

Structural Analysis and Antioxidant and Immunoregulatory Activities of an Exopolysaccharide Isolated from Bifidobacterium longum subsp. longum XZ01

Abstract
:
1. Introduction
2. Results and Discussion
2.1. Identification of Bacterial Strain XZ01
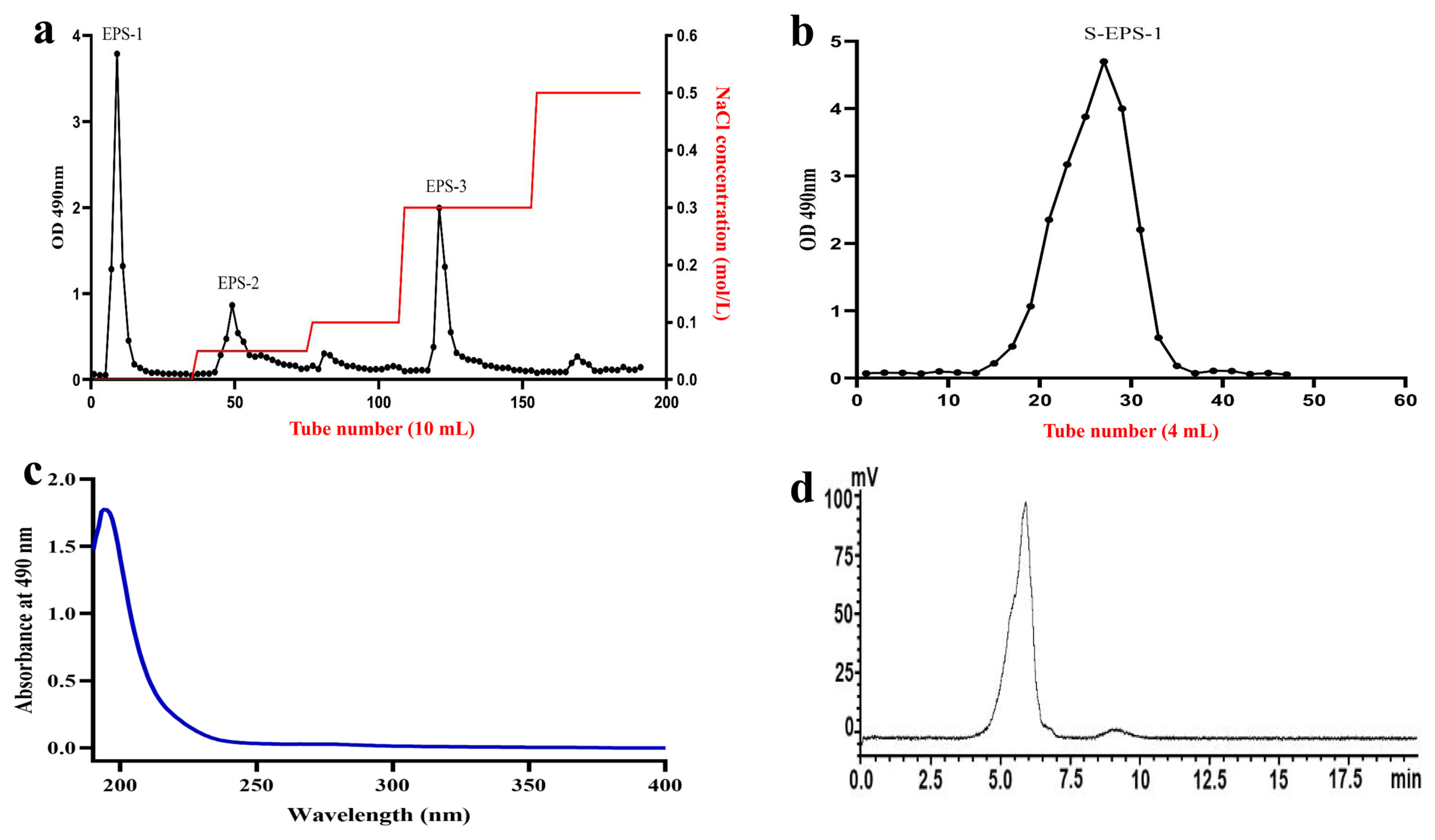
2.2. Isolation and Purification of S-EPS-1
2.3. Purity and Molecular Weight Determination of S-EPS-1
2.4. Structural Features
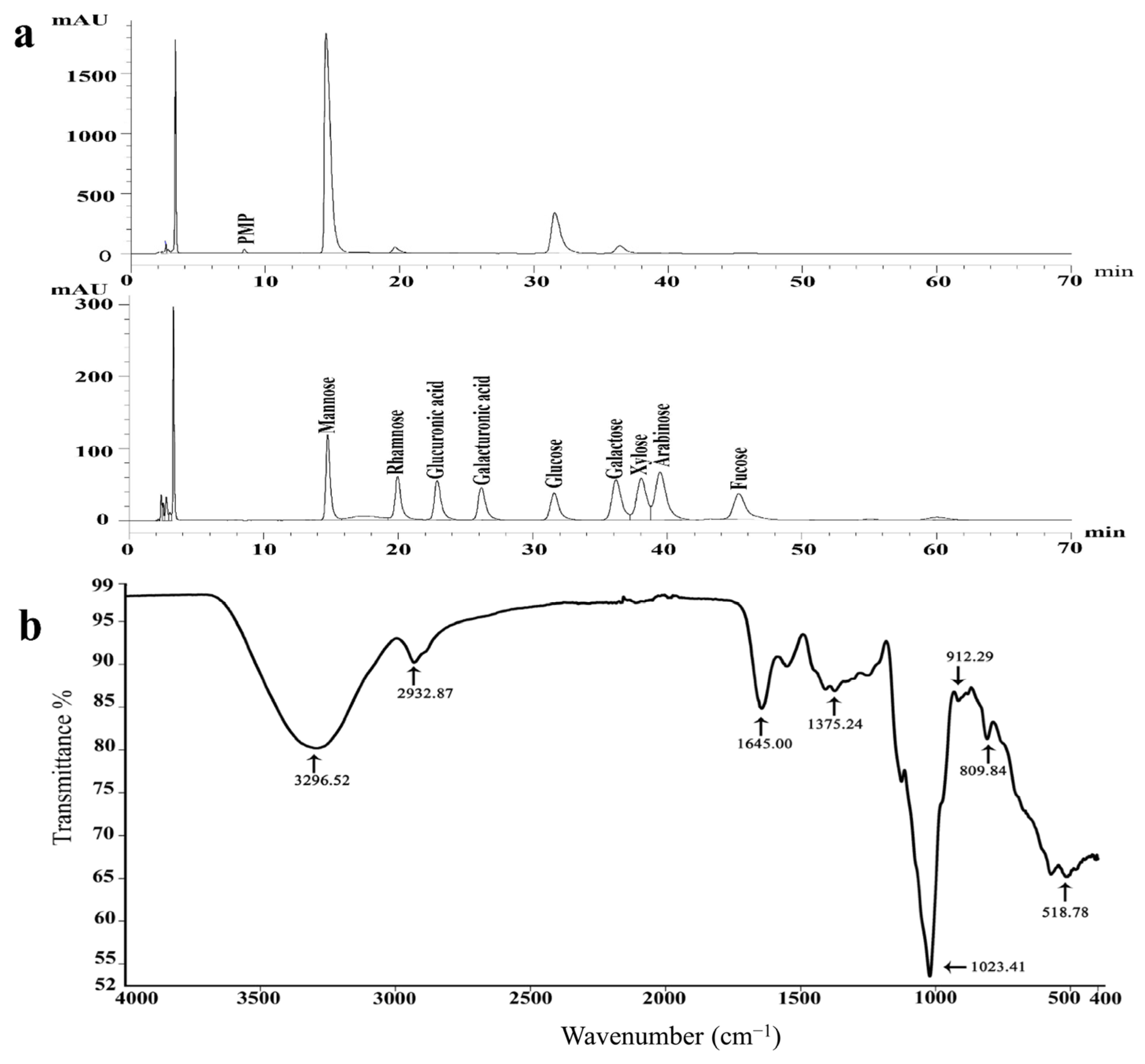
2.4.1. Monosaccharide Composition of S-EPS-1
2.4.2. FT-IR Spectrum of S-EPS-1
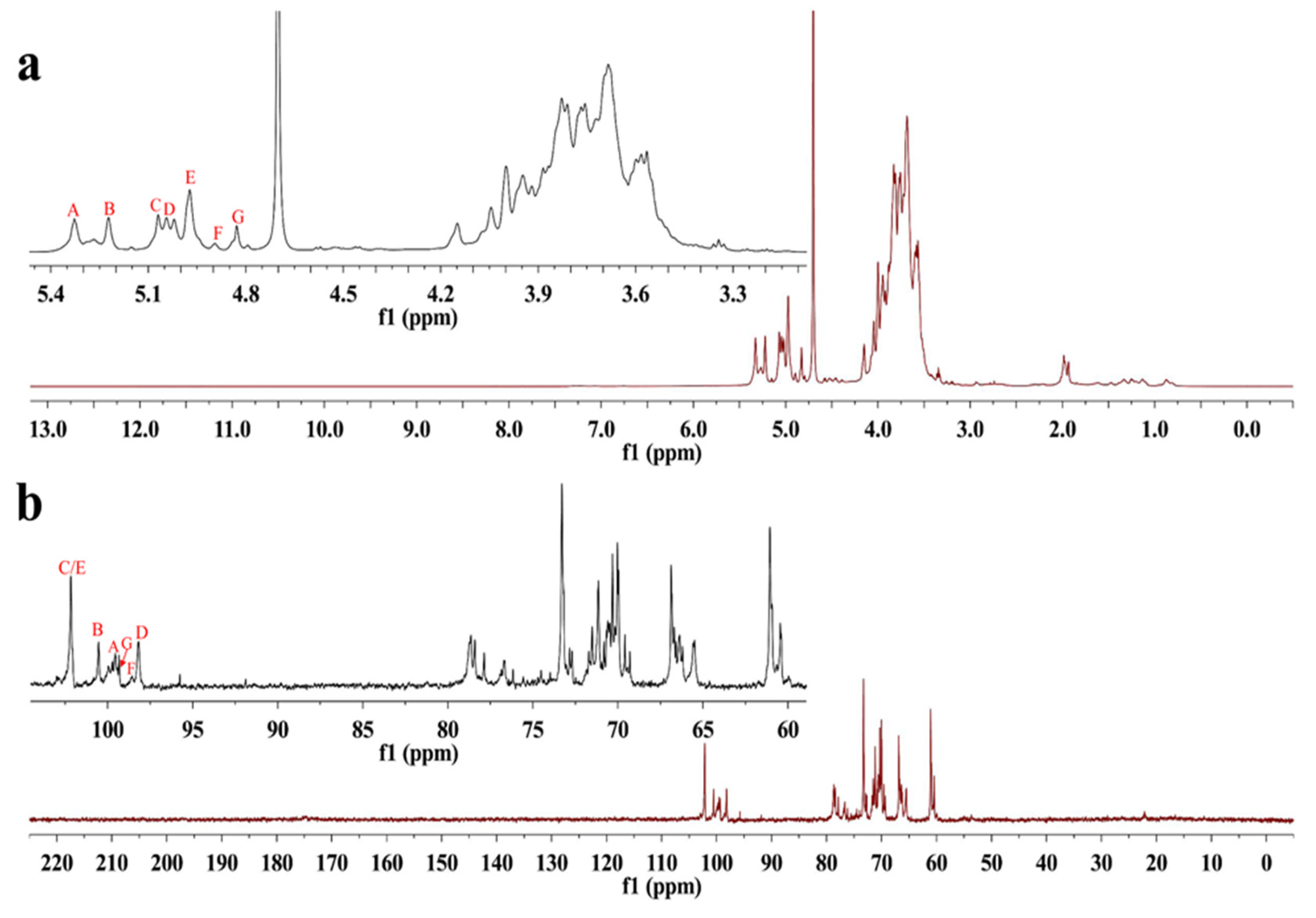
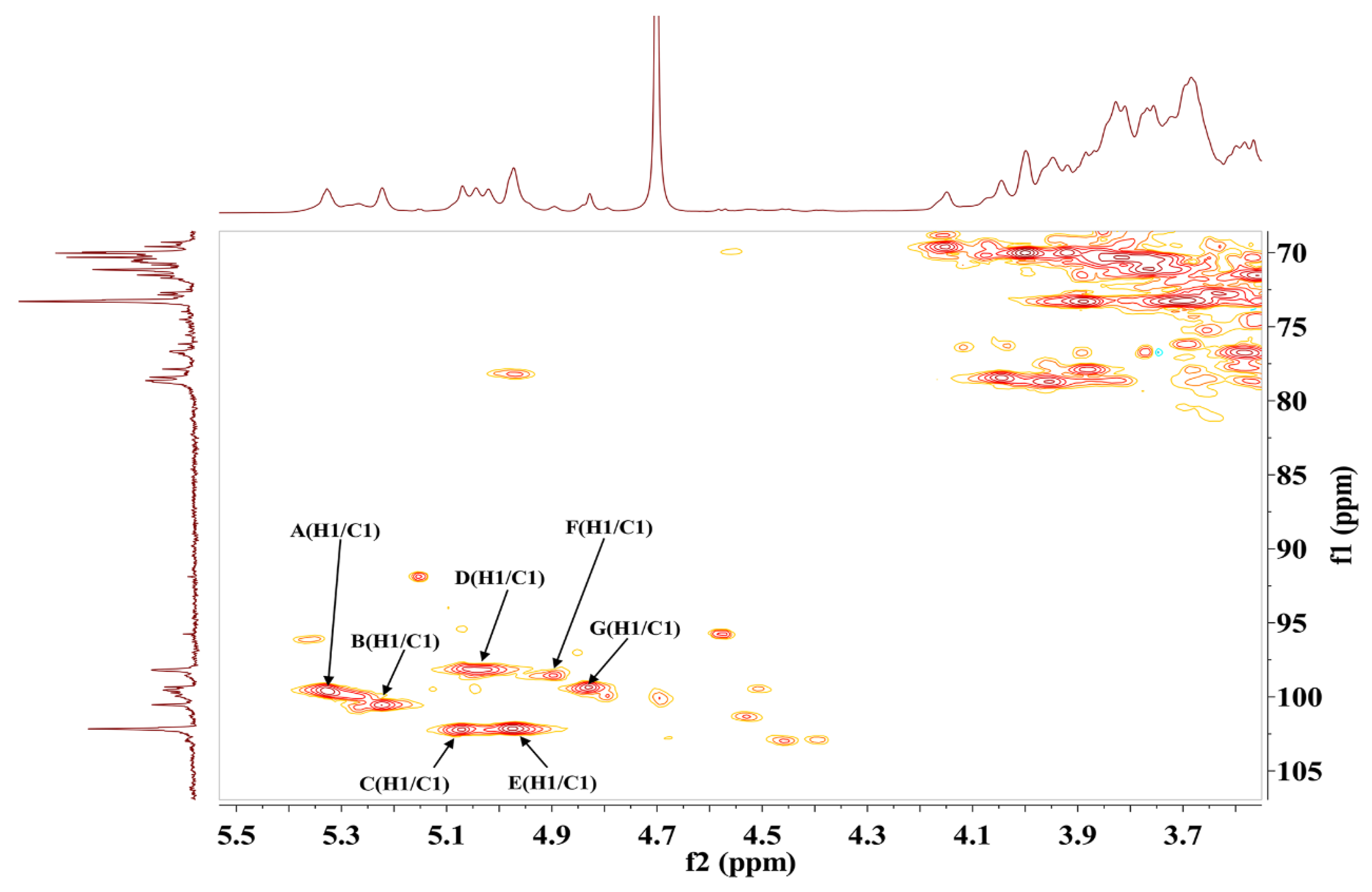
2.4.3. NMR Analysis of S-EPS-1
2.4.4. SEM Analysis
2.5. Immunoregulatory Activities of S-EPS-1
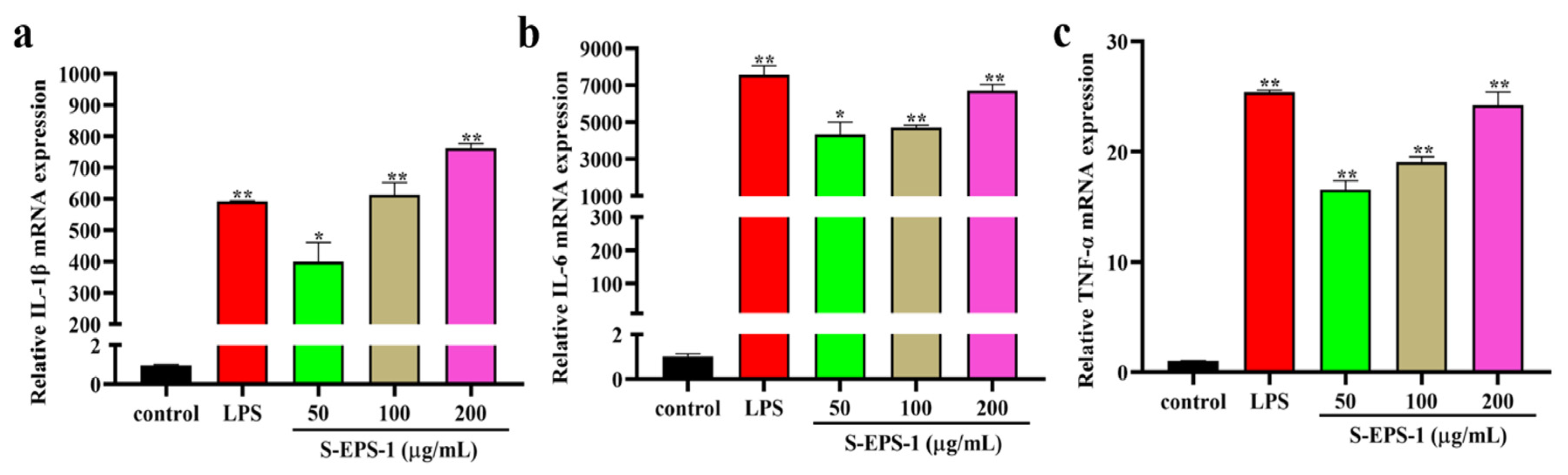
2.5.1. Effect on Cytokine Expressions
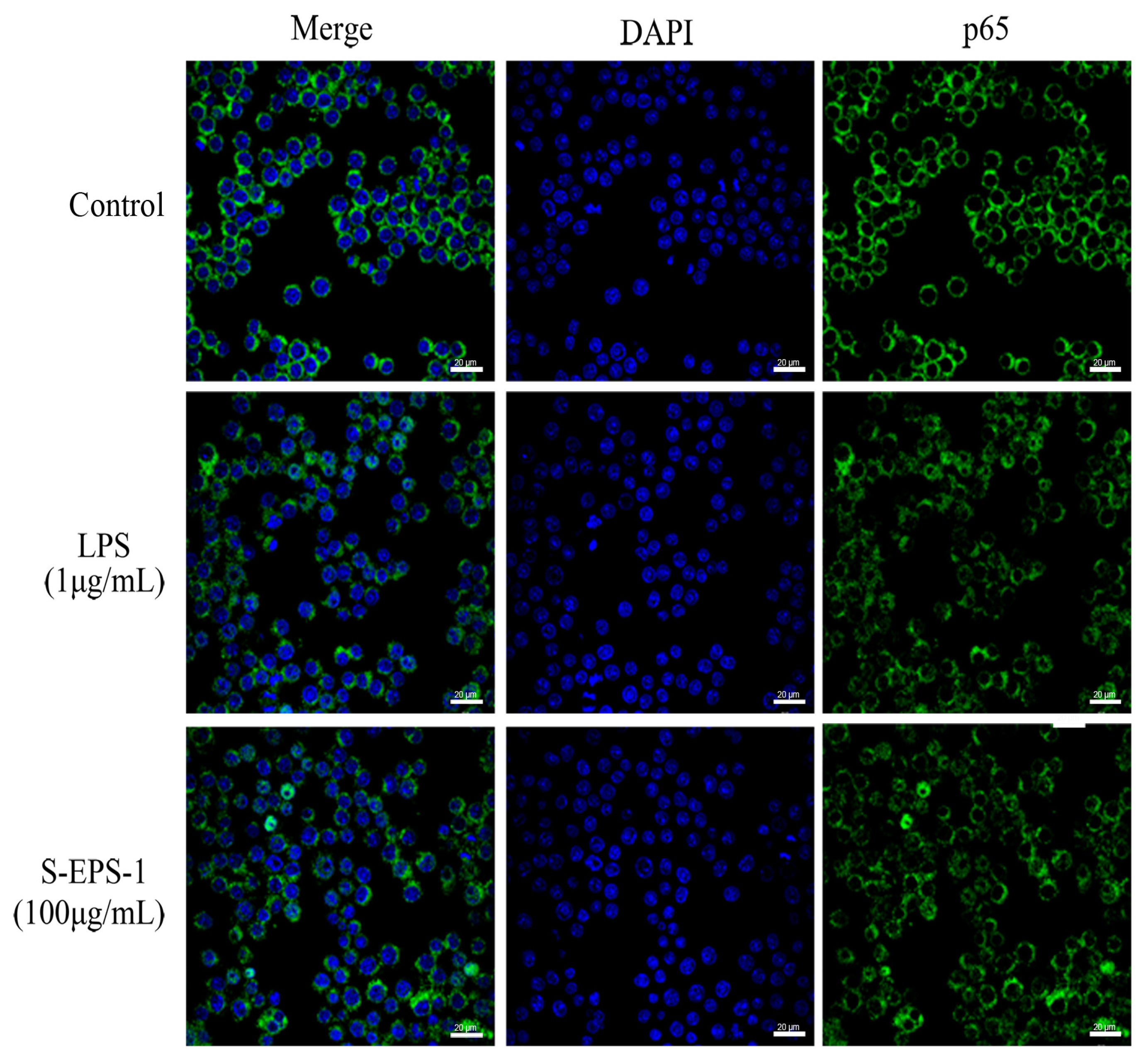
2.5.2. Effect on Nuclear Translocation of p65
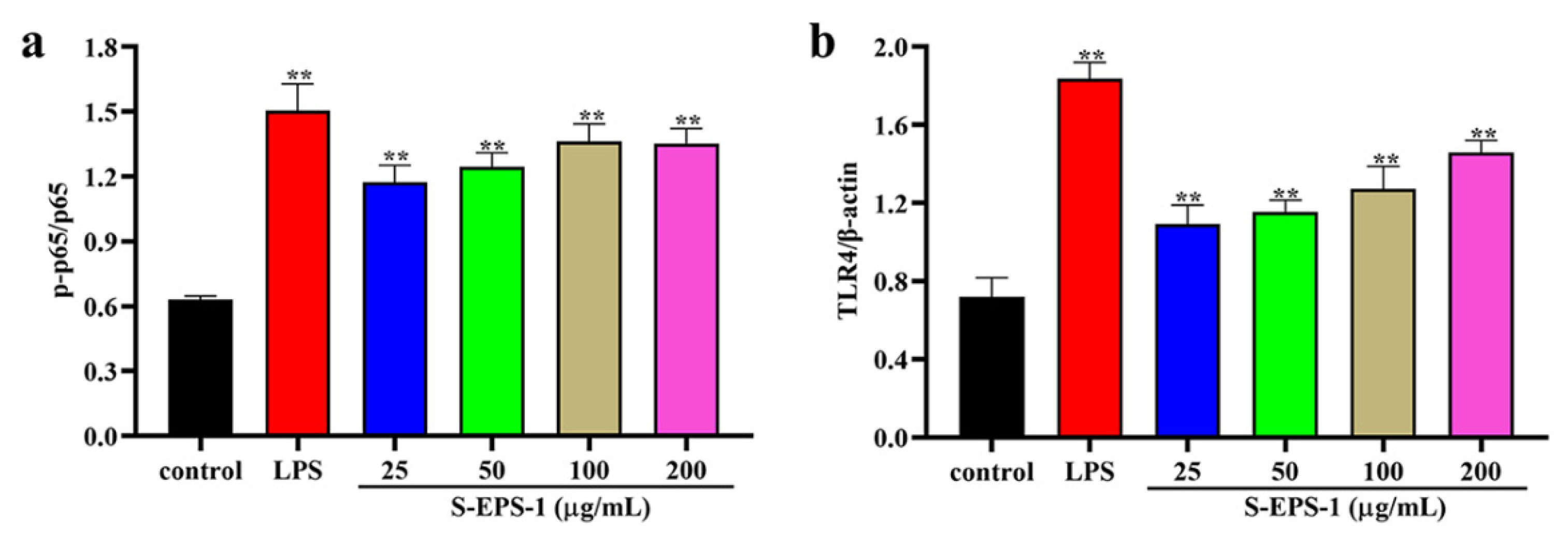
2.5.3. TLR4/NF-κB Signaling Pathway
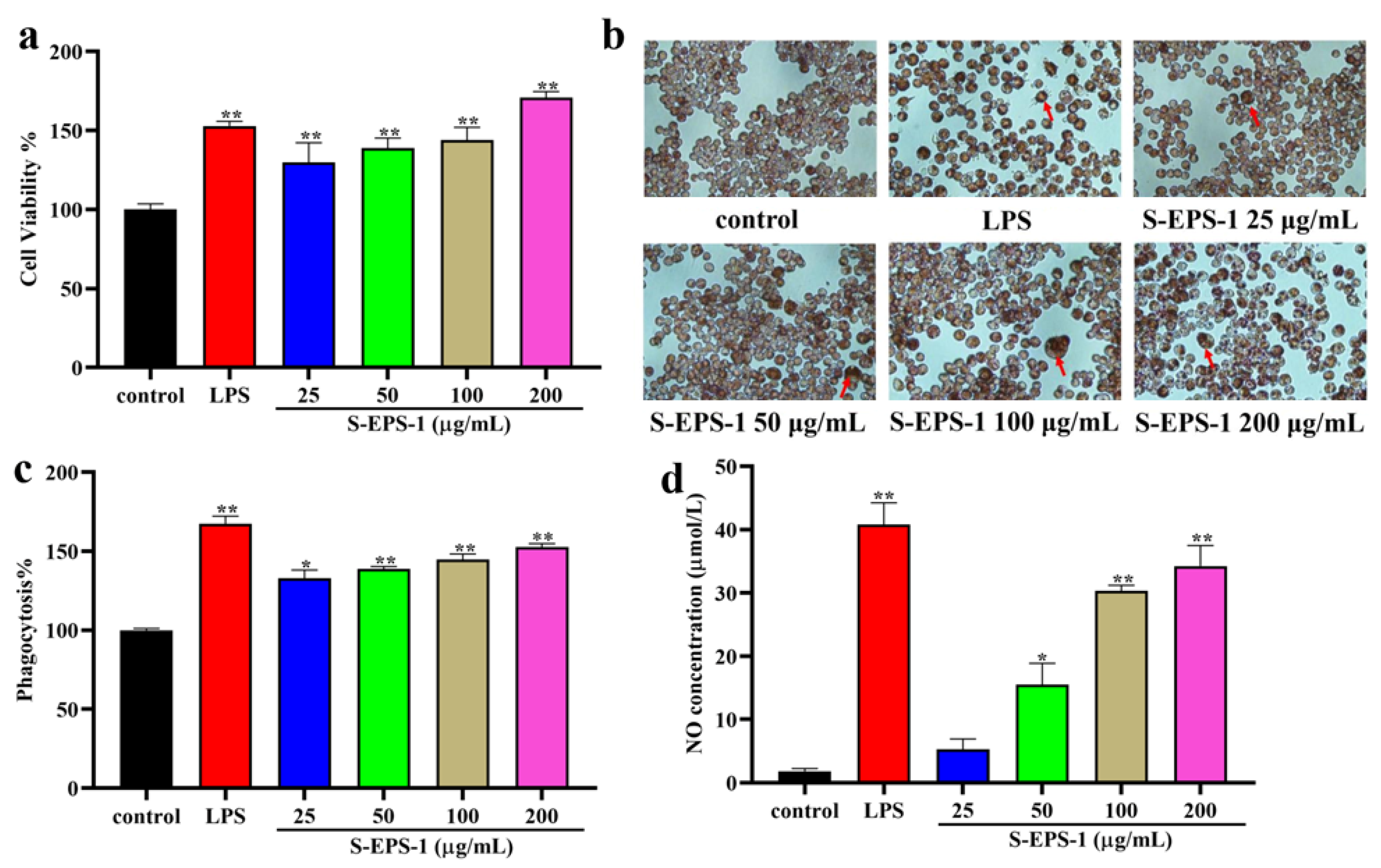
2.5.4. Effect on Phagocytic Activity
2.6. Cytoprotective Effects of S-EPS-1 on Cell Viability
2.6.1. Cell Viability
2.6.2. NO Production Regulation
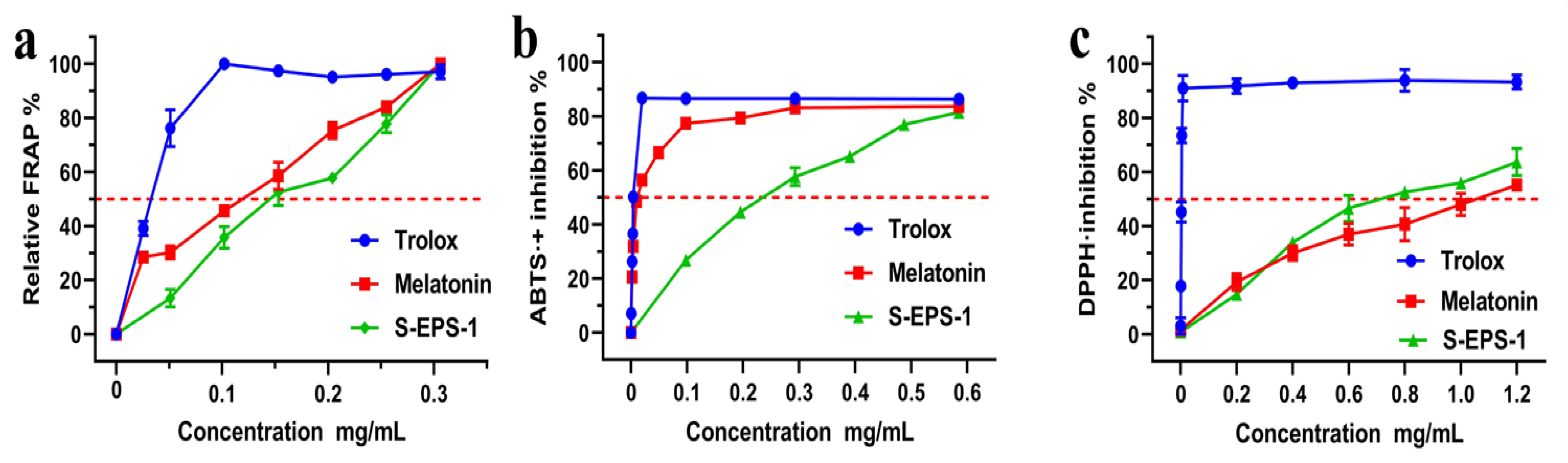
2.7. Antioxidant Activity of S-EPS-1
3. Materials and Methods
3.1. Materials and Reagents
3.2. Culture Condition and Identification of Bacterial Strain XZ01
3.3. Isolation and Purification of S-EPS-1
3.4. Chemical Properties and Determination of Molecular Weight
3.5. Structural Analysis
3.5.1. Monosaccharide Composition Analysis
3.5.2. UV-Vis, FT-IR, and NMR Spectra Analysis
3.5.3. SEM Analysis
3.6. Immunoregulatory Activities of S-EPS-1
3.6.1. Cell Culture and Treatment
3.6.2. Determination of Cytokine Expression
3.6.3. Immunofluorescence Assay
3.6.4. Determination of Phagocytic Activity
3.7. Cytoprotective Effects of S-EPS-1
3.7.1. Cytotoxicity Assay
3.7.2. Determination of NO Production
3.8. Antioxidant Activity
3.8.1. Ferric-Ion Reducing Antioxidant Power
3.8.2. Scavenging Ability of S-EPS-1 on ABTS+•
3.8.3. Scavenging Ability of S-EPS-1 on DPPH• Radicals
3.9. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, N.; Jia, G.; Wang, C.; Chen, M.; Xie, F.; Nepovinnykh, N.V.; Goff, H.D.; Guo, Q. Structural characterisation and immunomodulatory activity of exopolysaccharides from liquid fermentation of Monascus purpureus (Hong Qu). Food Hydrocoll. 2020, 103, 105636. [Google Scholar] [CrossRef]
