
Selected N-Terpenyl Organoselenium Compounds Possess Antimycotic Activity In Vitro and in a Mouse Model of Vulvovaginal Candidiasis


 , , ,
, , ,  and
and
Abstract
:1. Introduction
- (i)
- in vitro growth inhibition and medium acidification experiments, and
- (ii)
- an in vivo experiment using a mouse model of VVC.

2. Results
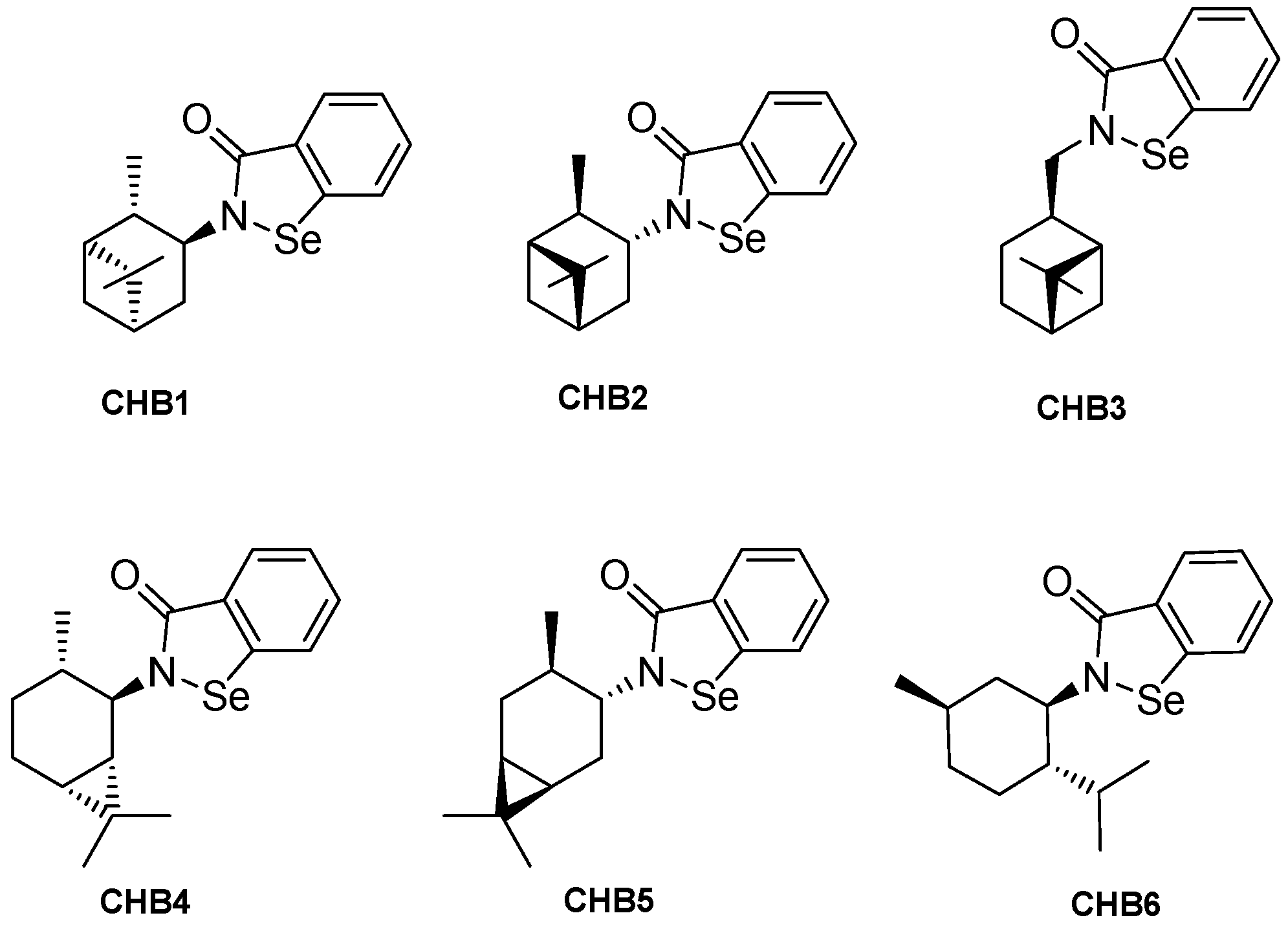
2.1. Chemical Properties and MIC Determination
2.2. Comparing Half Maximal Inhibitory Concentrations (IC50 Values) in KB-3-1 Cells and C. albicans
2.3. Effect on Medium Acidification in C. albicans S1
