
The Construction of the Self-Induced Sal System and Its Application in Salicylic Acid Production


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
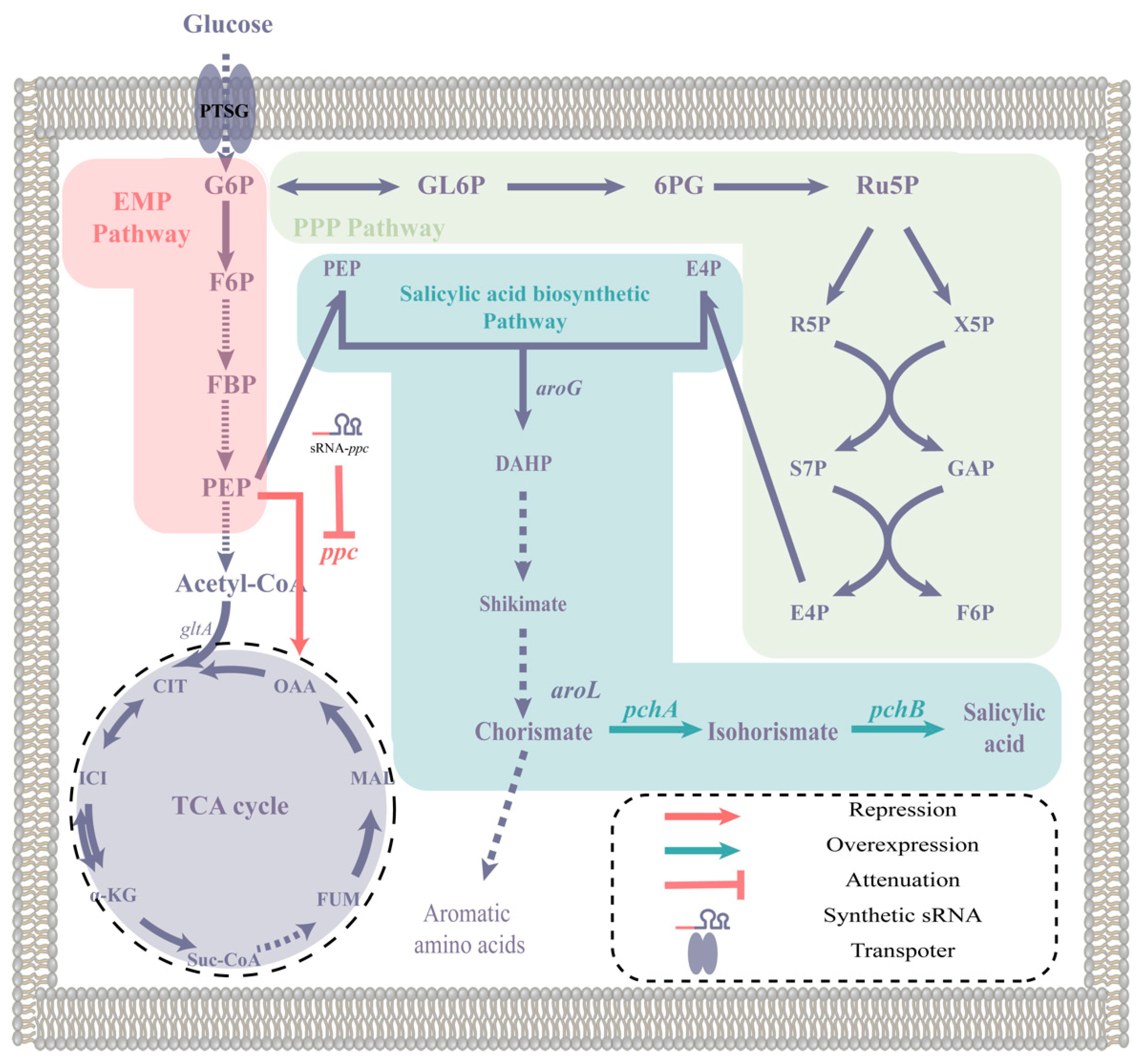
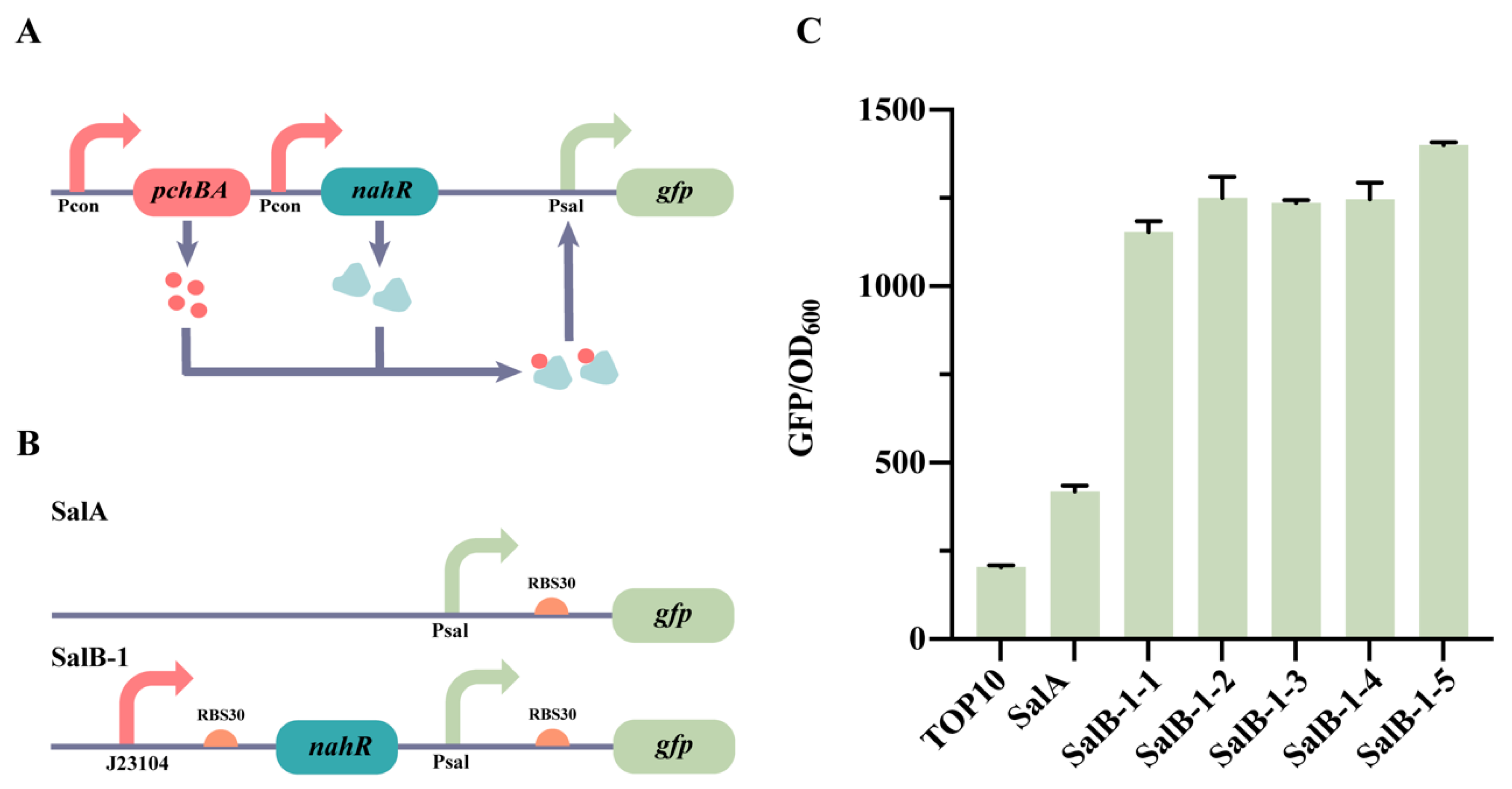
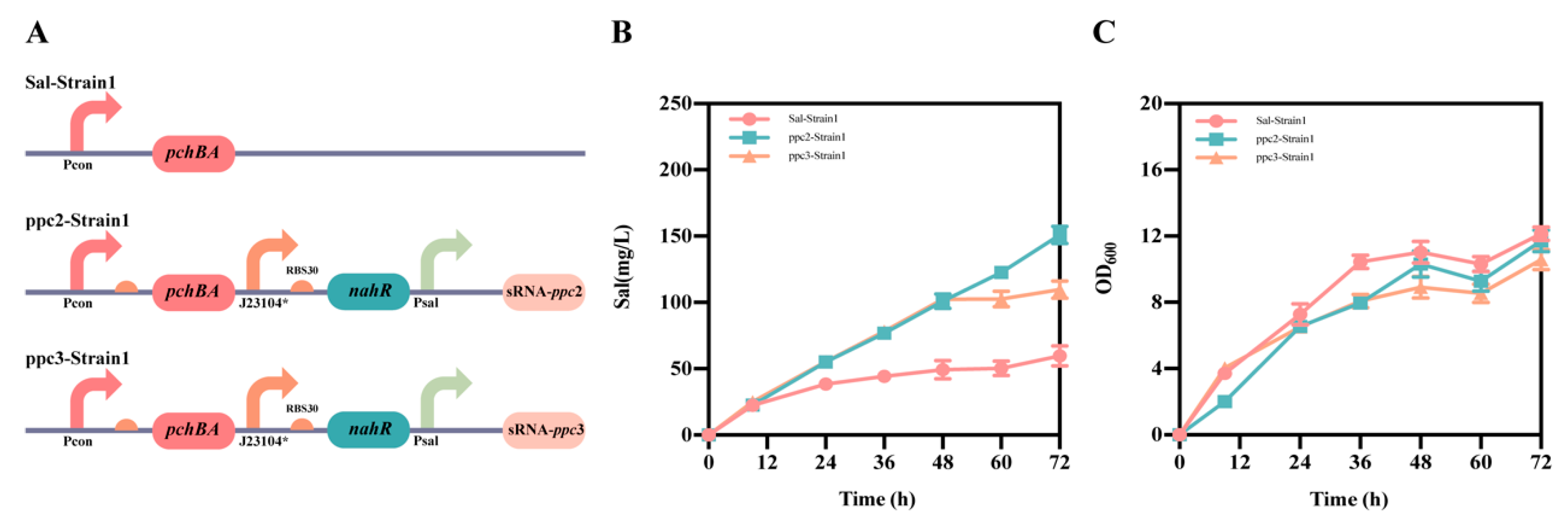
2.1. Design and Construction of the Self-Induced Sal System
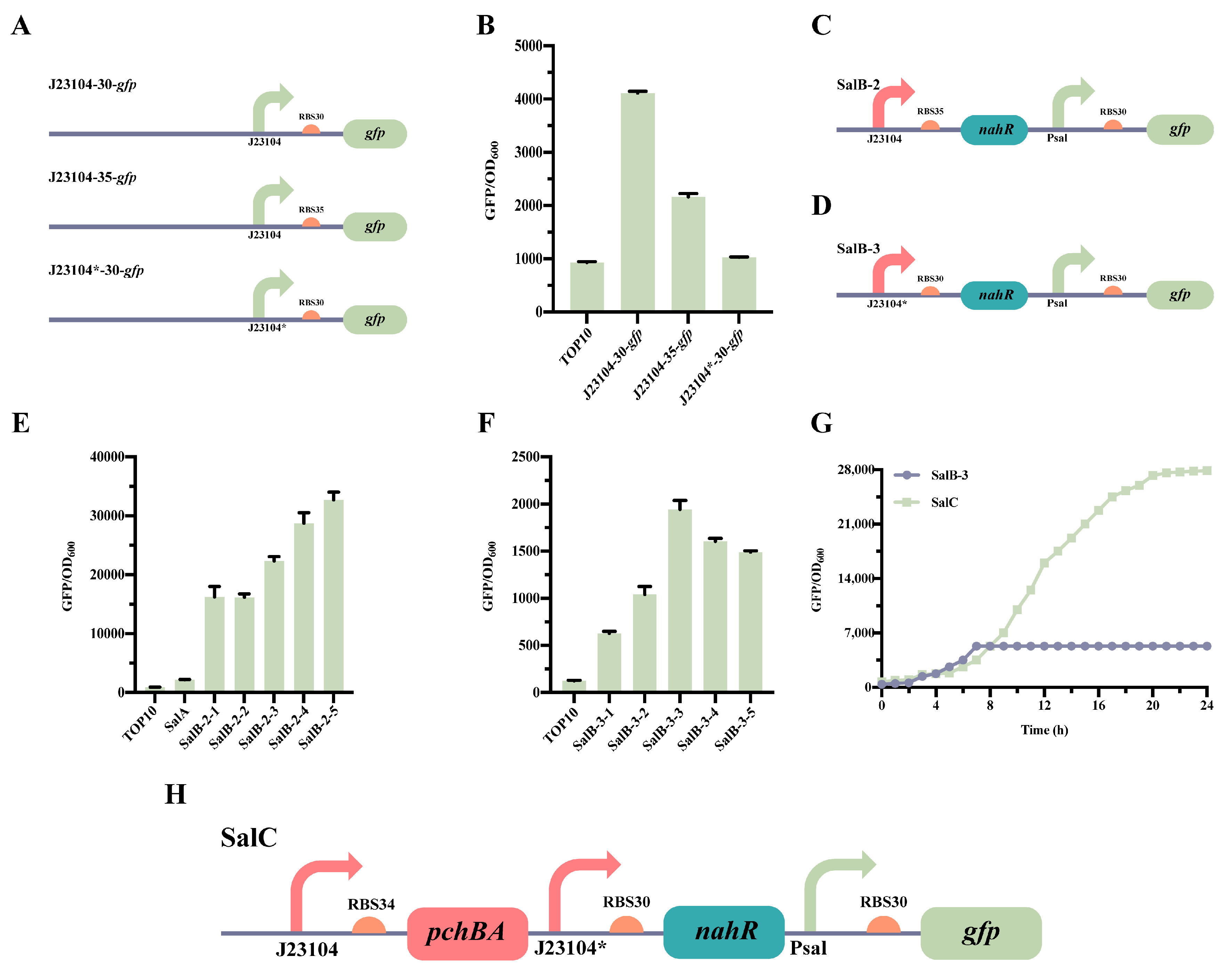
2.2. Optimization of the Self-Induced Sal System
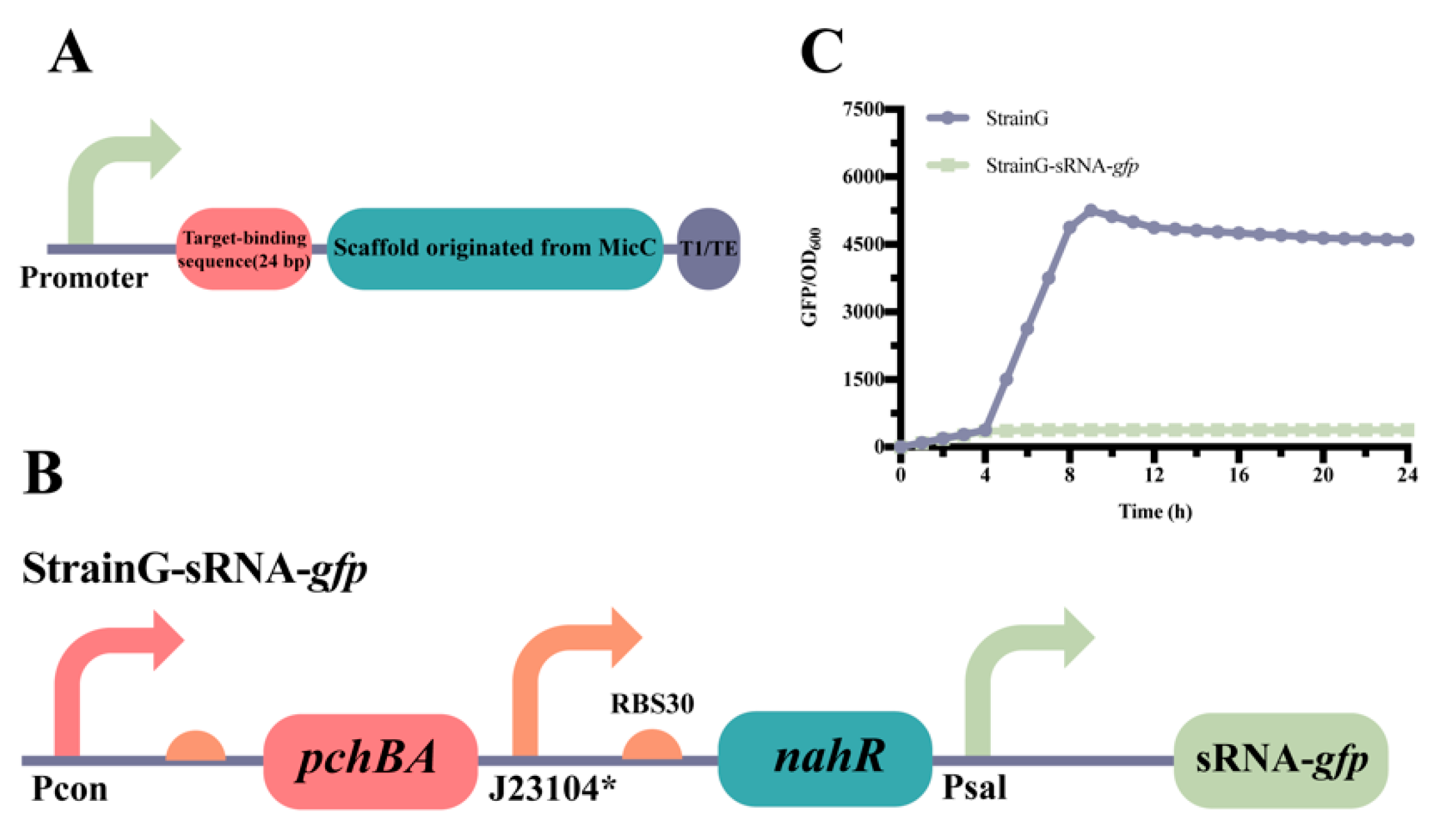
2.3. Construction of the atsRNAs That Combine with the Self-Induced Sal System
2.4. Application of the Sal System for the Production of Salicylic Acid
3. Conclusions
4. Materials and Methods
4.1. Culture Media and Salicylic Acid Fermentation
4.2. Strains and Plasmid Construction
4.3. PCR: Amplifying Files
4.4. Sal System Characterization
4.5. Analytical Methods
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gray, K.M. Intercellular communication and group behavior in bacteria. Trends Microbiol. 1997, 5, 184–188. [Google Scholar] [CrossRef] [PubMed]
- Fuqua, W.C.; Winans, S.C.; Greenberg, E.P. Quorum sensing in bacteria: The LuxR-LuxI family of cell density-responsive transcriptional regulators. J. Bacteriol. 1994, 176, 269–275. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.; Reizman, I.M.; Reisch, C.R.; Prather, K.L. Dynamic regulation of metabolic flux in engineered bacteria using a pathway-independent quorum-sensing circuit. Nat. Biotechnol. 2017, 35, 273–279. [Google Scholar] [CrossRef]