- Rajoka, M.S.R.; Mehwish, H.M.; Kitazawa, H.; Barba, F.J.; Berthelot, L.; Umair, M.; Zhu, Q.; He, Z.; Zhao, L. Techno-functional properties and immunomodulatory potential of exopolysaccharide from Lactiplantibacillus plantarum MM89 isolated from human breast milk. Food Chem. 2021, 377, 131954. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.H.; Pan, T.M.; Wu, Y.J.; Chang, S.J.; Chang, M.S.; Hu, C.Y. Exopolysaccharide activities from probiotic bifidobacterium: Immunomodulatory effects (on J774A.1 macrophages) and antimicrobial properties. Int. J. Food Microbiol. 2010, 144, 104–110. [Google Scholar] [CrossRef]
- Lü, J.; Lin, P.H.; Yao, Q.; Chen, C. Chemical and molecular mechanisms of antioxidants: Experimental approaches and model systems. J. Cell. Mol. Med. 2010, 14, 840–860. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.P.; Chandra, A.; Mahdi, F.; Ray, A.; Sharma, P. Reconvene and reconnect the antioxidant hypothesis in Human health and disease. Ind. J. Clin. Biochem. 2010, 25, 225–243. [Google Scholar] [CrossRef] [PubMed]
- Weitzman, S.A.; Gordon, L.I. Inflammation and cancer: Role of phagocyte-generated oxidants in carcinogenesis. Blood 1990, 76, 655–663. [Google Scholar] [CrossRef] [PubMed]
- De la Fuente, M. Effects of antioxidants on immune system ageing. Eur. J. Clin. Nutr. 2002, 56, S5–S8. [Google Scholar] [CrossRef]
- Li, X.C.; Wang, T.T.; Liu, J.J.; Liu, Y.L.; Zhang, J.; Lin, J.; Zhao, Z.X.; Chen, D.F. Effect and mechanism of wedelolactone as antioxidant-coumestan on •OH-treated mesenchymal stem cells. Arab. J. Chem. 2020, 13, 184–192. [Google Scholar] [CrossRef]
- Gutteridge, J.M.; Michell, J. Redox imbalance in the critically ill. Br. Med. Bull. 1999, 55, 49–75. [Google Scholar] [CrossRef]
- De La Fuente, M.; Victor, V. Antioxidants as modulators of immune function. Immunol. Cell Biol. 2000, 78, 49–54. [Google Scholar] [CrossRef]
- Schroeder, B.O.; Birchenough, G.M.H.; Stáhlman, M.; Arike, L.; Johansson, M.E.V.; Hansson, G.C.; Bäckhed, F. Bifidobacteria or fiber protects against diet-induced microbiota-mediated colonic mucus deterioration. Cell Host Microbe 2018, 23, 27–40. [Google Scholar] [CrossRef] [PubMed]
- Tena, D.; Losa, C.; Medina, M.J.; Sáez-Nieto, J.A. Peritonitis caused by Bifidobacterium longum: Case report and literature review. Anaerobe 2014, 27, 27–30. [Google Scholar] [CrossRef] [PubMed]
