
The Phenolic Profile and Anti-Inflammatory Effect of Ethanolic Extract of Polish Propolis on Activated Human Gingival Fibroblasts-1 Cell Line

 ,
,  ,
,  , ,
, ,  and
and 
Abstract
:1. Introduction
2. Results
2.1. The Phenolics Content in Ethanol Extract of Propolis
2.2. Effect of Ethanolic Extract of Propolis on Viability of HGF-1
2.3. Effect of CAPE on Viability of HGF-1
2.4. Effect of Ethanolic Extract of Propolis on Selected Pro-Inflammatory Cytokines Production in Fibroblast HGF-1 Cells Stimulated by LPS, IFN-α, LPS + IFN-α
2.5. Effect of CAPE on Selected Pro-Inflammatory Cytokines Production in Fibroblast HGF-1 Cells Stimulated by LPS, IFN-α, and LPS + IFN-α
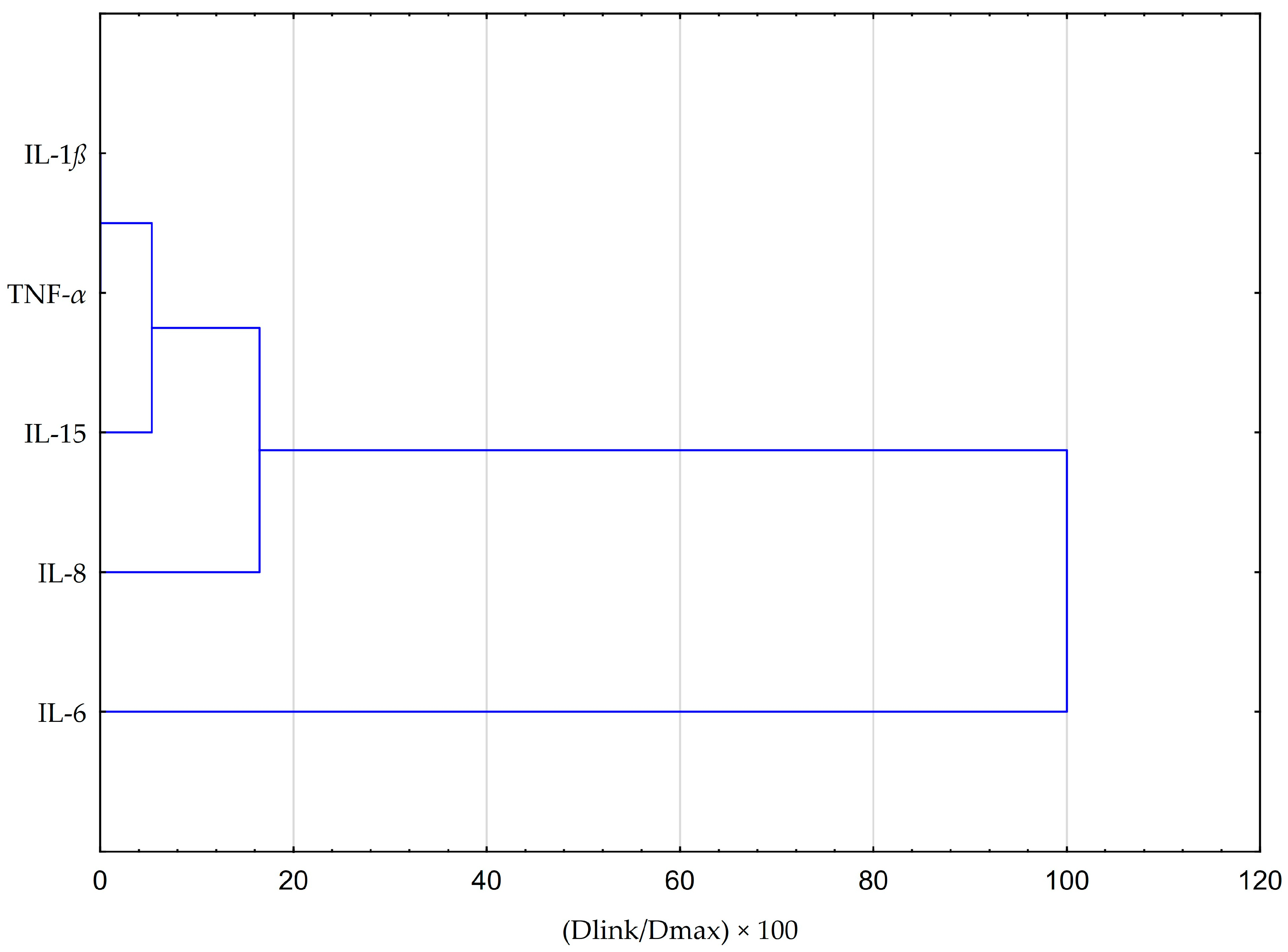
2.6. Statistical Analysis Comparing the Impact of EEP and CAPE on Cytokines Production in Fibroblasts HGF-1 Cells Stimulated by LPS, IFN-α, and LPS + IFN-α
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Ethanol Extract of Propolis Formulation
4.3. RP-HPLC-PDA Analysis
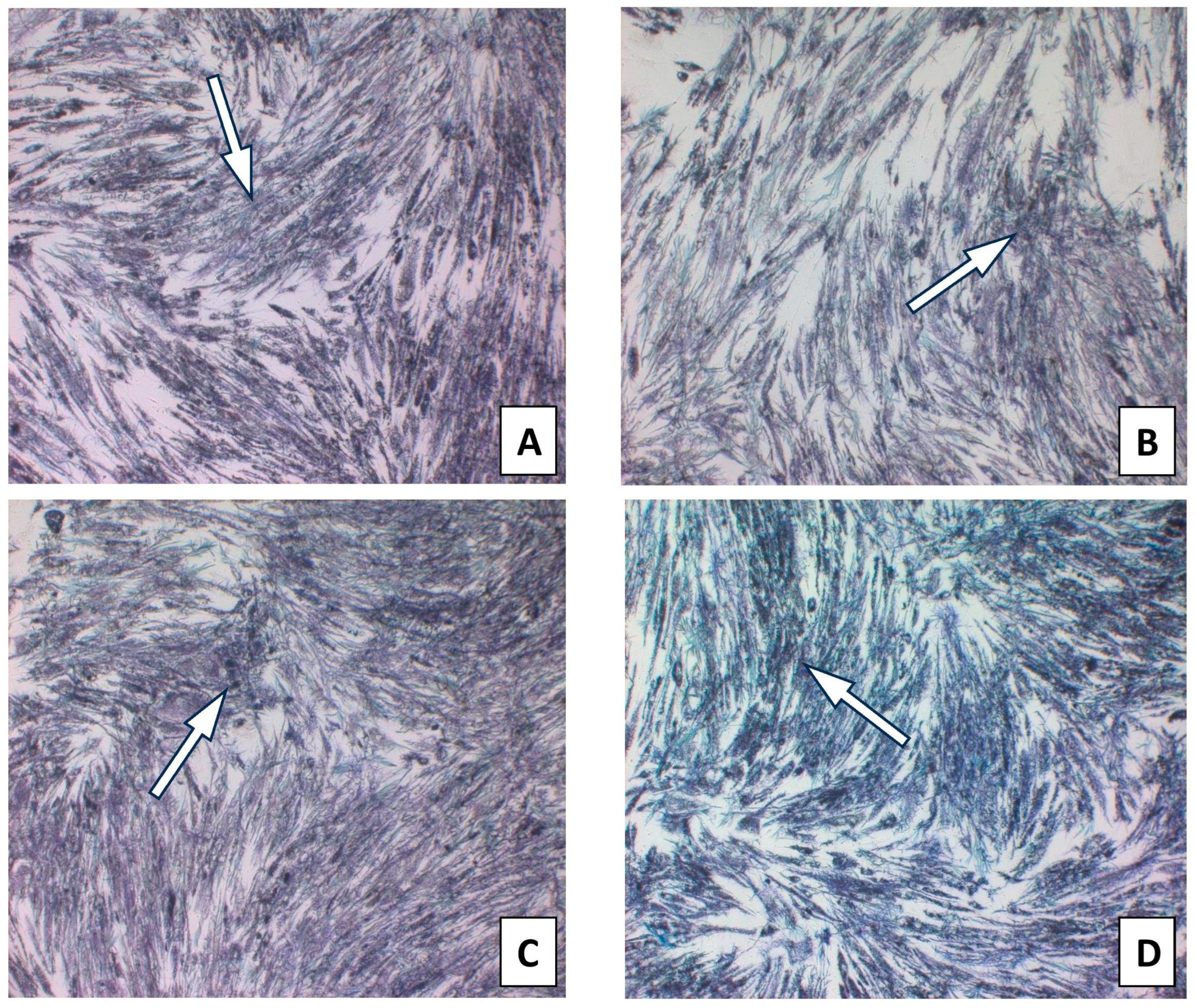
4.4. HGF-1 Collection
4.5. Cell Viability Assay
4.6. Multiplex Bead-Based Cytokine Assay
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kurek-Górecka, A.; Walczyńska-Dragon, K.; Felitti, R.; Nitecka-Buchta, A.; Baron, S.; Olczyk, P. The Influence of Propolis on Dental Plaque Reduction and the Correlation between Dental Plaque and Severity of COVID-19 Complications—A Literature Review. Molecules 2021, 26, 5516. [Google Scholar] [CrossRef]
- Liccardo, D.; Cannavo, A.; Spagnuolo, G.; Ferrara, N.; Cittadini, A.; Rengo, C.; Rengo, G. Periodontal Disease: A Risk Factor for Diabetes and Cardiovascular Disease. Int. J. Mol. Sci. 2019, 20, 1414. [Google Scholar] [CrossRef]
- Ceccarelli, F.; Saccucci, M.; Di Carlo, G.; Lucchetti, R.; Pilloni, A.; Pranno, N.; Luzzi, V.; Valesini, G.; Polimeni, A. Periodontitis and Rheumatoid Arthritis: The Same Inflammatory Mediators? Mediat. Inflamm. 2019, 2019, 6034546. [Google Scholar] [CrossRef]
- Sansores-España, D.; Carrillo-Avila, A.; Melgar-Rodriguez, S.; Díaz-Zuñiga, J.; Martínez-Aguilar, V. Periodontitis and alzheimer’s disease. Med. Oral. Patol. Oral. Cir. Bucal. 2021, 26, e43–e48. [Google Scholar] [CrossRef] [PubMed]