- Hu, B.; Du, J.; Zou, R.Y.; Yuan, Y.J. An environment-sensitive synthetic microbial ecosystem. PLoS ONE 2010, 5, e10619. [Google Scholar] [CrossRef] [PubMed]
- Celik Ozgen, V.; Kong, W.; Blanchard, A.E.; Liu, F.; Lu, T. Spatial interference scale as a determinant of microbial range expansion. Sci. Adv. 2018, 4, eaau0695. [Google Scholar] [CrossRef]
- Balagadde, F.K.; Song, H.; Ozaki, J.; Collins, C.H.; Barnet, M.; Arnold, F.H.; Quake, S.R.; You, L. A synthetic Escherichia coli predator-prey ecosystem. Mol. Syst. Biol. 2008, 4, 187. [Google Scholar] [CrossRef]
- Saeidi, N.; Wong, C.K.; Lo, T.M.; Nguyen, H.X.; Ling, H.; Leong, S.S.; Poh, C.L.; Chang, M.W. Engineering microbes to sense and eradicate Pseudomonas aeruginosa, a human pathogen. Mol. Syst. Biol. 2011, 7, 521. [Google Scholar] [CrossRef]
- Gu, F.; Jiang, W.; Mu, Y.; Huang, H.; Su, T.; Luo, Y.; Liang, Q.; Qi, Q. Quorum Sensing-Based Dual-Function Switch and Its Application in Solving Two Key Metabolic Engineering Problems. ACS Synth. Biol. 2020, 9, 209–217. [Google Scholar] [CrossRef]
- He, X.; Chen, Y.; Liang, Q.; Qi, Q. Autoinduced AND Gate Controls Metabolic Pathway Dynamically in Response to Microbial Communities and Cell Physiological State. ACS Synth. Biol. 2017, 6, 463–470. [Google Scholar] [CrossRef] [PubMed]