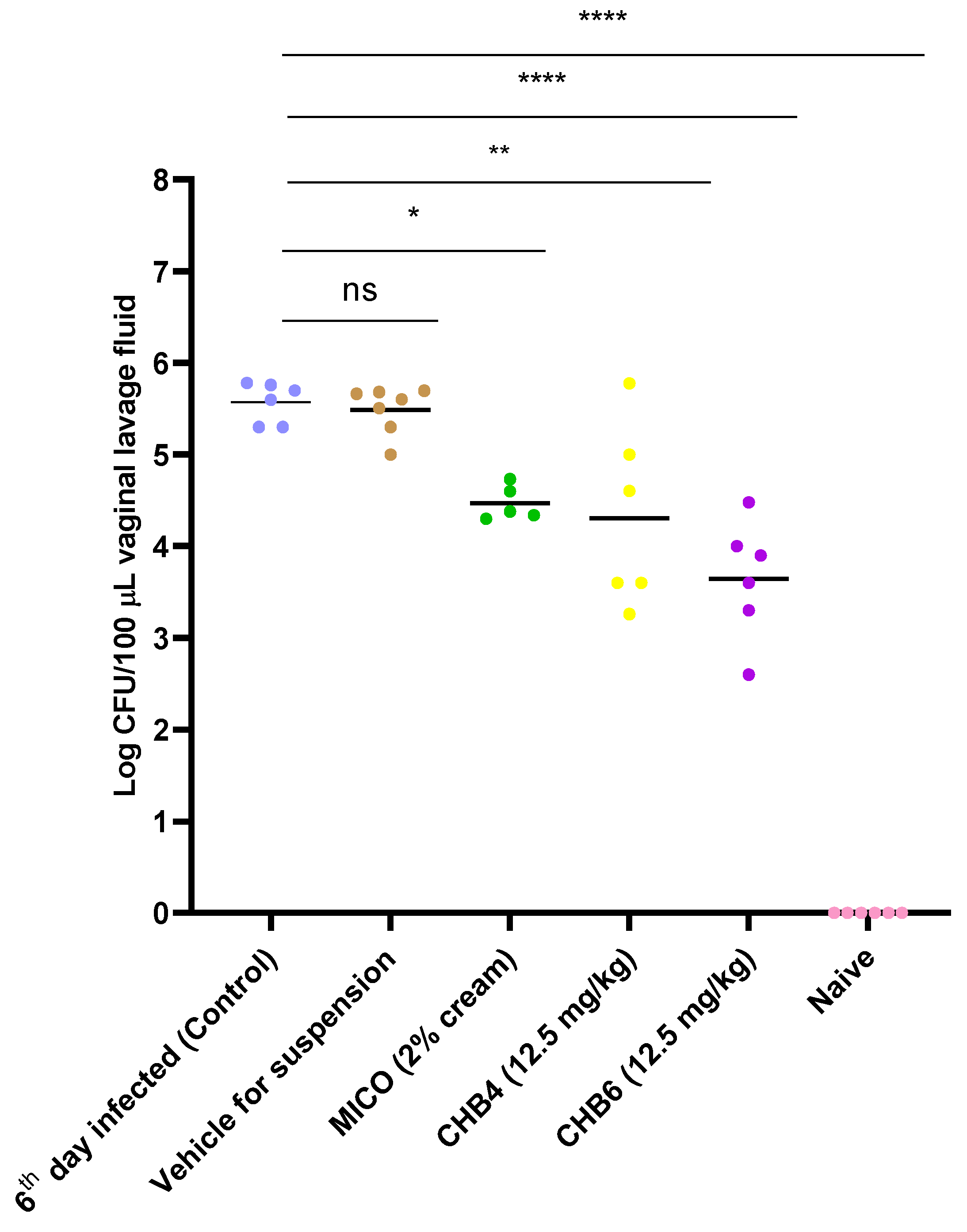
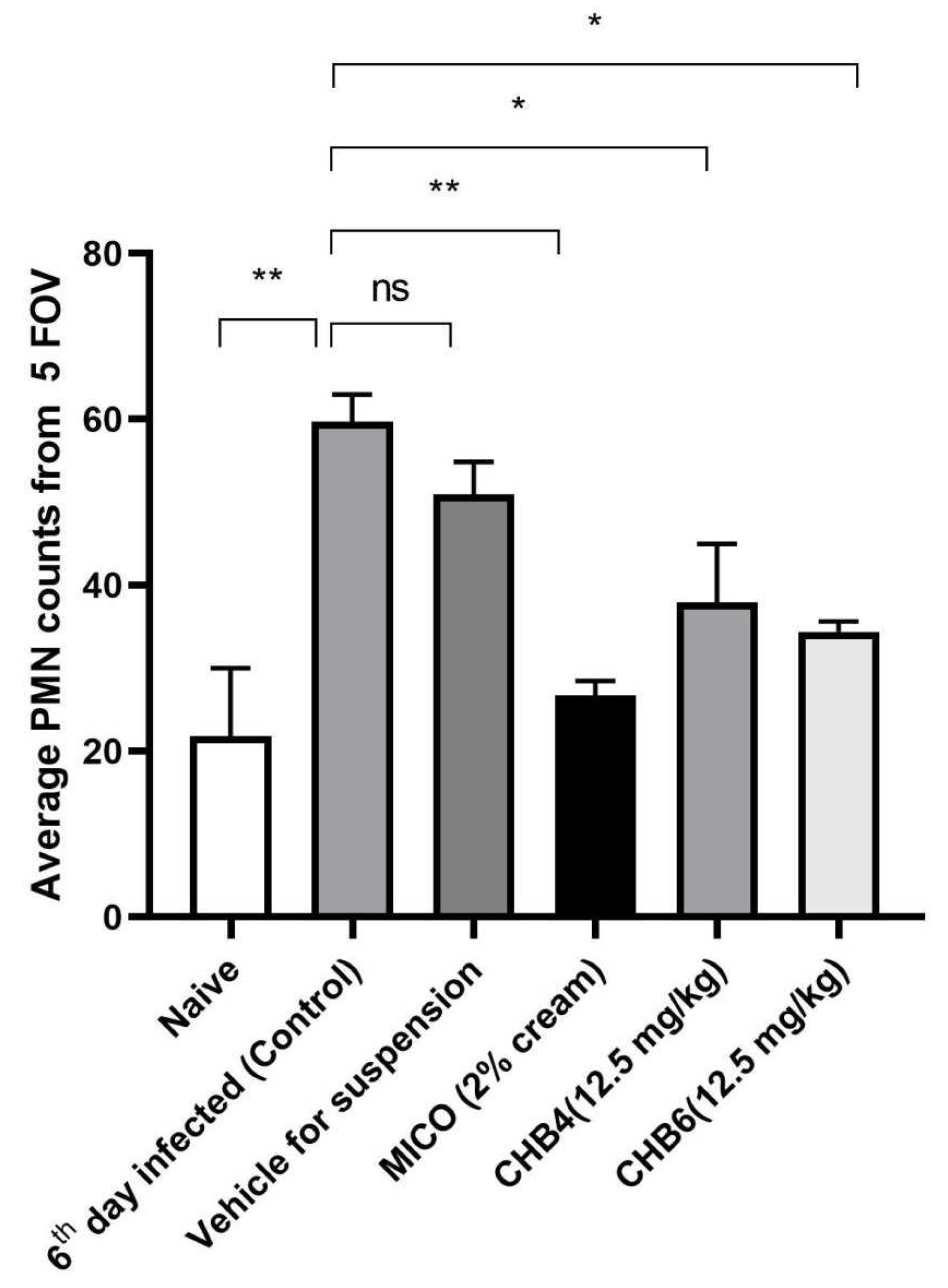
2.4. Efficacy of Treatments in the Mouse Model of VVC
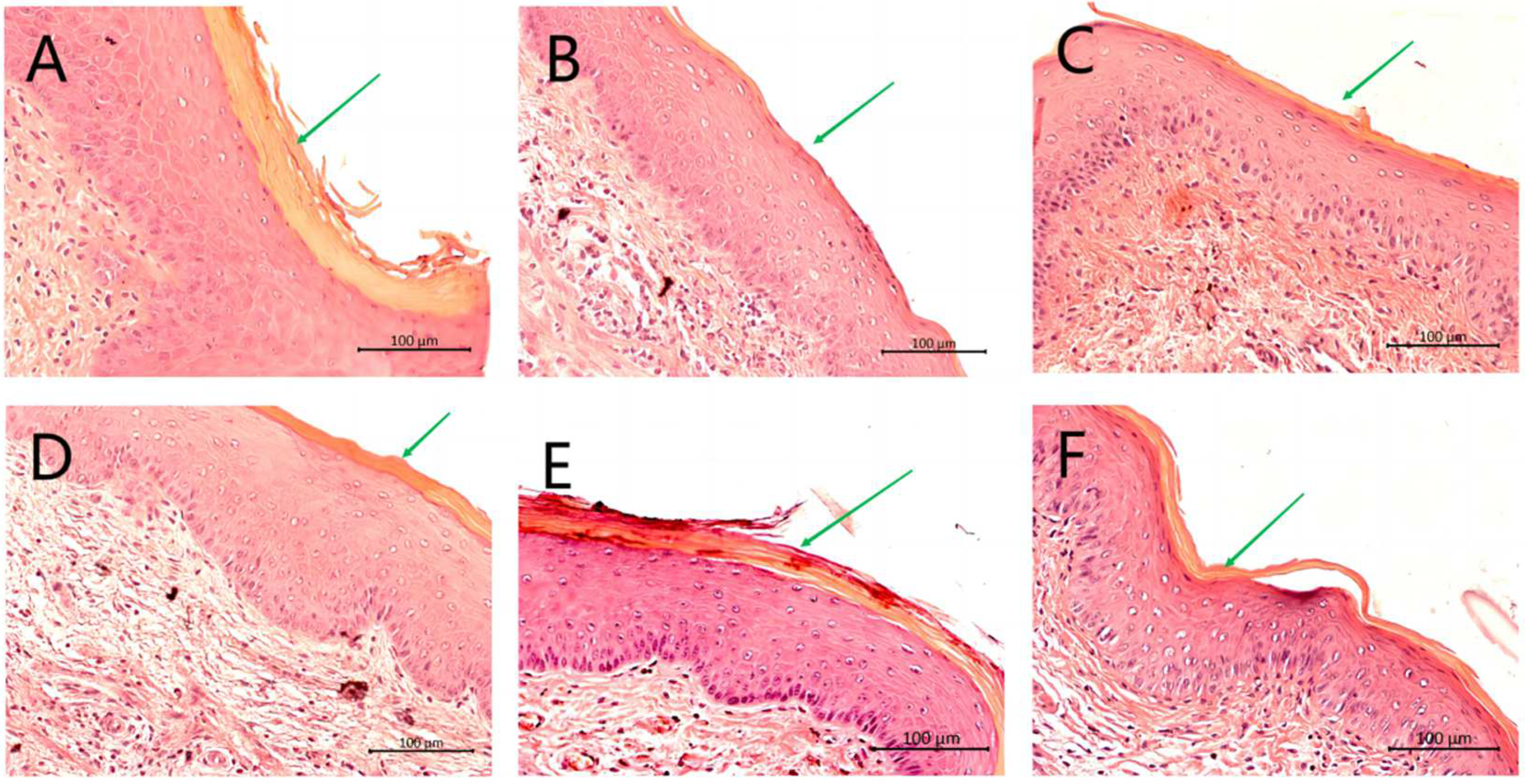
2.5. Histological Analysis of the Vaginal Tissue
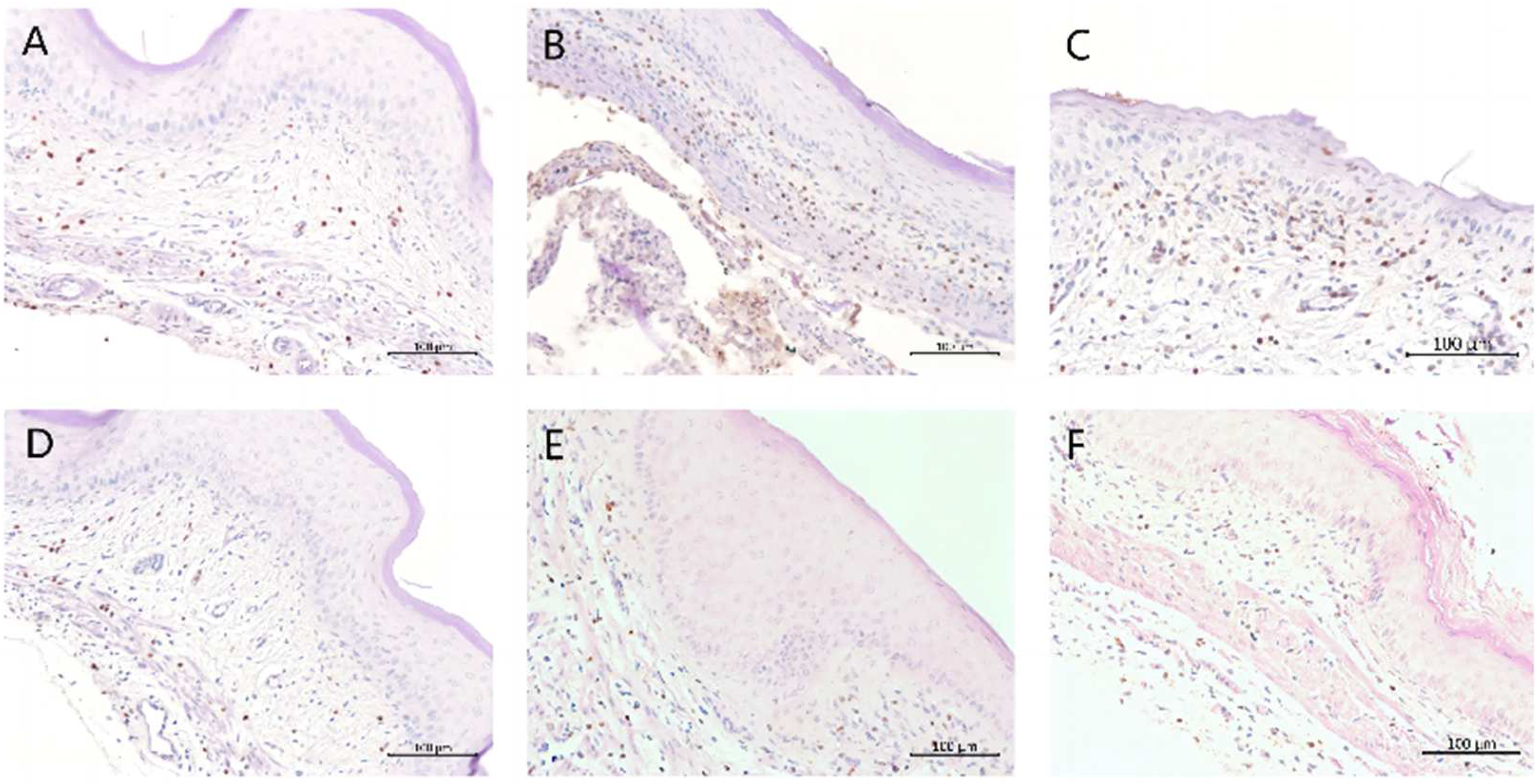