- Sabrina-Duranti, S.; Lugli, G.A.; Mancabelli, L.; Turroni, F.; Milani, C.; Mangifesta, M.; Ferrario, C.; Anzalone, R.; Viappiani, A.; Sinderen, D.V.; et al. Prevalence of antibiotic resistance genes among human gut-derived bifidobacteria. Appl. Environ. Microbiol. 2017, 83, e02894-16. [Google Scholar]
- Inturri, R.; Molinaro, A.; Di Lorenzo, F.; Blandino, G.; Tomasello, B.; Hidalgo-Cantabrana, C.; De Castro, C.; Ruas-Madiedo, P. Chemical and biological properties of the novel exopolysaccharide produced by a probiotic strain of Bifidobacterium longum. Carbohydr. Polym. 2017, 174, 1172–1180. [Google Scholar] [CrossRef] [PubMed]
- Angelin, J.; Kavitha, M. Exopolysaccharides from probiotic bacteria and their health potential. Int. J. Biol. Macromol. 2020, 162, 853–865. [Google Scholar] [CrossRef]
- Inturri, R.; Mangano, K.; Santagati, M.; Intrieri, M.; Marco, R.; Blandino, G. Immunomodulatory effects of Bifidobacterium longum W11 produced exopolysaccharide on cytokine production. Curr. Pharm. Biotechnol. 2017, 18, 883–889. [Google Scholar] [CrossRef] [PubMed]
- Salazar, N.; Prieto, A.; Leal, J.A.; Mayo, B.; Bada-Gancedo, J.C.; Reyes-Gavilán, C.G.; Ruas-Madiedo, P. Production of exopolysaccharides by Lactobacillus and Bifidobacterium strains of human origin, and metabolic activity of the producing bacteria in milk. J. Dairy Sci. 2009, 92, 4158–4168. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Shen, C.; Huo, K.; Cai, D.; Zhao, G. Antioxidant activity of yeast mannans and their growth-promoting effect on Lactobacillus strains. Food Funct. 2021, 12, 10423–10431. [Google Scholar] [CrossRef]
- Zhao, Y.; Wang, J.; Fu, Q.; Zhang, H.; Liang, J.; Xue, W.; Zhao, G.; Oda, H. Characterization and Antioxidant Activity of Mannans from Saccharomyces cerevisiae with Different Molecular Weight. Molecules 2022, 27, 4439. [Google Scholar] [CrossRef]
- Salazar, N.; Ruas-Madiedo, P.; Prieto, A.; Calle, L.P.; de Los Reyes-Gavilán, C.G. Characterization of exopolysaccharides produced by Bifidobacterium longum NB667 and its cholate-resistant derivative strain IPLA B667dCo. J. Agric. Food Chem. 2012, 60, 1028–1035. [Google Scholar] [CrossRef]
- Salazar, N.; Gueimonde, M.; de Los Reyes-Gavilán, C.G.; Ruas-Madiedo, P. Exopolysaccharides Produced by Lactic Acid Bacteria and Bifidobacteria as Fermentable Substrates by the Intestinal Microbiota. Crit. Rev. Food Sci. Nutr. 2016, 56, 1440–1453. [Google Scholar] [CrossRef]