- Gündogar, H.; Üstün, K.; Şenyurt, S.Z.; Özdemir, E.Ç.; Sezer, U.; Erciyas, K. Gingival crevicular fluid levels of cytokine, chemokine, and growth factors in patients with periodontitis or gingivitis and periodontally healthy subjects: A cross-sectional multiplex study. Central Eur. J. Immunol. 2021, 46, 474–480. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.M.; An, J. Cytokines, Inflammation, and Pain. Int. Anesthesiol. Clin. 2007, 45, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Dye, B.A.; Choudhary, K.; Shea, S.; Papapanou, P.N. Serum Antibodies to Periodontal Pathogens and Markers of Systemic Inflammation. J. Clin. Periodontol. 2005, 32, 1189–1199. [Google Scholar] [CrossRef] [PubMed]
- Kurek-Górecka, A.; Walczyńska-Dragon, K.; Felitti, R.; Baron, S.; Olczyk, P. Propolis and Diet Rich in Polyphenols as Cariostatic Agents Reducing Accumulation of Dental Plaque. Molecules 2022, 27, 271. [Google Scholar] [CrossRef]
- Benakanakere, M.R.; Finoti, L.S.; Tanaka, U.; Grant, G.R.; Scarel-Caminaga, R.M.; Kinane, D.F. Investigation of the Functional Role of Human Interleukin-8 Gene Haplotypes by CRISPR/Cas9 Mediated Genome Editing. Sci. Rep. 2016, 6, 31180. [Google Scholar] [CrossRef]
- Palomo, J.; Dietrich, D.; Martin, P.; Palmer, G.; Gabay, C. The Interleukin (IL)-1 Cytokine Family–Balance between Agonists and Antagonists in Inflammatory Diseases. Cytokine 2015, 76, 25–37. [Google Scholar] [CrossRef]
- Lopez-Castejon, G.; Brough, D. Understanding the Mechanism of IL-1β Secretion. Cytokine Growth Factor Rev. 2011, 22, 189–195. [Google Scholar] [CrossRef]
- Paludan, S.R.; Pradeu, T.; Masters, S.L.; Mogensen, T.H. Constitutive Immune Mechanisms: Mediators of Host Defence and Immune Regulation. Nat. Rev. Immunol. 2021, 21, 137–150. [Google Scholar] [CrossRef]
- Popko, K.; Gorska, E.; Stelmaszczyk-Emmel, A.; Plywaczewski, R.; Stoklosa, A.; Gorecka, D.; Pyrzak, B.; Demkow, U. Proinflammatory Cytokines IL-6 and TNF-α and the Development of Inflammation in Obese Subjects. Eur. J. Med. Res. 2010, 15 (Suppl. 2), 120–122. [Google Scholar] [CrossRef] [PubMed]
- Parihar, A.S.; Jain, P.; Ved, A.; Dubey, R.; Singh, N.; Maytreyee, R. Comparative Evaluation of Serum Tumor Necrosis Factor α in Health and Chronic Periodontitis: A Case–Control Study. Contemp. Clin. Dent. 2020, 11, 342–349. [Google Scholar] [CrossRef]
