
Chemometric Evaluation of RI-Induced Phytochemicals in Phaseolus vulgaris Seeds Indicate an Improvement on Liver Enzymes in Obese Rats
 , and
, and
Abstract
:1. Introduction
2. Results and Discussion
2.1. Phytochemical Profile of Cooked Dalia Bean Grown under Different Irrigation Regimes
2.2. Effect of Cooked Dalia Bean Intake on Liver Enzymes
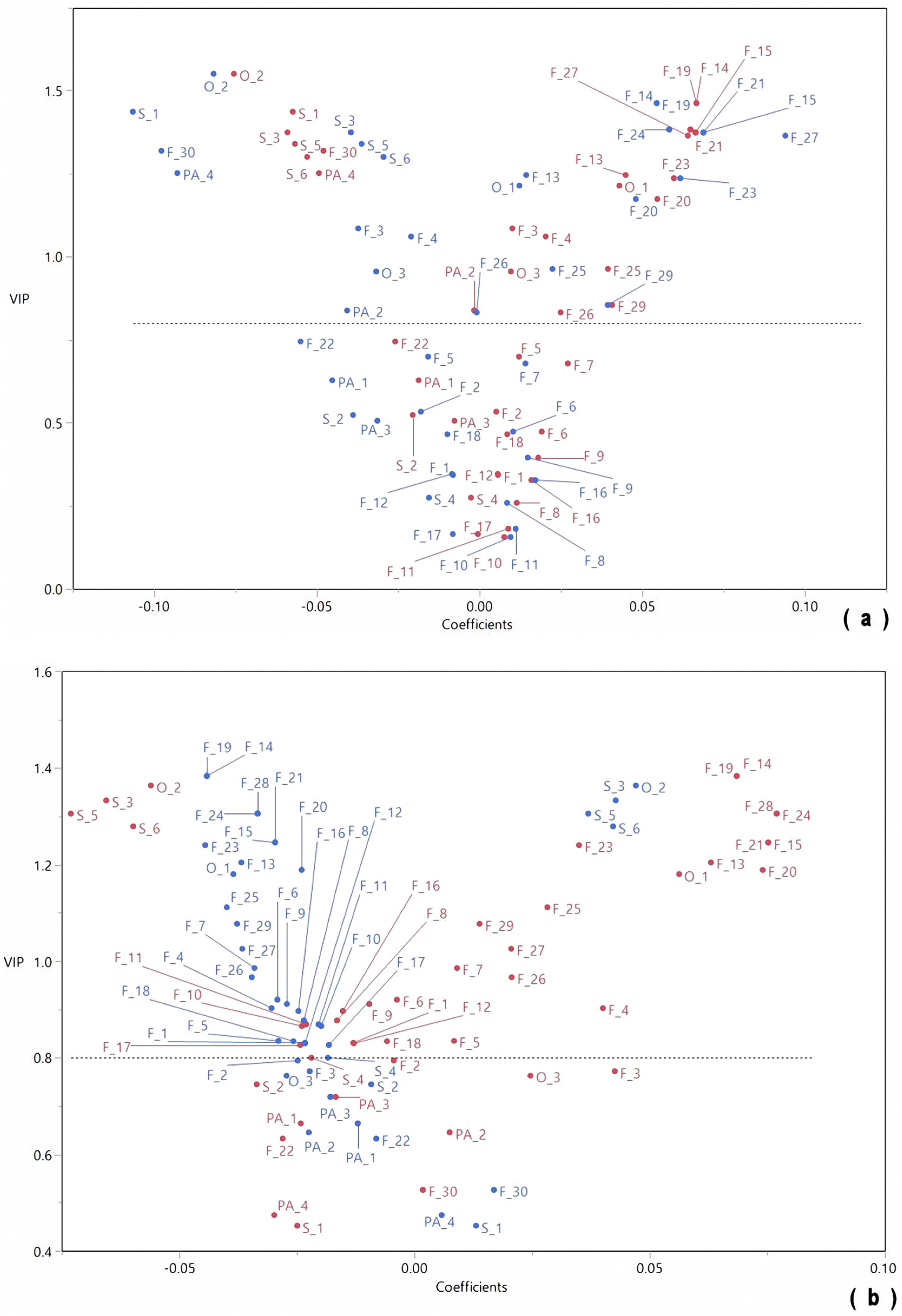
2.3. Chemometric Analysis
3. Material and Methods
3.1. Dalia Bean Origin and Selection
3.2. Experimental Design Followed for Bean Production
3.3. Sample Preparation and Phytochemicals Quantification
3.4. Experimental Animals
3.5. Quantification of Liver Enzymes Activity
4. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Khan, A.R.; Awan, F.R.; Najam, S.S.; Islam, M.; Siddique, T.; Zain, M. Elevated serum level of human alkaline phosphatase in obesity. J. Pak. Med. Assoc. 2015, 65, 1182–1185. [Google Scholar]
- Liu, C.; Shao, M.; Lu, L.; Zhao, C.; Qiu, L.; Liu, Z. Obesity, insulin resistance and their interaction on liver enzymes. PLoS ONE 2021, 16, e0249299. [Google Scholar] [CrossRef]
- Bekkelund, S.I.; Jorde, R. Alanine Aminotransferase and Body Composition in Obese Men and Women. Dis. Markers 2019, 2019, 1695874. [Google Scholar] [CrossRef] [PubMed]
- Ndrepepa, G.; Colleran, R.; Kastrati, A. Gamma-glutamyl transferase and the risk of atherosclerosis and coronary heart disease. Clin. Chim. Acta 2018, 476, 130–138. [Google Scholar] [CrossRef] [PubMed]
- Nolan, R.; Shannon, O.M.; Robinson, N.; Joel, A.; Houghton, D.; Malcomson, F.C. It’s No Has Bean: A Review of the Effects of White Kidney Bean Extract on Body Composition and Metabolic Health. Nutrients 2020, 12, 1398. [Google Scholar] [CrossRef] [PubMed]
- Viguiliouk, E.; Blanco Mejia, S.; Kendall, C.W.; Sievenpiper, J.L. Can pulses play a role in improving cardiometabolic health? Evidence from systematic reviews and meta-analyses. Ann. N. Y. Acad. Sci. 2017, 1392, 43–57. [Google Scholar] [CrossRef] [PubMed]
- Food and Agriculture Organization. 2021. Available online: https://www.fao.org/faostat/en/#data/FBS (accessed on 23 March 2023).