2.6. Immunohistochemical Analysis
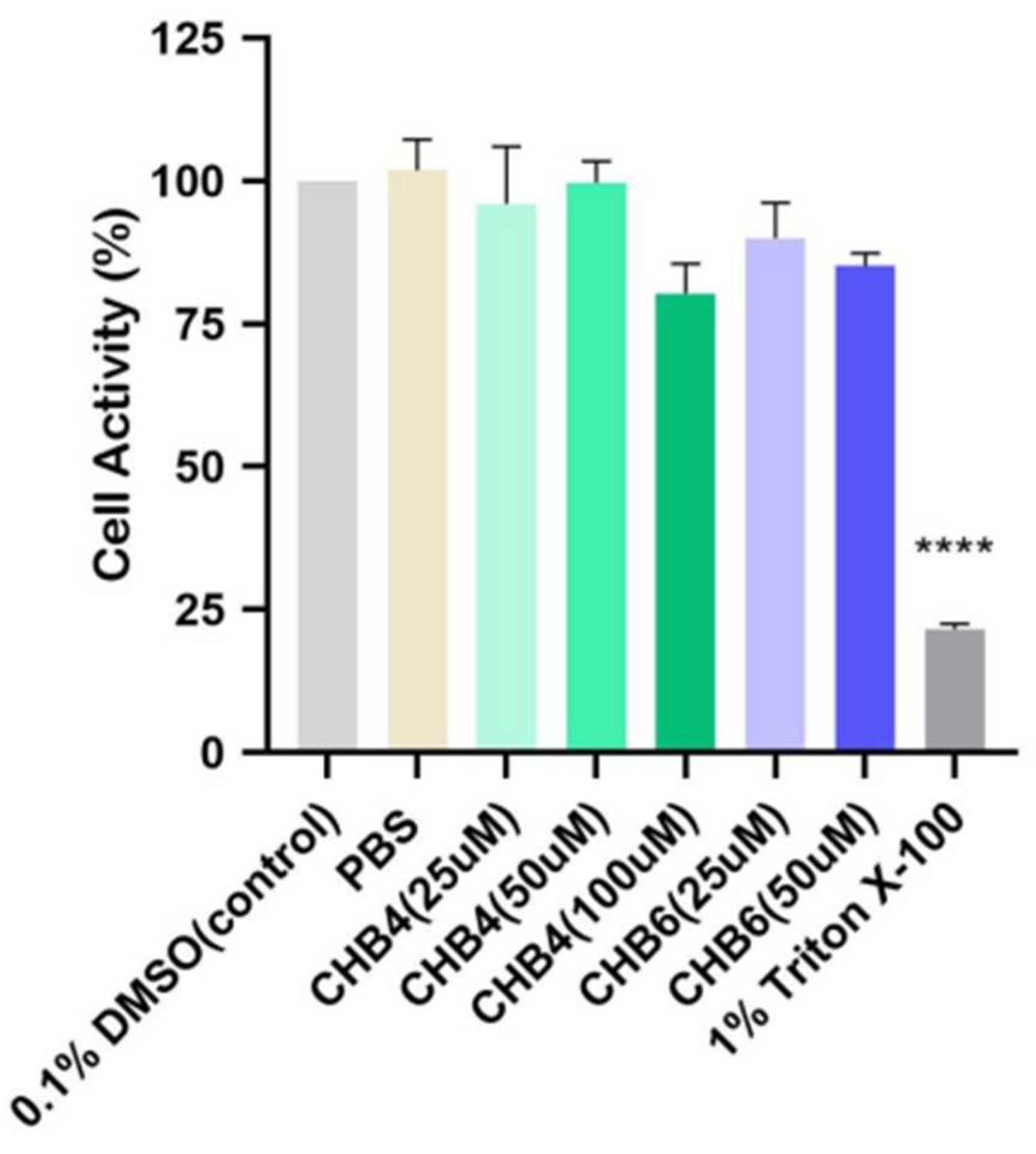
2.7. In Vitro Study on Reconstructed Human Epidermal Model
3. Discussion
3.1. Conclusions
3.2. Summary Points
- Vulvovaginal candidiasis (VVC) is often referred to as, “vaginal yeast infection”.