- Sun, W.; Xie, W.; Huang, D.; Cui, Y.; Yue, J.; He, Q.; Jiang, L.; Xiong, J.; Sun, W.; Yi, Q. Caffeic acid phenethyl ester attenuates osteoarthritis progression by activating NRF2/HO-1 and inhibiting the NF-κB signaling pathway. Int. J. Mol. Med. 2022, 50, 134. [Google Scholar] [CrossRef] [PubMed]
- Segueni, N.; Boutaghane, N.; Asma, S.T.; Tas, N.; Acaroz, U.; Arslan-Acaroz, D.; Shah, S.R.A.; Abdellatieff, H.A.; Akkal, S.; Peñalver, R.; et al. Review on Propolis Applications in Food Preservation and Active Packaging. Plants 2023, 12, 1654. [Google Scholar] [CrossRef] [PubMed]
- Naeem, M.M.; Sarwar, H.; Nisar, A.; Ahmed, S.; Shabbir, J.; Khurshid, Z.; Palma, P.J. Effect of Propolis on Root Dentine Microhardness When Used as an Intracanal Medicament: An In Vitro Study. J. Funct. Biomater. 2023, 14, 144. [Google Scholar] [CrossRef]
- Całkosiński, I.; Dobrzyński, M.; Całkosińska, M.; Seweryn, E.; Bronowicka-Szydełko, A.; Dzierzba, K.; Ceremuga, I.; Gamian, A. Characterization of an Inflammatory Response. Postep. Hig. Med. Dosw. 2009, 63, 395–408. [Google Scholar]
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory responses and inflammation-associated diseases in organs. Oncotarget 2017, 9, 7204–7218. [Google Scholar] [CrossRef]
- Zamarrenho, L.G.; de Lima, M.H.F.; Hori, J.I.; Lima, J.A.; Ambrósio, S.R.; Bastos, J.K.; De Jong, D.; Berretta, A.A. Effects of Three Different Brazilian Green Propolis Extract Formulations on Pro- and Anti-Inflammatory Cytokine Secretion by Macrophages. Appl. Sci. 2023, 13, 6247. [Google Scholar] [CrossRef]
- Abdulkhaleq, L.A.; Assi, M.A.; Abdullah, R.; Zamri-Saad, M.; Taufiq-Yap, Y.H.; Hezmee, M.N.M. The Crucial Roles of Inflammatory Mediators in Inflammation: A review. Vet. World 2018, 11, 627–635. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Xian, X.; Wang, Z.; Bi, Y.; Chen, Q.; Han, X.; Tang, D.; Chen, R. Research Progress on the Relationship between Atherosclerosis and Inflammation. Biomolecules 2018, 8, 80. [Google Scholar] [CrossRef] [PubMed]
- Scheller, J.; Chalaris, A.; Schmidt-Arras, D.; Rose-John, S. The Pro- and Anti-Inflammatory Properties of the Cytokine Interleukin-6. Biochim. Biophys. Acta (BBA) Mol. Cell Res. 2011, 1813, 878–888. [Google Scholar] [CrossRef] [PubMed]