- Xu, P. Production of chemicals using dynamic control of metabolic fluxes. Curr. Opin. Biotechnol. 2018, 53, 12–19. [Google Scholar] [CrossRef]
- Papenfort, K.; Bassler, B.L. Quorum sensing signal-response systems in Gram-negative bacteria. Nat. Rev. Microbiol. 2016, 14, 576–588. [Google Scholar] [CrossRef] [PubMed]
- Hanzelka, B.L.; Greenberg, E.P. Quorum sensing in Vibrio fischeri: Evidence that S-adenosylmethionine is the amino acid substrate for autoinducer synthesis. J. Bacteriol. 1996, 178, 5291–5294. [Google Scholar] [CrossRef]
- Val, D.L.; Cronan, J.E., Jr. In vivo evidence that S-adenosylmethionine and fatty acid synthesis intermediates are the substrates for the LuxI family of autoinducer synthases. J. Bacteriol. 1998, 180, 2644–2651. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.; Wang, J.H.; Swatton, J.E.; Davenport, P.; Price, B.; Mikkelsen, H.; Stickland, H.; Nishikawa, K.; Gardiol, N.; Spring, D.R.; et al. Variations on a theme: Diverse N-acyl homoserine lactone-mediated quorum sensing mechanisms in gram-negative bacteria. Sci. Prog. 2006, 89 Pt 3–4, 167–211. [Google Scholar] [CrossRef] [PubMed]
- Scott, S.R.; Hasty, J. Quorum Sensing Communication Modules for Microbial Consortia. ACS Synth. Biol. 2016, 5, 969–977. [Google Scholar] [CrossRef]
- Daer, R.; Barrett, C.M.; Melendez, E.L.; Wu, J.; Tekel, S.J.; Xu, J.; Dennison, B.; Muller, R.; Haynes, K.A. Characterization of diverse homoserine lactone synthases in Escherichia coli. PLoS ONE 2018, 13, e0202294. [Google Scholar] [CrossRef]