- Shang, N.; Xu, R.; Li, P. Structure characterization of an exopolysaccharide produced by Bifidobacterium animalis RH. Carbohydr. Polym. 2013, 91, 128–134. [Google Scholar] [CrossRef] [PubMed]
- Amiri, S.; Rezaei Mokarram, R.; Sowti Khiabani, M.; Rezazadeh Bari, M.; Alizadeh Khaledabad, M. Exopolysaccharides production by Lactobacillus acidophilus LA5 and Bifidobacterium animalis subsp. lactis BB12: Optimization of fermentation variables and characterization of structure and bioactivities. Int. J. Biol. Macromol. 2019, 123, 752–765. [Google Scholar] [CrossRef] [PubMed]
- Rjeibi, I.; Feriani, A.; Hentati, F.; Hfaiedh, N.; Michaud, P.; Pierre, G. Structural characterization of water-soluble polysaccharides from Nitraria retusa fruits and their antioxidant and hypolipidemic activities. Int. J. Biol. Macromol. 2019, 129, 422–432. [Google Scholar] [CrossRef] [PubMed]
- Ramamoorthy, S.; Gnanakan, A.; Lakshmana, S.S.; Meivelu, M.; Jeganathan, A. Structural characterization and anticancer activity of extracellular polysaccharides from ascidian symbiotic bacterium Bacillus thuringiensis. Carbohydr. Polym. 2018, 190, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Sran, K.S.; Bisht, B.; Mayilraj, S.; Roy-Choudhury, A. Structural characterization and antioxidant potential of a novel anionic exopolysaccharide produced by marine Microbacterium aurantiacum FSW-25. Int. J. Biol. Macromol. 2019, 131, 343–352. [Google Scholar] [CrossRef]
- Zhao, D.; Jiang, J.; Du, R.; Guo, S.; Ping, W.; Ling, H.; Ge, J. Purification and characterization of an exopolysaccharide from Leuconostoc lactis L2. Int. J. Biol. Macromol. 2019, 139, 1224–1231. [Google Scholar] [CrossRef]
- Jenny Angel, S.; Vidyadharani, G.; Santhosh, S.; Dhandapani, R. Optimization and Characterisation of Thermo Stable Exopolysaccharide Produced from Bacillus licheniformis WSF-1 Strain. J. Polym. Environ. 2018, 26, 3824–3833. [Google Scholar] [CrossRef]
- Liu, T.; Zhou, K.; Yin, S.; Liu, S.; Zhu, Y.; Yang, Y.; Wang, C. Purification and characterization of an exopolysaccharide produced by Lactobacillus plantarum HY isolated from home-made Sichuan Pickle. Int. J. Biol. Macromol. 2019, 134, 516–526. [Google Scholar] [CrossRef]