- Dağ, A.; Fırat, E.T.; Kadiroğlu, A.K.; Kale, E.; Yılmaz, M.E. Significance of Elevated Gingival Crevicular Fluid Tumor Necrosis Factor-α and Interleukin-8 Levels in Chronic Hemodialysis Patients with Periodontal Disease. J. Periodontal Res. 2010, 45, 445–450. [Google Scholar] [CrossRef]
- Andia, D.C.; de Oliveira, N.F.P.; Letra, A.M.; Nociti, F.H.; Line, S.R.P.; de Souza, A.P. Interleukin-8 Gene Promoter Polymorphism (rs4073) May Contribute to Chronic Periodontitis. J. Periodontol. 2011, 82, 893–899. [Google Scholar] [CrossRef]
- Perera, P.-Y.; Lichy, J.H.; Waldmann, T.A.; Perera, L.P. The Role of Interleukin-15 in Inflammation and Immune Responses to Infection: Implications for Its Therapeutic Use. Microbes Infect. 2012, 14, 247–261. [Google Scholar] [CrossRef] [PubMed]
- Kany, S.; Vollrath, J.T.; Relja, B. Cytokines in Inflammatory Disease. Int. J. Mol. Sci. 2019, 20, 6008. [Google Scholar] [CrossRef]
- Jang, D.-I.; Lee, A.-H.; Shin, H.-Y.; Song, H.-R.; Park, J.-H.; Kang, T.-B.; Lee, S.-R.; Yang, S.-H. The Role of Tumor Necrosis Factor Alpha (TNF-α) in Autoimmune Disease and Current TNF-α Inhibitors in Therapeutics. Int. J. Mol. Sci. 2021, 22, 2719. [Google Scholar] [CrossRef]
- Boyce, B.E.; Li, P.; Yao, Z.; Zhang, Q.; Badell, I.R.; Schwarz, E.M.; O’keefe, R.J.; Xing, L. TNF-Alpha and Pathologic Bone Resorption. Keio J. Med. 2005, 54, 127–131. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.; Gupta, N.D.; Bey, A.; Khan, S. Salivary TNF-alpha: A Potential Marker of Periodontal Destruction. J. Indian Soc. Periodontol. 2014, 18, 306–310. [Google Scholar] [CrossRef]
- Can, Z.; Yildiz, O.; Sahin, H.; Turumtay, E.A.; Silici, S.; Kolayli, S. An Investigation of Turkish Honeys: Their Physico-Chemical Properties, Antioxidant Capacities and Phenolic Profiles. Food Chem. 2015, 180, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Kara, Y.; Can, Z.; Kolaylı, S. Applicability of Phenolic Profile Analysis Method Developed with RP-HPLC-PDA to some Bee Product. Braz. Arch. Biol. Technol. 2022, 65, 2022. [Google Scholar] [CrossRef]