- Two potent and novel N-terpenyl analogs of ebselen (EB), named CHB4 and CHB6, were characterized for anti-Candida efficacy.
- CHB4 and CHB6 inhibited the growth of two clinical isolates of C. albicans in vitro.
- CHB4 inhibited the plasma membrane H+-ATPase (Pma1p), an essential protein in C. albicans, indicating that this mechanism is involved, at least in part, in its antifungal activity.
- EB and the N-terpenyl analogs CHB4 and CHB6 all inhibited the Na+, K+-ATPase, indicating that this class of compounds has the potential to exhibit off-target effects.
- CHB4 and CHB6 were not irritating to reconstructed human epidermis.
- CHB4 and CHB6, administered intravaginally, both showed potent efficacy in the mouse model of VVC.
- Histopathologic analysis of vaginal tissues from CHB4 or CHB6-treated mice showed less inflammation and tissue injury than tissues obtained from infected but untreated control mice.
- CHB4 and CHB6 represent new N-terpenyl analogs of EB that should be investigated as potential treatments for human VVC.
4. Materials and Methods
4.1. Chemicals and Strains
4.2. Determination of Minimum Inhibitory Concentration (MIC)
4.3. Evaluation of Medium Acidification Effects
4.4. Cell Viability Assay
4.5. Antifungal Activity in the Mouse Model of VVC
- Group 1: Naive: non-infected (no treatment);
- Group 2: 6th day infected: estrogenized infected (inoculated with yeast) with no additional treatments (baseline value for infection);
- Group 3: MICO (2% cream): estrogenized infected (inoculated with yeast) treated with 27 µL of 2% MICO cream (positive control);
- Group 4: Vehicle: estrogenized infected (inoculated with yeast) treated with a 27 µL mixture of DMA (2.7 µL) + 1% HPMC (24.3 µL);
- Group 5: CHB4 (12.5 mg/kg): estrogenized infected (inoculated with yeast) treated with 27 µL suspension of 0.25 mg CHB4 dissolved in DMA (2.7 µL) + 1% HPMC (24.3 µL).
- Group 6: CHB6 (12.5 mg/kg): estrogenized infected (inoculated with yeast) treated with 27 µL suspension of 0.25 mg CHB6 dissolved in DMA (2.7 µL) + 1% HPMC (24.3 µL).
4.6. Immunohistochemistry
4.7. Acute Toxicity Study
4.8. In Vitro Skin Irritation Test
4.9. Statistical Analysis
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yano, J.; Sobel, J.D.; Nyirjesy, P.; Sobel, R.; Williams, V.L.; Yu, Q.; Noverr, M.C.; Fidel, P.L., Jr. Current patient perspectives of vulvovaginal candidiasis: Incidence, symptoms, management and post-treatment outcomes. BMC Women’s Health 2019, 19, 48. [Google Scholar] [CrossRef] [PubMed]
- Neal, C.M.; Martens, M.G. Clinical challenges in diagnosis and treatment of recurrent vulvovaginal candidiasis. SAGE Open Med. 2022, 10. [Google Scholar] [CrossRef] [PubMed]
- Sobel, J.D. Pathogenesis and treatment of recurrent vulvovaginal candidiasis. Clin. Infect. Dis. 1992, 14 (Suppl. S1), S148–S153. [Google Scholar] [CrossRef]
- Gaziano, R.; Sabbatini, S.; Monari, C. The interplay between Candida albicans, vaginal mucosa, host Immunity and resident microbiota in health and disease: An overview and future perspectives. Microorganisms 2023, 11, 1211. [Google Scholar] [CrossRef]
- Denning, D.W.; Kneale, M.; Sobel, J.D.; Rautemaa-Richardson, R. Global burden of recurrent vulvovaginal candidiasis: A systematic review. Lancet Infect. Dis. 2018, 18, e339–e347. [Google Scholar] [CrossRef]
- Willems, H.M.E.; Ahmed, S.S.; Liu, J.; Xu, Z.; Peters, B.M. Vulvovaginal candidiasis: A current understanding and burning questions. J. Fungi 2020, 6, 27. [Google Scholar] [CrossRef]
- Cassone, A. Vulvovaginal Candida albicans infections: Pathogenesis, immunity and vaccine prospects. BJOG 2015, 122, 785–794. [Google Scholar] [CrossRef] [PubMed]
- Kumar, D.; Kumar, A. Cellular attributes of Candida albicans biofilm-associated in resistance against multidrug and host immune system. Microb. Drug Resist. 2023, 29, 423–437. [Google Scholar] [CrossRef] [PubMed]
- File, B.; Sobel, R.; Becker, M.; Nyirjesy, P. Fluconazole-resistant Candida albicans vaginal infections at a referral center and treated with boric acid. J. Low. Genit. Tract Dis. 2023, 27, 262–265. [Google Scholar] [CrossRef]