- Hu, S.M.; Zhou, J.M.; Zhou, Q.Q.; Li, P.; Xie, Y.Y.; Zhou, T.; Gu, Q. Purification, characterization and biological activities of exopolysaccharides from Lactobacillus rhamnosus ZFM231 isolated from milk. LWT-Food Sci. Technol. 2021, 147, 111561. [Google Scholar] [CrossRef]
- Wang, J.B.; Yu, L.Y.; Zeng, X.; Zheng, J.W.; Wang, B.; Pan, L. Screening of probiotics with efficient α-glucosidase inhibitory ability and study on the structure and function of its extracellular polysaccharide. Food Biosci. 2022, 45, 101452. [Google Scholar] [CrossRef]
- Saravanan, C.; Kavitake, D.; Kandasamy, S.; Devi, P.B.; Shetty, P.H. Production, partial characterization and antioxidant properties of exopolysaccharide α-d-glucan produced by Leuconostoc lactis KC117496 isolated from an idli batter. J. Food Sci. Technol. 2019, 56, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, P.K. NMR Spectroscopy in the structural elucidation of oligosaccharides and glycosides. Phytochemistry 1992, 31, 3307–3330. [Google Scholar] [CrossRef] [PubMed]
- Graham, S.T.W.; Fryhle, B.C. Organic Chemistry, 8th ed.; Chemical Industry Press: Beijing, China, 2004; p. 1117. [Google Scholar]
- Feng, F.; Zhou, Q.Q.; Yang, Y.F.; Zhao, F.K.; Du, R.P.; Han, Y.; Xiao, H.Z.; Zhou, Z.J. Characterization of highly branched dextran produced by Leuconostoc citreum B-2 from pineapple fermented product. Int. J. Biol. Macromol. 2018, 113, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Casas-Arrojo, V.; Arrojo Agudo, M.L.Á.; Cárdenas García, C.; Carrillo, P.; Pérez Manríquez, C.; Martínez-Manzanares, E.; Abdala Díaz, R.T. Antioxidant, Immunomodulatory and Potential Anticancer Capacity of Polysaccharides (Glucans) from Euglena gracilis G.A. Klebs. Pharmaceuticals 2022, 15, 1379. [Google Scholar] [CrossRef] [PubMed]
- Kwon, M.; Lee, J.; Park, S.; Kwon, O.H.; Seo, J.; Roh, S. Exopolysaccharide Isolated from Lactobacillus plantarum L-14 Has Anti-Inflammatory Effects via the Toll-Like Receptor 4 Pathway in LPS-Induced RAW 264.7 Cells. Int. J. Mol. Sci. 2020, 23, 9283. [Google Scholar] [CrossRef] [PubMed]
- Xie, S.Z.; Shang, Z.Z.; Li, Q.M.; Zha, X.Q.; Pan, L.H.; Luo, J.P. Dendrobium huoshanense polysaccharide regulates intestinal lamina propria immune response by stimulation of intestinal epithelial cells via toll-like receptor 4. Carbohydr. Polym. 2019, 222, 115028. [Google Scholar] [CrossRef]