- Kłósek, M.; Sędek, L.; Lewandowska, H.; Czuba, Z.P. The effect of ethanolic extract of Brazilian green propolis and artepillin C on aFGF-1, Eselectin, and CD40L secreted by human gingival fibroblasts. Central Eur. J. Immunol. 2021, 46, 438–445. [Google Scholar] [CrossRef] [PubMed]
- Bronikowska, J.; Kłósek, M.; Janeczko, T.; Kostrzewa-Susłow, E.; Czuba, Z.P. The modulating effect of methoxy-derivatives of 2’-hydroxychalcones on the release of IL-8, MIF, VCAM-1 and ICAM-1 by colon cancer cells. Biomed. Pharmacother. 2022, 145, 112428. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | Phenolic Compounds | Content [µg Phenolic/g EEP] | p |
|---|---|---|---|
| 1. | gallic acid | 12.789 ± 0.170 | 0.000 |
| 2. | protocatechuic acid | - | - |
| 3. | chlorogenic acid | 11.022 ± 0.02 | 0.000 |
| 4. | p-OH Benzoic acid | - | - |
| 5. | epicatechin | - | - |
| 6. | caffeic acid | 320.344 ± 1.14 | 0.001 |
| 7. | syringic acid | - | - |
| 8. | m-OH benzoic acid | - | - |
| 9. | routine | - | - |
| 10. | ellagic acid | - | - |
| 11. | p-coumaric acid | 3002.604 ± 32.06 | 0.009 |
| 12. | ferulic acid | 318.816 ± 18.01 | 0.002 |
| 13. | myricetin | - | - |
| 14. | resveratrol | - | - |
| 15. | daidzein | - | - |
| 16. | luteolin | 8.799 ± 0.08 | 0.000 |
| 17. | quercetin | 90.700 ± 2.1 | 0.000 |
| 18. | t-cinnamic acid | 59.210 ± 1.06 | 0.002 |
| 19. | apigenin | 328.005 ± 12.3 | 0.003 |
| 20. | hesperitin | - | - |
| 21. | rhamnetin | - | - |
| 22. | chrysin | 1998.700 ± 56.4 | 0.005 |
| 23. | pinocembrin | 7.899 ± 0.7 | 0.000 |
| 24. | CAPE | 989.699 ± 1.82 | 0.000 |
| 25. | curcumin | - | - |
| Sample | IL-1β | IL-6 | IL-8 | IL-15 | TNF-α | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| AVG | SD | p | AVG | SD | p | AVG | SD | p | AVG | SD | p | AVG | SD | p | |
| Control baseline (EEP) | 1.224 | 3.410 | 1.360 | 1256.310 | 4.070 | ||||||||||
| Control DMSO (EI) | 3.261 | 1.778 | 109.283 | 8.940 | 27.600 | 6.946 | 1482.840 | 156.863 | 11.330 | 7.335 | |||||
| Control LPS (ELI) | 1.315 | 0.079 | 131.800 | 23.512 | 41.693 | 4.151 | 1439.930 | 373.413 | 3.642 | 0.214 | |||||
| Control IFN-α (EI) | 2.153 | 1.374 | 105.797 | 21.038 | 24.810 | 5.595 | 1730.793 | 161.382 | 7.222 | 6.018 | |||||
| Control LPS + IFN-α (ELI) | 1.156 | 0.068 | 139.400 | 27.655 | 27.477 | 4.218 | 1534.390 | 32.684 | 3.427 | 0.742 | |||||