- Xia, P.F.; Ling, H.; Foo, J.L.; Chang, M.W. Synthetic genetic circuits for programmable biological functionalities. Biotechnol. Adv. 2019, 37, 107393. [Google Scholar] [CrossRef]
- Brophy, J.A.; Voigt, C.A. Principles of genetic circuit design. Nat. Methods 2014, 11, 508–520. [Google Scholar] [CrossRef]
- Du, P.; Zhao, H.; Zhang, H.; Wang, R.; Huang, J.; Tian, Y.; Luo, X.; Luo, X.; Wang, M.; Xiang, Y.; et al. De novo design of an intercellular signaling toolbox for multi-channel cell-cell communication and biological computation. Nat. Commun. 2020, 11, 4226. [Google Scholar] [CrossRef]
- Jiang, W.; He, X.; Luo, Y.; Mu, Y.; Gu, F.; Liang, Q.; Qi, Q. Two Completely Orthogonal Quorum Sensing Systems with Self-Produced Autoinducers Enable Automatic Delayed Cascade Control. ACS Synth. Biol. 2020, 9, 2588–2599. [Google Scholar] [CrossRef]
- Qian, S.; Li, Y.; Cirino, P.C. Biosensor-guided improvements in salicylate production by recombinant Escherichia coli. Microb. Cell Fact. 2019, 18, 18. [Google Scholar] [CrossRef]
- Liu, L.; Li, W.; Li, X.; Sun, X.; Yuan, Q. Constructing an efficient salicylate biosynthesis platform by Escherichia coli chromosome integration. J. Biotechnol. 2019, 298, 5–10. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Sun, X.; Yuan, Q.; Yan, Y. Extending shikimate pathway for the production of muconic acid and its precursor salicylic acid in Escherichia coli. Metab. Eng. 2014, 23, 62–69. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Shen, X.; Yuan, Q.; Yan, Y. Microbial biosynthesis of the anticoagulant precursor 4-hydroxycoumarin. Nat. Commun. 2013, 4, 2603. [Google Scholar] [CrossRef] [PubMed]
- Gaille, C.; Kast, P.; Haas, D. Salicylate biosynthesis in Pseudomonas aeruginosa. Purification and characterization of PchB, a novel bifunctional enzyme displaying isochorismate pyruvate-lyase and chorismate mutase activities. J. Biol. Chem. 2002, 277, 21768–21775. [Google Scholar] [CrossRef] [PubMed]
- Xue, H.; Shi, H.; Yu, Z.; He, S.; Liu, S.; Hou, Y.; Pan, X.; Wang, H.; Zheng, P.; Cui, C.; et al. Design, construction, and characterization of a set of biosensors for aromatic compounds. ACS Synth. Biol. 2014, 3, 1011–1014. [Google Scholar] [CrossRef]
- Nasr, M.A.; Timmins, L.R.; Martin, V.J.J.; Kwan, D.H. A Versatile Transcription Factor Biosensor System Responsive to Multiple Aromatic and Indole Inducers. ACS Synth. Biol. 2022, 11, 1692–1698. [Google Scholar] [CrossRef]
- Smolke, C.D. Building outside of the box: iGEM and the BioBricks Foundation. Nat. Biotechnol. 2009, 27, 1099–1102. [Google Scholar] [CrossRef]
- Silva, K.P.T.; Chellamuthu, P.; Boedicker, J.Q. Quantifying the strength of quorum sensing crosstalk within microbial communities. PLoS Comput. Biol. 2017, 13, e1005809. [Google Scholar] [CrossRef]