- Sobel, J.D. Resistance to fluconazole of Candida albicans in vaginal isolates: A 10-year study in a clinical referral center. Antimicrob. Agents Chemother. 2023, 67, e0018123. [Google Scholar] [CrossRef]
- Menon, S.; Liang, X.; Vartak, R.; Patel, K.; Di Stefano, A.; Cacciatore, I.; Marinelli, L.; Billack, B. Antifungal activity of novel formulations based on terpenoid prodrugs against C. albicans in a mouse model. Pharmaceutics 2021, 13, 633. [Google Scholar] [CrossRef] [PubMed]
- Ray, A.; Ray, S.; George, A.T.; Swaminathan, N. Interventions for prevention and treatment of vulvovaginal candidiasis in women with HIV infection. Cochrane Database Syst. Rev. 2011, CD008739. [Google Scholar] [CrossRef]
- Pienaar, E.D.; Young, T.; Holmes, H. Interventions for the prevention and management of oropharyngeal candidiasis associated with HIV infection in adults and children. Cochrane Database Syst. Rev. 2006, CD003940. [Google Scholar] [CrossRef]
- Garland, M.; Hryckowian, A.J.; Tholen, M.; Bender, K.O.; Van Treuren, W.W.; Loscher, S.; Sonnenburg, J.L.; Bogyo, M. The clinical drug ebselen attenuates inflammation and promotes microbiome recovery in mice after antibiotic treatment for CDI. Cell Rep. Med. 2020, 1, 100005. [Google Scholar] [CrossRef]
- Lee, M.Y.; Kabara, L.L.; Swiderski, D.L.; Raphael, Y.; Duncan, R.K.; Kim, Y.H. ROS scavenger, ebselen, has no preventive effect in new hearing loss model using a cholesterol-chelating agent. J. Audiol. Otol. 2019, 23, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Felli Kubiça, T.; Bedin Denardi, L.; Silva de Loreto, É.; Zeni, G.; Weiblen, C.; Oliveira, V.; Morais Santurio, J.; Hartz Alves, S. In vitro activity of diphenyl diselenide and ebselen alone and in combination with antifungal agents against Trichosporon asahii. Mycoses 2019, 62, 428–433. [Google Scholar] [CrossRef]
- Orie, N.N.; Warren, A.R.; Basaric, J.; Lau-Cam, C.; Piętka-Ottlik, M.; Młochowski, J.; Billack, B. In vitro assessment of the growth and plasma membrane H+-ATPase inhibitory activity of ebselen and structurally related selenium- and sulfur-containing compounds in Candida albicans. J. Biochem. Mol. Toxicol. 2017, 31, 6. [Google Scholar] [CrossRef] [PubMed]
- Chan, G.; Hardej, D.; Santoro, M.; Lau-Cam, C.; Billack, B. Evaluation of the antimicrobial activity of ebselen: Role of the yeast plasma membrane H+-Atpase. J. Biochem. Mol. Toxicol. 2007, 21, 252–264. [Google Scholar] [CrossRef] [PubMed]
- Billack, B.; Santoro, M.; Lau-Cam, C. Growth inhibitory action of ebselen on fluconazole-resistant Candida albicans: Role of the plasma membrane H+-Atpase. Microb. Drug Resist. 2009, 15, 77–83. [Google Scholar] [CrossRef]
- Wang, J.; Wang, P.; Dong, C.; Zhao, Y.; Zhou, J.; Yuan, C.; Zou, L. Mechanisms of ebselen as a therapeutic and its pharmacology applications. Future Med. Chem. 2020, 12, 2141–2160. [Google Scholar] [CrossRef] [PubMed]
- Nogueira, C.W.; Barbosa, N.V.; Rocha, J.B.T. Toxicology and pharmacology of synthetic organoselenium compounds: An update. Arch. Toxicol. 2021, 95, 1179–1226. [Google Scholar] [CrossRef] [PubMed]
- Santi, C.; Scimmi, C.; Sancineto, L. Ebselen and analogues: Pharmacological properties and synthetic strategies for their preparation. Molecules 2021, 26, 4230. [Google Scholar] [CrossRef] [PubMed]
- Obieziurska, M.; Pacuła, A.J.; Długosz-Pokorska, A.; Krzemiński, M.; Janecka, A.; Ścianowski, J. Bioselectivity induced by chirality of new terpenyl organoselenium compounds. Materials 2019, 12, 3579. [Google Scholar] [CrossRef]
- De Vasconcelos, L.C.; Sampaio, F.C.; Albuquerque, A.J.; Vasconcelos, L.C. Cell viability of Candida albicans against the antifungal activity of thymol. Braz. Dent. J. 2014, 25, 277–281. [Google Scholar] [CrossRef] [PubMed]
- Perlin, D.S.; Brown, C.L.; Haber, J.E. Membrane potential defect in hygromycin b-resistant pma1 mutants of Saccharomyces cerevisiae. J. Biol. Chem. 1988, 263, 18118–18122. [Google Scholar] [CrossRef]