| EEP 25 µg/mL | 3.603 | 2.178 | 87.747 | 33.217 | 42.423 | 1.686 | 1505.647 | 44.126 | 7.294 | 5.959 | |||||
| EEP 50 µg/mL | 1.315 | 0.079 | 114.503 | 25.121 | 40.233 | 6.905 | 1539.677 | 248.184 | 3.642 | 0.214 | |||||
| EEP 25 µg/mL + LPS | 1.224 | 0.000 | 0.9442 | 85.727 | 35.643 | 0.9941 | 48.643 | 13.891 | 0.9889 | 1450.367 | 116.122 | 0.9178 | 3.856 | 0.214 | 0.9736 |
| EEP 50 µg/mL + LPS | 1.088 | 0.118 | 0.8611 | 143.770 | 67.882 | 0.9985 | 87.427 | 53.326 | 0.9271 | 1470.890 | 87.826 | 0.7596 | 5.251 | 2.416 | 0.8040 |
| EEP 25 µg/mL + IFN-α | 1.315 | 0.079 | 0.5182 | 392.797 | 35.264 | 0.9633 | 608.307 | 204.011 | 0.2460 | 1711.937 | 53.431 | 0.8521 | 4.965 | 2.663 | 0.7277 |
| EEP 50 µg/mL + IFN-α | 1.269 | 0.079 | 0.4960 | 371.620 | 27.279 | 0.9660 | 422.833 | 164.744 | 0.4271 | 1386.310 | 86.914 | 0.0012 | 3.356 | 0.539 | 0.5514 |
| EEP 25 µg/mL + LPS + IFN-α | 1.156 | 0.118 | 1.0000 | 408.203 | 35.695 | 0.9656 | 870.680 | 139.463 | 0.0959 | 1483.853 | 134.711 | 0.6177 | 7.080 | 6.164 | 0.5736 |
| EEP 50 µg/mL + LPS + IFN-α | 2.811 | 1.610 | 0.2050 | 449.213 | 54.085 | 0.9604 | 1209.220 | 246.153 | 0.0212 | 1581.167 | 42.413 | 0.6440 | 10.259 | 8.171 | 0.2943 |
| Sample | IL-1β | IL-6 | IL-8 | IL-15 | TNF-α | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| AVG | SD | p | AVG | SD | p | AVG | SD | p | AVG | SD | p | AVG | SD | p | |
| Control baseline (CAPE) | 2.440 | 7.020 | 1.420 | 97.040 | 7.779 | ||||||||||
| Control DMSO (CI) | 1.988 | 0.157 | 315.380 | 99.899 | 37.013 | 10.225 | 150.359 | 5.142 | 6.741 | 1.372 | |||||
| Control LPS (CLI) | 4.616 | 4.247 | 64,261.308 | 28,492.806 | 6603.190 | 1208.399 | 230.669 | 148.971 | 23.070 | 12.780 | |||||
| Control IFN-α (CI) | 2.079 | 0.157 | 446.373 | 41.877 | 17.213 | 6.016 | 211.100 | 94.533 | 5.877 | 0.599 | |||||
| Control LPS + IFN-α (CLI) | 5.016 | 2.708 | 91,701.724 | 26,094.008 | 6392.827 | 913.894 | 177.278 | 39.430 | 43.047 | 22.899 | |||||
| CAPE 25 µg/mL | 2.079 | 0.157 | 310.367 | 25.022 | 40.087 | 1.184 | 140.821 | 7.954 | 6.223 | 1.037 | |||||
| CAPE 50 µg/mL | 1.717 | 0.313 | 338.890 | 33.819 | 56.207 | 10.582 | 152.042 | 8.637 | 6.050 | 0.792 | |||||
| CAPE 25 µg/mL + LPS | 2.350 | 0.157 | 0.0846 | 6551.583 | 418.252 | 0.0000 | 6002.223 | 1171.625 | 0.2323 | 223.604 | 120.550 | 0.9443 | 12.063 | 4.726 | 0.0938 |