- Schaefer, A.L.; Greenberg, E.P.; Oliver, C.M.; Oda, Y.; Huang, J.J.; Bittan-Banin, G.; Peres, C.M.; Schmidt, S.; Juhaszova, K.; Sufrin, J.R.; et al. A new class of homoserine lactone quorum-sensing signals. Nature 2008, 454, 595–599. [Google Scholar] [CrossRef]
- Kylilis, N.; Tuza, Z.A.; Stan, G.B.; Polizzi, K.M. Tools for engineering coordinated system behaviour in synthetic microbial consortia. Nat. Commun. 2018, 9, 2677. [Google Scholar] [CrossRef]
- Biarnes-Carrera, M.; Lee, C.K.; Nihira, T.; Breitling, R.; Takano, E. Orthogonal Regulatory Circuits for Escherichia coli Based on the gamma-Butyrolactone System of Streptomyces coelicolor. ACS Synth. Biol. 2018, 7, 1043–1055. [Google Scholar] [CrossRef] [PubMed]
- Green, P.J.; Pines, O.; Inouye, M. The role of antisense RNA in gene regulation. Annu. Rev. Biochem. 1986, 55, 569–597. [Google Scholar] [CrossRef] [PubMed]
- Na, D.; Yoo, S.M.; Chung, H.; Park, H.; Park, J.H.; Lee, S.Y. Metabolic engineering of Escherichia coli using synthetic small regulatory RNAs. Nat. Biotechnol. 2013, 31, 170–174. [Google Scholar] [CrossRef] [PubMed]
- Desai, S.K.; Gallivan, J.P. Genetic screens and selections for small molecules based on a synthetic riboswitch that activates protein translation. J. Am. Chem. Soc. 2004, 126, 13247–13254. [Google Scholar] [CrossRef]