- Rehring, J.F.; Bui, T.M.; Galán-Enríquez, C.S.; Urbanczyk, J.M.; Ren, X.; Wiesolek, H.L.; Sullivan, D.P.; Sumagin, R. Released myeloperoxidase attenuates neutrophil migration and accumulation in inflamed tissue. Front. Immunol. 2021, 12, 654259. [Google Scholar] [CrossRef]
- Organisation for Economic Cooperation and Development (OECD). Test Guideline No. 439, “In Vitro Skin Irritation (Reconstructed Human Epidermis Test Method)”; Organisation for Economic Cooperation and Development: Paris, France, 2021. [Google Scholar]
- Chapple, A. Vaginal thrush: Perceptions and experiences of women of South Asian descent. Health Educ. Res. 2001, 16, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Sobel, J.D.; Sobel, R. Current treatment options for vulvovaginal candidiasis caused by azole-resistant Candida species. Expert Opin. Pharmacother. 2018, 19, 971–977. [Google Scholar] [CrossRef]
- Benelli, J.L.; Poester, V.R.; Munhoz, L.S.; Melo, A.M.; Trápaga, M.R.; Stevens, D.A.; Xavier, M.O. Ebselen and diphenyl diselenide against fungal pathogens: A systematic review. Med. Mycol. 2021, 59, 409–421. [Google Scholar] [CrossRef] [PubMed]
- Menon, S.; Vartak, R.; Patel, K.; Billack, B. Evaluation of the antifungal activity of an ebselen-loaded nanoemulsion in a mouse model of vulvovaginal candidiasis. Nanomed. Nanotechnol. Biol. Med. 2021, 37, 102428. [Google Scholar] [CrossRef]
- Vartak, R.; Patki, M.; Menon, S.; Jablonski, J.; Mediouni, S.; Fu, Y.; Valente, S.T.; Billack, B.; Patel, K. β-cyclodextrin polymer/Soluplus® encapsulated Ebselen ternary complex (EβpolySol) as a potential therapy for vaginal candidiasis and pre-exposure prophylactic for HIV. Int. J. Pharm. 2020, 589, 119863. [Google Scholar] [CrossRef]
- Pacuła, A.J.; Kaczor, K.B.; Antosiewicz, J.; Janecka, A.; Długosz, A.; Janecki, T.; Wojtczak, A.; Ścianowski, J. New chiral ebselen analogues with antioxidant and cytotoxic potential. Molecules 2017, 22, 492. [Google Scholar] [CrossRef]
- Pacuła, A.J.; Kaczor, K.B.; Wojtowicz, A.; Antosiewicz, J.; Janecka, A.; Długosz, A.; Janecki, T.; Ścianowski, J. New glutathione peroxidase mimetics-Insights into antioxidant and cytotoxic activity. Bioorg. Med. Chem. 2017, 25, 126–131. [Google Scholar] [CrossRef]
- Palmgren, M.; Morsomme, P. The plasma membrane H(+)-Atpase, a simple polypeptide with a long History. Yeast 2019, 36, 201–210. [Google Scholar] [CrossRef] [PubMed]
- Tung, T.T.; Nielsen, J. Drug discovery and development on Pma1, where are we now? A critical review from 1995 to 2022. ChemMedChem 2022, 17, e202200356. [Google Scholar] [CrossRef] [PubMed]
- Monk, B.C.; Perlin, D.S. Fungal plasma membrane proton pumps as promising new antifungal targets. Crit. Rev. Microbiol. 1994, 20, 209–223. [Google Scholar] [CrossRef] [PubMed]
- Franz, R.; Ruhnke, M.; Morschhauser, J. Molecular aspects of fluconazole resistance development in Candida albicans. Mycoses 1999, 42, 453–458. [Google Scholar] [CrossRef] [PubMed]
- Miller, E.A.; Beasley, D.E.; Dunn, R.R.; Archie, E.A. Lactobacilli dominance and vaginal pH: Why is the human vaginal microbiome unique? Front. Microbiol. 2016, 7, 1936. [Google Scholar] [CrossRef]
- Fidel, P.L., Jr. The neutral vaginal pH in mice that is typical of most mammalian species should not deter research using experimental murine models of Candida vaginitis. Infect. Immun. 2021, 89, e00624-20. [Google Scholar] [CrossRef]
- Garvey, E.P.; Hoekstra, W.J.; Schotzinger, R.J.; Sobel, J.D.; Lilly, E.A.; Fidel, P.L., Jr. Efficacy of the clinical agent VT-1161 against fluconazole-sensitive and -resistant Candida albicans in a murine model of vaginal candidiasis. Antimicrob. Agents Chemother. 2015, 59, 5567–5573. [Google Scholar] [CrossRef] [PubMed]
- Bisht, N.; Phalswal, P.; Khanna, P.K. Selenium nanoparticles: A review on synthesis and biomedical applications. Mater. Adv. 2022, 3, 1415–1431. [Google Scholar] [CrossRef]
- Kumar, A.; Prasad, K.S. Role of nano-selenium in health and environment. J. Biotechnol. 2021, 325, 152–163. [Google Scholar] [CrossRef] [PubMed]