- Peng, S.; Hou, Y.; Yao, J.; Fang, J. Activation of Nrf2-driven antioxidant enzymes by cardamonin confers neuroprotection of PC12 cells against oxidative damage. Food Funct. 2017, 8, 997–1007. [Google Scholar] [CrossRef]
- Sykiotis, G.P.; Habeos, I.G.; Samuelson, A.V.; Bohmann, D. The role of the antioxidant and longevity-promoting Nrf2 pathway in metabolic regulation. Curr. Opin. Clin. Nutr. Metab. Care 2011, 14, 41–48. [Google Scholar] [CrossRef]
- Jin, Y.X.; Miao, W.Y.; Lin, X.J.; Pan, X.H.; Ye, Y.; Xu, M.J.; Fu, Z.W. Acute exposure to 3-methylcholanthrene induces hepatic oxidative stress via activation of the Nrf2/ARE signaling pathway in mice. Environ. Toxicol. 2015, 29, 1399–1408. [Google Scholar] [CrossRef]
- Lee, D.; Hong, S.; Jung, K.; Choi, S.; Kang, K.S. Suppressive Effects of Flavonoids on Macrophage-Associated Adipocyte Inflammation in a Differentiated Murine Preadipocyte 3T3-L1 Cells Co-Cultured with a Murine Macrophage RAW264.7 Cells. Plants 2022, 11, 3552. [Google Scholar] [CrossRef] [PubMed]
- Choi, G.; Han, A.R.; Lee, J.H.; Park, J.Y.; Kang, U.; Hong, J.; Kim, Y.S.; Seo, E.K. A comparative study on hulled adlay and unhulled adlay through evaluation of their LPS-induced anti-inflammatory effects, and isolation of pure compounds. Chem. Biodivers. 2015, 12, 380–387. [Google Scholar] [CrossRef] [PubMed]
- Badowski, M.; Shultz, C.L.; Eason, Y.; Ahmad, N.; Harris, D.T. The influence of intrinsic and extrinsic factors on immune system aging. Immunobiology 2014, 219, 482–485. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.K. Anti-inflammatory Effects of Eriodictyol in Lipopolysaccharide stimulated raw 264.7 murine macrophages. Arch. Pharm. Res. 2011, 34, 671–679. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Li, X.; Chen, B.; Wei, G.; Chen, D. E-Configuration Improves Antioxidant and Cytoprotective Capacities of Resveratrols. Molecules 2018, 23, 1790. [Google Scholar] [CrossRef] [PubMed]
- Li, X. 2-Phenyl-4,4,5,5-tetramethylimidazoline-1-oxyl 3-Oxide (PTIO) radical scavenging: A new and simple antioxidant assay in vitro. J. Agric. Food. Chem. 2017, 65, 6288–6297. [Google Scholar] [CrossRef] [PubMed]
- Bergeron, F.; Auvré, F.; Radicella, J.P.; Ravanat, J.L. HO• radicals induce an unexpected high proportion of tandem base lesions refractory to repair by DNA glycosylases. Proc. Natl. Acad. Sci. USA 2010, 107, 5528–5533. [Google Scholar] [CrossRef] [PubMed]