- Isaacs, F.J.; Dwyer, D.J.; Ding, C.; Pervouchine, D.D.; Cantor, C.R.; Collins, J.J. Engineered riboregulators enable post-transcriptional control of gene expression. Nat. Biotechnol. 2004, 22, 841–847. [Google Scholar] [CrossRef] [PubMed]
- Yin, G.; Peng, A.; Zhang, L.; Wang, Y.; Du, G.; Chen, J.; Kang, Z. Design of artificial small regulatory trans-RNA for gene knockdown in Bacillus subtilis. Synth. Syst. Biotechnol. 2023, 8, 61–68. [Google Scholar] [CrossRef]
- Man, S.; Cheng, R.; Miao, C.; Gong, Q.; Gu, Y.; Lu, X.; Han, F.; Yu, W. Artificial trans-encoded small non-coding RNAs specifically silence the selected gene expression in bacteria. Nucleic Acids Res. 2011, 39, e50. [Google Scholar] [CrossRef]
- Bongaerts, J.; Kramer, M.; Muller, U.; Raeven, L.; Wubbolts, M. Metabolic engineering for microbial production of aromatic amino acids and derived compounds. Metab. Eng. 2001, 3, 289–300. [Google Scholar] [CrossRef]
- Matsumoto, T.; Tanaka, T.; Kondo, A. Engineering metabolic pathways in Escherichia coli for constructing a “microbial chassis” for biochemical production. Bioresour. Technol. 2017, 245, 1362–1368. [Google Scholar] [CrossRef]
- Peng, L.; Arauzo-Bravo, M.J.; Shimizu, K. Metabolic flux analysis for a ppc mutant Escherichia coli based on 13C-labelling experiments together with enzyme activity assays and intracellular metabolite measurements. Fems Microbiol. Lett. 2004, 235, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Morita, T.; El-Kazzaz, W.; Tanaka, Y.; Inada, T.; Aiba, H. Accumulation