- Manoj, B.S.; Gupta, M.; Jeelani, M.I.; Gupta, S. Metabolic footprints of chitosan primed red kidney bean under restricted irrigation. Int. J. Biol. Macromol. 2022, 208, 367–380. [Google Scholar] [CrossRef] [PubMed]
- Herrera, M.D.; Reynoso-Camacho, R.; Melero-Meraz, V.; Guzmán-Maldonado, S.H.; Acosta-Gallegos, J.A. Impact of soil moisture on common bean (Phaseolus vulgaris L.) phytochemicals. J. Food Compos. Anal. 2021, 99, 103883. [Google Scholar] [CrossRef]
- Herrera, M.D.; Acosta-Gallegos, J.A.; Reynoso-Camacho, R.; Perez-Ramirez, I.F. Common bean seeds from plants subjected to severe drought, restricted- and full-irrigation regimes show differential phytochemical fingerprint. Food Chem. 2019, 294, 368–377. [Google Scholar] [CrossRef] [PubMed]
- Coku, V.; Shkembi, X. Serum Gamma-glutamyltransferase and Obesity: Is there a Link? Med. Arch. 2018, 72, 112–115. [Google Scholar] [CrossRef]
- Farouk, S.; Amany, A.R. Improving growth and yield of cowpea by foliar application of chitosan under water stress. Egypt. J. Biol. 2012, 14, 14–16. [Google Scholar] [CrossRef]
- Perez-Alvarez, E.P.; Intrigliolo, D.S.; Almajano, M.P.; Rubio-Breton, P.; Garde-Cerdan, T. Effects of Water Deficit Irrigation on Phenolic Composition and Antioxidant Activity of Monastrell Grapes under Semiarid Conditions. Antioxidants 2021, 10, 1301. [Google Scholar] [CrossRef] [PubMed]
- Agati, G.; Azzarello, E.; Pollastri, S.; Tattini, M. Flavonoids as antioxidants in plants: Location and functional significance. Plant Sci. 2012, 196, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Monika, M.; Yadav, S.K. Effect of quercetin and epicatechin on the transcript expression and activity of antioxidant enzymes in tobacco seedlings. Am. J. Biochem. Mol. Biol. 2013, 3, 81–90. [Google Scholar]
- Zarrouk, O.; Brunetti, C.; Egipto, R.; Pinheiro, C.; Genebra, T.; Gori, A.; Lopes, C.M.; Tattini, M.; Chaves, M.M. Grape Ripening Is Regulated by Deficit Irrigation/Elevated Temperatures According to Cluster Position in the Canopy. Front. Plant Sci. 2016, 7, 1640. [Google Scholar] [CrossRef] [PubMed]
- Yildiztugay, E.; Ozfidan-Konakci, C.; Kucukoduk, M.; Turkan, I. Flavonoid Naringenin Alleviates Short-Term Osmotic and Salinity Stresses Through Regulating Photosynthetic Machinery and Chloroplastic Antioxidant Metabolism in Phaseolus vulgaris. Front. Plant Sci. 2020, 11, 682. [Google Scholar] [CrossRef] [PubMed]
- Gundaraniya, S.A.; Ambalam, P.S.; Tomar, R.S. Metabolomic Profiling of Drought-Tolerant and Susceptible Peanut (Arachis hypogaea L.) Genotypes in Response to Drought Stress. ACS Omega 2020, 5, 31209–31219. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.K.; Kaur, N. Sugar signalling and gene expression in relation to carbohydrate metabolism under abiotic stresses in plants. J. Biosci. 2005, 30, 761–776. [Google Scholar] [CrossRef]
- Shafiee-Nick, R.; Ghorbani, A.; Vafaee Bagheri, F.; Rakhshandeh, H. Chronic administration of a combination of six herbs inhibits the progression of hyperglycemia and decreases serum lipids and aspartate amino transferase activity in diabetic rats. Adv. Pharmacol. Sci. 2012, 2012, 789796. [Google Scholar] [CrossRef]
- Nagell, W.; Tonus, C.; Nier, H.; Dancygier, H. Primary prophylactic therapy of esophageal varices—A literature review. Hepatogastroenterology 2002, 49, 423–427. [Google Scholar]
