mizuta, S.; Fujisawa, S.; Nishimoto, M.; Yoshinaka, R. Biochemical and immunochemical detection of types I and V collagens in tiger puffer Takifugu rubripes. Food Chem. 2005, 89, 373–377. [Google Scholar] [CrossRef]
- Weigel, P.H.; Hascall, V.C.; Tammi, M. Hyaluronan synthases. J. Biol. Chem. 1997, 272, 13997–14000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noble, P.W.; Lake, F.R.; Henson, P.M.; Henson, P.M. Hyaluronate activation of CD44 induces insulin-like growth factor-1 expression by a tumor necrosis factor-alpha-dependent mechanism in murine macrophages. J. Clin. Investig. 1993, 91, 2368–2377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arunkumar, K.; Raj, R.; Raja, R.; Carvalho, I.S. Brown seaweeds as a source of anti-hyaluronidase compounds. S. Afr. J. Bot. 2021, 139, 470–477. [Google Scholar] [CrossRef]
- Aryani, A.; Suprayitno, E.; Sasmito, B.B.; Hardoko, H. Characterization and identification of charcoal of inedible Kerandang fish (Channa pleurophthalmus Blkr) body parts and potential antiallergenic properties. Vet. World 2020, 13, 1480–1486. [Google Scholar] [CrossRef]
- Kimura, S. Wide distribution of the skin type I collagen alpha 3 chain in bony fish. Comp. Biochem. Physiol. B 1992, 102, 255–260. [Google Scholar] [CrossRef]
- Kimura, S.; Ohno, Y. Fish type I collagen: Tissue-specific existence of two molecular forms, (α1)2α2 and α1α2α3, in Alaska pollack. Comp. Biochem. Physiol. B 1987, 88, 409–413. [Google Scholar] [CrossRef]
- Duan, R.; Zhang, J.; Du, X.; Yao, X.; Konno, K. Properties of collagen from skin, scale and bone of carp (Cyprinus carpio). Food Chem. 2009, 112, 702–706. [Google Scholar] [CrossRef]
- Sun, L.; Hou, H.; Li, B.; Zhang, Y. Characterization of acid-and pepsin-soluble collagen extracted from the skin of Nile tilapia (Oreochromis niloticus). Int. J. Biol. Macromol. 2017, 99, 8–14. [Google Scholar] [CrossRef]
- Wang, L.; Liang, Q.; Wang, Z.; Xu, J.; Liu, Y.; Ma, H. Preparation and characterisation of type I and V collagens from the skin of Amur sturgeon (Acipenser schrenckii). Food Chem. 2014, 148, 410–414. [Google Scholar] [CrossRef]
- Kittiphattanabawon, P.; Benjakul, S.; Visessanguan, W.; Nagai, T.; Tanaka, M. Characterisation of acid-soluble collagen from skin and bone of bigeye snapper (Priacanthus tayenus). Food Chem. 2005, 89, 363–372. [Google Scholar] [CrossRef]
- Li, H.; Liu, B.L.; Gao, L.Z.; Chen, H.L. Studies on bullfrog skin collagen. Food Chem. 2004, 84, 65–69. [Google Scholar] [CrossRef]
- Zamorano-Apodaca, J.C.; García-Sifuentes, C.O.; Carvajal-Millán, E.; Vallejo-Galland, B.; Scheuren-Acevedo, S.M.; Lugo-Sánchez, M.E. Biological and functional properties of peptide fractions obtained from collagen hydrolysate derived from mixed by-products of different fish species. Food Chem. 2020, 331, 127350. [Google Scholar] [CrossRef]
- Jeevithan, E.; Bao, B.; Zhang, J.; Hong, S.; Wu, W. Purification, characterization and antioxidant properties of low molecular weight collagenous polypeptide (37 kDa) prepared from whale shark cartilage (Rhincodon typus). J. Food Sci. Technol. 2015, 52, 6312–6322. [Google Scholar] [CrossRef] [Green Version]
- Han, Q.Y.; Ishizaki, S.; Bao, B. Optimization for preparation of collagen peptide with hyaluronidase inhibition activity by response surface method. Food Mach. 2021, 11, 37. [Google Scholar]
- Raghavan, S.; Kristinsson, H.G. Antioxidative efficacy of alkali-treated tilapia protein hydrolysates: A comparative study of five enzymes. J. Agric. Food Chem. 2008, 56, 1434–1441. [Google Scholar] [CrossRef]
- Bamdad, F.; Wu, J.; Chen, L. Effects of enzymatic hydrolysis on molecular structure and antioxidant activity of barley hordein. J. Cereal Sci. 2011, 54, 20–28. [Google Scholar] [CrossRef]
- Cheung, I.W.; Cheung, L.K.; Tan, N.Y.; Li-Chan, E.C. The role of molecular size in antioxidant activity of peptide fractions from Pacific hake (Merluccius productus) hydrolysates. Food Chem. 2012, 134, 1297–1306. [Google Scholar] [CrossRef]
- Liu, Z.Y.; Chen, D.; Su, Y.C.; Zeng, M.Y. Optimization of hydrolysis conditions for the production of the angiotensin-I converting enzyme inhibitory peptides from sea cucumber collagen hydrolysates. J. Aquat. Food Prod. Technol. 2011, 20, 222–232. [Google Scholar] [CrossRef]
- Tavianatou, A.G.; Caon, I.; Franchi, M.; Piperigkou, Z.; Galesso, D.; Karamanos, N.K. Hyaluronan: Molecular size-dependent signaling and biological functions in inflammation and cancer. FEBS J. 2019, 286, 2883–2908. [Google Scholar] [CrossRef] [Green Version]