of glucose 6-phosphate or fructose 6-phosphate is responsible for destabilization of glucose transporter mRNA in Escherichia coli. J. Biol. Chem. 2003, 278, 15608–15614. [Google Scholar] [CrossRef] [PubMed]
- Gibson, D.G.; Young, L.; Chuang, R.Y.; Venter, J.C.; Hutchison, C.A., 3rd; Smith, H.O. Enzymatic assembly of DNA molecules up to several hundred kilobases. Nat. Methods 2009, 6, 343–345. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jin, X.; Gao, Y.; Chen, X.; Wang, S.; Qi, Q.; Liang, Q. The Construction of the Self-Induced Sal System and Its Application in Salicylic Acid Production. Molecules 2023, 28, 7825. https://doi.org/10.3390/molecules28237825
Jin X, Gao Y, Chen X, Wang S, Qi Q, Liang Q. The Construction of the Self-Induced Sal System and Its Application in Salicylic Acid Production. Molecules. 2023; 28(23):7825. https://doi.org/10.3390/molecules28237825
Chicago/Turabian StyleJin, Xin, Yaping Gao, Xuanmu Chen, Sumeng Wang, Qingsheng Qi, and Quanfeng Liang. 2023. "The Construction of the Self-Induced Sal System and Its Application in Salicylic Acid Production" Molecules 28, no. 23: 7825. https://doi.org/10.3390/molecules28237825
APA StyleJin, X., Gao, Y., Chen, X., Wang, S., Qi, Q., & Liang, Q. (2023). The Construction of the Self-Induced Sal System and Its Application in Salicylic Acid Production. Molecules, 28(23), 7825. https://doi.org/10.3390/molecules28237825