- Kazmierczak, L.; Wolszczak, M.; Swiatla-Wojcik, D. Ionic-Equilibrium-Based Mechanism of •OH Conversion to Dichloride Radical Anion in Aqueous Acidic Solutions by Kinetic and Theoretical Studies. J. Phys. Chem. B 2019, 123, 528–533. [Google Scholar] [CrossRef]
- Lu, Q.; Liu, Y.; Li, B.; Feng, L.; Du, Z.; Zhang, L. Reaction kinetics of dissolved black carbon with hydroxyl radical, sulfate radical and reactive chlorine radicals. Sci. Total Environ. 2022, 828, 153984. [Google Scholar] [CrossRef]
- Gu, H.; Gwon, M.G.; Kim, J.H.; Leem, J.; Lee, S.J. Oridonin Attenuates Cisplatin-Induced Acute Kidney Injury via Inhibiting Oxidative Stress, Apoptosis, and Inflammation in Mice. Biomed. Res. Int. 2022, 16, 3002962. [Google Scholar] [CrossRef]
- Rauf, A.; Uddin, G.; Siddiqui, B.S.; Khan, H.; Shah, S.U.; Ben Hadda, T.; Mabkhot, Y.N.; Farooq, U.; Khan, A. Antinociceptive and anti-inflammatory activities of flavonoids isolated from Pistacia integerrima galls. Complement. Ther. Med. 2016, 25, 132–138. [Google Scholar] [CrossRef] [PubMed]
- Abdallah, H.M.; El-Agamy, D.S.; Ibrahim, S.R.M.; Mohamed, G.A.; Elsaed, W.M.; Elghamdi, A.A.; Safo, M.K.; Malebari, A.M. Euphorbiacuneata Represses LPS-induced Acute Lung Injury in Mice via its Antioxidative and Anti-inflammatory Activities. Plants 2020, 11, 1620. [Google Scholar] [CrossRef]
- Salazar, N.; López, P.; Garrido, P.; Moran, J.; Cabello, E.; Gueimonde, M.; Suárez, A.; González, C.; de los Reyes-Gavilán, C.G.; Ruas-Madiedo, P. Immune modulating capability of two exopolysaccharide-producing Bifidobacterium strains in a Wistar rat model. Biomed. Res. Int. 2014, 2014, 106290. [Google Scholar] [CrossRef] [PubMed]
- Praveen, M.A.; Parvathy, K.R.K.; Patra, S.; Khan, I.; Natarajan, P.; Balasubramanian, P. Cytotoxic and pharmacokinetic studies of Indian seaweed polysaccharides for formulating raindrop synbiotic candy. Int. J. Biol. Macromol. 2020, 154, 557–566. [Google Scholar] [CrossRef] [PubMed]
- Pandeirada, C.O.; Maricato, É.; Ferreira, S.S.; Correia, V.G.; Pinheiro, B.A.; Evtuguin, D.V.; Palma, A.S.; Correia, A.; Vilanova, M.; Coimbra, M.A.; et al. Structural analysis and potential immunostimulatory activity of Nannochloropsis oculata polysaccharides. Carbohydr. Polym. 2019, 222, 114962. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Li, X.; Chen, J.; Deng, Y.; Lu, W.; Chen, D. pH Effect and chemical mechanisms of antioxidant higenamine. Molecules 2018, 23, 2176. [Google Scholar] [CrossRef] [PubMed]