| CAPE 50 µg/mL + LPS | 2.395 | 0.078 | 0.0909 | 6175.093 | 940.004 | 0.0000 | 7580.387 | 1974.254 | 0.0548 | 153.725 | 3.887 | 0.4480 | 16.351 | 13.503 | 0.3022 |
| CAPE 25 µg/mL + IFN-α | 1.988 | 0.157 | 0.5182 | 582.057 | 72.812 | 0.9827 | 28.070 | 11.323 | 0.9827 | 135.772 | 8.303 | 0.4575 | 7.260 | 1.037 | 0.8310 |
| CAPE 50 µg/mL + IFN-α | 1.807 | 0.564 | 0.4960 | 565.647 | 36.875 | 0.9848 | 35.733 | 14.639 | 0.9704 | 137.455 | 16.173 | 0.4676 | 6.914 | 0.599 | 0.8729 |
| CAPE 25 µg/mL + LPS + IFN-α | 4.056 | 3.037 | 0.4598 | 11,247.297 | 2229.940 | 0.0000 | 6558.080 | 384.514 | 0.7410 | 150.359 | 5.142 | 0.7902 | 23.873 | 17.071 | 0.0045 |
| CAPE 50 µg/mL + LPS + IFN-α | 3.876 | 3.194 | 0.3803 | 14,107.183 | 2445.806 | 0.0000 | 8240.420 | 1333.419 | 0.0005 | 344.700 | 85.699 | 0.1022 | 24.301 | 13.609 | 0.0054 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kurek-Górecka, A.; Kłósek, M.; Pietsz, G.; Czuba, Z.P.; Kolayli, S.; Can, Z.; Balwierz, R.; Olczyk, P. The Phenolic Profile and Anti-Inflammatory Effect of Ethanolic Extract of Polish Propolis on Activated Human Gingival Fibroblasts-1 Cell Line. Molecules 2023, 28, 7477. https://doi.org/10.3390/molecules28227477
Kurek-Górecka A, Kłósek M, Pietsz G, Czuba ZP, Kolayli S, Can Z, Balwierz R, Olczyk P. The Phenolic Profile and Anti-Inflammatory Effect of Ethanolic Extract of Polish Propolis on Activated Human Gingival Fibroblasts-1 Cell Line. Molecules. 2023; 28(22):7477. https://doi.org/10.3390/molecules28227477
Chicago/Turabian StyleKurek-Górecka, Anna, Małgorzata Kłósek, Grażyna Pietsz, Zenon P. Czuba, Sevgi Kolayli, Zehra Can, Radosław Balwierz, and Paweł Olczyk. 2023. "The Phenolic Profile and Anti-Inflammatory Effect of Ethanolic Extract of Polish Propolis on Activated Human Gingival Fibroblasts-1 Cell Line" Molecules 28, no. 22: 7477. https://doi.org/10.3390/molecules28227477
APA StyleKurek-Górecka, A., Kłósek, M., Pietsz, G., Czuba, Z. P., Kolayli, S., Can, Z., Balwierz, R., & Olczyk, P. (2023). The Phenolic Profile and Anti-Inflammatory Effect of Ethanolic Extract of Polish Propolis on Activated Human Gingival Fibroblasts-1 Cell Line. Molecules, 28(22), 7477. https://doi.org/10.3390/molecules28227477