- Mal’tseva, V.N.; Goltyaev, M.V.; Turovsky, E.A.; Varlamova, E.G. Immunomodulatory and Anti-Inflammatory Properties of Selenium-Containing Agents: Their Role in the Regulation of Defense Mechanisms against COVID-19. Int. J. Mol. Sci. 2022, 23, 2360. [Google Scholar] [CrossRef] [PubMed]
- Yano, J.; Fidel, P.L., Jr. Protocols for vaginal inoculation and sample collection in the experimental mouse model of Candida vaginitis. J. Vis. Exp. 2011, 58, 3382. [Google Scholar] [CrossRef]
- Fidel, P.L., Jr.; Cutright, J.; Steele, C. Effects of reproductive hormones on experimental vaginal candidiasis. Infect. Immun. 2000, 68, 651–657. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | Molecular Weight (g/mol) | Water Solubility (µg/mL) |
|---|---|---|
| CHB1 | 334.31 | 4.78 |
| CHB2 | 334.31 | 4.78 |
| CHB3 | 334.31 | 6.07 |
| CHB4 | 334.31 | 4.78 |
| CHB5 | 334.31 | 6.30 |
| CHB6 | 336.33 | 2.75 |
| EB | 274.18 | 33.00 |
| Compounds | C. albicans S1 (µM) | C. albicans S2 (µM) | ||
|---|---|---|---|---|
| 24 h | 48 h | 24 h | 48 h | |
| CHB1 | 12.5 | 12.5 | 3.1 | 3.1 |
| CHB2 | 12.5 | 12.5 | 12.5 | 12.5 |
| CHB3 | 25.0 | 25.0 | 50.0 | 25.0 |
| CHB4 | 12.5 | 12.5 | 3.1 | 3.1 |
| CHB5 | 25.0 | 12.5 | 12.5 | 25.0 |
| CHB6 | 3.1 | 3.1 | 3.1 | 3.1 |
| EB | 25.0 | 25.0 | 25.0 | 25.0 |
| FLU | 25.0 | 25.0 | >100 | >100 |
| Compounds | C. albicans S1 (µM) | C. albicans S2 (µM) | ||
|---|---|---|---|---|
| 24 h | 48 h | 24 h | 48 h | |
| CHB1 | 12.5 | 6.3 | 12.5 | 12.5 |
| CHB2 | 12.5 | 12.5 | 6.3 | 3.1 |
| CHB3 | 12.5 | 25.0 | 25.0 | 25.0 |
| CHB4 | 6.3 | 12.5 | 3.1 | 3.1 |
| CHB5 | 12.5 | 25.0 | 12.5 | 12.5 |
| CHB6 | 3.1 | 3.1 | 6.3 | 6.3 |
| EB | 25.0 | 25.0 | 25.0 | 25.0 |
| FLU | >100 | >100 | >100 | >100 |
| Compounds | MTT Assay IC50(µM) KB-3-1 Cells | Turbidity Assay IC50 (µM) | |
|---|---|---|---|
| C. albicans S1 | C. albicans S2 | ||
| CHB1 | 28.1 ± 1.5 | 7.3 ± 0.1 | 1.8 ± 0.1 |
| CHB2 | 25.5 ± 1.9 | 7.6 ± 0.1 | 6.2 ± 0.1 |
| CHB3 | 65.7 ± 4.4 | 18.1 ± 0.2 | 15.1 ± 0.2 |
| CHB4 | 17.9 ± 3.1 | 7.7 ± 0.1 | 1.8 ± 0.1 |
| CHB5 | 94.1 ± 4.7 | 7.5 ± 0.1 | 14.4 ± 0.5 |
| CHB6 | 48.5 ± 5.9 | 1.8 ± 0.1 | 1.8 ± 0.1 |
| EB | 89.74 ± 3.42 | 17.06 ± 1.01 | 14.28 ± 0.46 |
| FLU | >1000 | 6.01 ± 0.10 | >100 |
| Compound | IC50MA, µM |
|---|---|
| CHB1 | >30 |
| CHB2 | >30 |
| CHB3 | >30 |
| CHB4 | 13.8 ± 1.1 |
| CHB5 | >30 |
| CHB6 | >30 |
| EB | 12.5 + 1.1 |
| Groups | Log CFU/100 μL | Remaining C. albicans S1 Compared to Control (100%) |
|---|---|---|
| 6th day infected (control) | 5.628 | 100.000 |
| Miconazole (MICO) | 4.470 | 6.950 |
| Vehicle | 5.630 | 99.541 |
| CHB4 | 4.448 | 6.607 |
| CHB6 | 3.655 | 1.064 |
| Naive | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liang, X.; Pacuła-Miszewska, A.J.; Obieziurska-Fabisiak, M.; Vartak, R.; Mao, G.; Patel, K.; Fedosova, N.U.; Ścianowski, J.; Billack, B. Selected N-Terpenyl Organoselenium Compounds Possess Antimycotic Activity In Vitro and in a Mouse Model of Vulvovaginal Candidiasis. Molecules 2023, 28, 7377. https://doi.org/10.3390/molecules28217377
Liang X, Pacuła-Miszewska AJ, Obieziurska-Fabisiak M, Vartak R, Mao G, Patel K, Fedosova NU, Ścianowski J, Billack B. Selected N-Terpenyl Organoselenium Compounds Possess Antimycotic Activity In Vitro and in a Mouse Model of Vulvovaginal Candidiasis. Molecules. 2023; 28(21):7377. https://doi.org/10.3390/molecules28217377
Chicago/Turabian StyleLiang, Xiuyi, Agata J. Pacuła-Miszewska, Magdalena Obieziurska-Fabisiak, Richa Vartak, Ganming Mao, Ketankumar Patel, Natalya U. Fedosova, Jacek Ścianowski, and Blase Billack. 2023. "Selected N-Terpenyl Organoselenium Compounds Possess Antimycotic Activity In Vitro and in a Mouse Model of Vulvovaginal Candidiasis" Molecules 28, no. 21: 7377. https://doi.org/10.3390/molecules28217377
APA StyleLiang, X., Pacuła-Miszewska, A. J., Obieziurska-Fabisiak, M., Vartak, R., Mao, G., Patel, K., Fedosova, N. U., Ścianowski, J., & Billack, B. (2023). Selected N-Terpenyl Organoselenium Compounds Possess Antimycotic Activity In Vitro and in a Mouse Model of Vulvovaginal Candidiasis. Molecules, 28(21), 7377. https://doi.org/10.3390/molecules28217377