- Moran Jr, E.T. Gastric digestion of protein through pancreozyme action optimizes intestinal forms for absorption, mucin formation and villus integrity. Anim. Feed Sci. Technol. 2016, 221, 284–303. [Google Scholar] [CrossRef]
- Wu, J.; Ding, X. Characterization of inhibition and stability of soy-protein-derived angiotensin I-converting enzyme inhibitory peptides. Food Res. Int. 2002, 35, 367–375. [Google Scholar] [CrossRef]
- Wu, W.; He, L.; Liang, Y.; Yue, L.; Peng, W.; Jin, G.; Ma, M. Preparation process optimization of pig bone collagen peptide-calcium chelate using response surface methodology and its structural characterization and stability analysis. Food Chem. 2019, 284, 80–89. [Google Scholar] [CrossRef] [PubMed]
- Pu, S.Z.; Zhang, W.H.; Shi, B. Effect of pH on structure and stability of collagen-like peptide: Insight from molecular dynamics simulation. J. Theor. Comput. Chem. 2011, 10, 245–259. [Google Scholar] [CrossRef]
- Bergman, I.; Loxley, R. Two improved and simplified methods for the spectrophotometric determination of hydroxyproline. Anal. Chem. 1963, 35, 1961–1965. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Kittiphattanabawon, P.; Benjakul, S.; Visessanguan, W.; Kishimura, H.; Shahidi, F. Isolation and characterisation of collagen from the skin of brownbanded bamboo shark (Chiloscyllium punctatum). Food Chem. 2010, 119, 1519–1526. [Google Scholar] [CrossRef]
- Meyer, K. The biological significance of hyaluronic acid and hyaluronidase. Physiol. Rev. 1947, 27, 335–359. [Google Scholar] [CrossRef] [Green Version]
- Adler-Nissen, J. Determination of the degree of hydrolysis of food protein hydrolysates by trinitrobenzenesulfonic acid. J. Agric. Food Chem. 1979, 27, 1256–1262. [Google Scholar] [CrossRef]
- United States Pharmacopoeial Convention (USPC). United States Pharmacopoeia-National Formulary (USP-NF); US Pharmacopoeial Convention, Inc.: Rockville, MD, USA, 2019; pp. 639–641+814+820. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Hyp Content (mg/g) | HAase-I Rate (%) | DH (%) | Yield 1 (%) | |
|---|---|---|---|---|---|
| PSC-I peptide | Shortbill spearfish | 132.37 ± 0.43 a | 39.83 ± 0.06 a | 24.81 ± 0.23 a | 12.04 ± 0.65 a |
| Striped marlin | 129.51 ± 0.20 a | 37.25 ± 0.84 b | 22.11 ± 0.28 b | 12.50 ± 0.66 a | |
| PSC-V peptide | Shortbill spearfish | 109.87 ± 0.33 b | 24.13 ± 0.32 c | 9.82 ± 0.74 c | 3.73 ± 0.08 b |
| Striped marlin | 116.56 ± 0.19 ab | 23.84 ± 0.17 c | 9.37 ± 0.41 d | 4.02 ± 0.91 b |
| Sample | Size | Concentrations (mg/mL) | |||
|---|---|---|---|---|---|
| 1 | 2 | 4 | 8 | ||
| PSC-I peptide | Crude | 5.96 ± 0.33 a | 23.91 ± 0.39 a | 34.78 ± 0.05 c | 39.83 ± 0.28 d |
| >30 kDa | 5.13 ± 0.41 a | 21.54 ± 0.17 b | 24.84 ± 0.05 c | 37.75 ± 0.25 d | |
| <30 kDa | 8.58 ± 0.08 a | 29.93 ± 0.75 b | 36.20 ± 0.86 c | 56.67 ± 0.10 d | |
| PSC-V peptide | Crude | 20.32 ± 0.26 a | 20.18 ± 0.81 a | 18.05 ± 0.08 b | 16.13 ± 0.33 c |
| >30 kDa | 18.53 ± 0.57 a | 18.17 ± 1.11 a | 17.16 ± 0.33 a | 14.38 ± 0.14 b | |
| <30 kDa | 24.02 ± 0.35 a | 24.00 ± 0.43 a | 22.67 ± 0.21 b | 18.89 ± 0.14 c | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, Q.-Y.; Koyama, T.; Watabe, S.; Nagashima, Y.; Ishizaki, S. Isolation and Characterization of Collagen and Collagen Peptides with Hyaluronidase Inhibition Activity Derived from the Skin of Marlin (Istiophoridae). Molecules 2023, 28, 889. https://doi.org/10.3390/molecules28020889
Han Q-Y, Koyama T, Watabe S, Nagashima Y, Ishizaki S. Isolation and Characterization of Collagen and Collagen Peptides with Hyaluronidase Inhibition Activity Derived from the Skin of Marlin (Istiophoridae). Molecules. 2023; 28(2):889. https://doi.org/10.3390/molecules28020889
Chicago/Turabian StyleHan, Qiu-Yu, Tomoyuki Koyama, Shugo Watabe, Yuji Nagashima, and Shoichiro Ishizaki. 2023. "Isolation and Characterization of Collagen and Collagen Peptides with Hyaluronidase Inhibition Activity Derived from the Skin of Marlin (Istiophoridae)" Molecules 28, no. 2: 889. https://doi.org/10.3390/molecules28020889
APA StyleHan, Q. -Y., Koyama, T., Watabe, S., Nagashima, Y., & Ishizaki, S. (2023). Isolation and Characterization of Collagen and Collagen Peptides with Hyaluronidase Inhibition Activity Derived from the Skin of Marlin (Istiophoridae). Molecules, 28(2), 889. https://doi.org/10.3390/molecules28020889