- Herzi, N.; Bouajila, J.; Camy, S.; Romdhane, M.; Condoret, J.S. Comparison of different methods for extraction from Tetraclinis articulata: Yield, chemical composition and antioxidant activity. Food Chem. 2013, 141, 3537–3545. [Google Scholar] [CrossRef]
- Kalinowska, M.; Gołębiewska, E.; Mazur, L.; Lewandowska, H.; Pruszyński, M.; Świderski, G.; Wyrwas, M.; Pawluczuk, N.; Lewandowski, W. Crystal Structure, Spectroscopic Characterization, Antioxidant and Cytotoxic Activity of New Mg(II) and Mn(II)/Na(I) Complexes of Isoferulic Acid. Materials 2021, 14, 3236. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Assays | S-EPS-1 (μg/mL) | Positive Controls | |
|---|---|---|---|
| Trolox (μg/mL) | Melatonin (μg/mL) | ||
| ABTS+• scavenging | 221.8 ± 2.5 b | 4.7 ± 0.1 b | 14.1 ± 0.4 b |
| DPPH• scavenging | 740.2 ± 14.9 b | 2.4 ± 0.1 b | 1068.1 ± 171.3 b |
| FRAP | 141.3 ± 3.8 a | 30.7 ± 2.1 a | 89.5 ± 4.9 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Gong, J.; Huang, W.; Liu, W.; Ma, C.; Liang, R.; Chen, Y.; Xie, Z.; Li, P.; Liao, Q. Structural Analysis and Antioxidant and Immunoregulatory Activities of an Exopolysaccharide Isolated from Bifidobacterium longum subsp. longum XZ01. Molecules 2023, 28, 7448. https://doi.org/10.3390/molecules28217448
Zhang X, Gong J, Huang W, Liu W, Ma C, Liang R, Chen Y, Xie Z, Li P, Liao Q. Structural Analysis and Antioxidant and Immunoregulatory Activities of an Exopolysaccharide Isolated from Bifidobacterium longum subsp. longum XZ01. Molecules. 2023; 28(21):7448. https://doi.org/10.3390/molecules28217448
Chicago/Turabian StyleZhang, Xingyuan, Jing Gong, Wenyi Huang, Wen Liu, Chong Ma, Rongyao Liang, Ye Chen, Zhiyong Xie, Pei Li, and Qiongfeng Liao. 2023. "Structural Analysis and Antioxidant and Immunoregulatory Activities of an Exopolysaccharide Isolated from Bifidobacterium longum subsp. longum XZ01" Molecules 28, no. 21: 7448. https://doi.org/10.3390/molecules28217448
APA StyleZhang, X., Gong, J., Huang, W., Liu, W., Ma, C., Liang, R., Chen, Y., Xie, Z., Li, P., & Liao, Q. (2023). Structural Analysis and Antioxidant and Immunoregulatory Activities of an Exopolysaccharide Isolated from Bifidobacterium longum subsp. longum XZ01. Molecules, 28(21), 7448. https://doi.org/10.3390/molecules28217448