- Salas-Lumbreras, G.; Reveles-Torres, L.R.; Servin-Palestina, M.; Acosta-Gallegos, J.A.; Herrera, M.D.; Reyes-Estrada, C.A.; Lopez, J.A. Common Bean Seeds Obtained by Plant Water Restriction Ameliorates Obesity-Associated Cardiovascular Risk and Insulin Resistance. Plant Foods Hum. Nutr. 2023, 78, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Rej, R. Measurement of aminotransferases: Part 1. Aspartate aminotransferase. Crit. Rev. Clin. Lab. Sci. 1984, 21, 99–186. [Google Scholar] [CrossRef] [PubMed]
- Akuyam, S.A.; Abubakar, A.; Lawal, N.; Yusuf, R.; Aminu, S.M.; Hassan, A.; Musa, A.; Bello, A.K.; Yahaya, I.A.; Okafor, P.A. Assessment of biochemical liver function tests in relation to age among steady state sickle cell anemia patients. Niger. J. Clin. Pract. 2017, 20, 1428–1433. [Google Scholar] [CrossRef] [PubMed]
- Oh, R.C.; Hustead, T.R.; Ali, S.M.; Pantsari, M.W. Mildly Elevated Liver Transaminase Levels: Causes and Evaluation. Am. Fam. Physician 2017, 96, 709–715. [Google Scholar] [PubMed]
- Corti, A.; Belcastro, E.; Dominici, S.; Maellaro, E.; Pompella, A. The dark side of gamma-glutamyltransferase (GGT): Pathogenic effects of an ‘antioxidant’ enzyme. Free Radic. Biol. Med. 2020, 160, 807–819. [Google Scholar] [CrossRef] [PubMed]
- Khan, K.N.; Tsutsumi, T.; Nakata, K.; Nakao, K.; Kato, Y.; Nagataki, S. Regulation of alkaline phosphatase gene expression in human hepatoma cells by bile acids. J. Gastroenterol. Hepatol. 1998, 13, 643–650. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.W. Association between elevated serum hepatic enzyme activity and total body fat in obese humans. Ann. Clin. Lab. Sci. 2003, 33, 257–264. [Google Scholar]
- Biancolillo, A.; Marini, F. Chemometric Methods for Spectroscopy-Based Pharmaceutical Analysis. Front. Chem. 2018, 6, 576. [Google Scholar] [CrossRef]
- Acosta-Gallegos, J.A.; Montero-Tavera, V.; Jiménez-Hernández, Y.; Anaya-López, J.L.; Gonzalez-Chavira, M.M. ‘Dalia’, a new variety of bean grain, Flor de Junio type for the central region of Mexico. Rev. Mex. Cienc. Agric. 2014, 5, 331–336. [Google Scholar] [CrossRef]
- Servicion Nacional de Inección y Certificación de Semillas. SNICS (Catálogo Nac. De Variedades Vegetales) 2023. Available online: https://www.gob.mx/snics (accessed on 2 June 2023).
- Reynoso-Camacho, R.; Rodriguez-Villanueva, L.D.; Sotelo-Gonzalez, A.M.; Ramos-Gomez, M.; Perez-Ramirez, I.F. Citrus decoction by-product represents a rich source of carotenoid, phytosterol, extractable and non-extractable polyphenols. Food Chem. 2021, 350, 129239. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family | Code | Component Name | Retention Time (min) | Molecular Formula | Expected Mass (Da) | Observed Mass (Da) | Observed m/z †† | Mass Error (ppm) | Soil Moisture Treatments | p Value | |||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 100/100 | 100/50 | 50/50 | 50/100 | ||||||||||
| Flavanols | F_1 | (−)-Epicatechin hexoside | 2.05 | C21H24O11 | 452.1319 | 452.1332 | 451.1259 | 3.00 | 15.10 ± 1.78 b | 31.75 ± 4.15 a | 14.13 ± 1.71 b | 17.71 ± 2.19 b | 0.0087 |
| F_2 | (+)-Catechin hexoside | 2.61 | C21H24O11 | 452.1319 | 452.1330 | 451.1257 | 2.50 | 4.52 ± 0.58 ab | 7.24 ± 0.93 a | 3.84 ± 0.48 b | 5.15 ± 0.67 ab | 0.0288 | |
| F_3 | (+)-Catechin † | 2.62 | C15H14O6 | 290.0790 | 290.0798 | 289.0725 | 2.56 | 14.73 ± 1.15 a | 14.62 ± 1.16 a | 12.93 ± 1.01 a | 15.07 ± 1.23 a | 0.3625 | |
| F_4 | (−)-Epicatechin † | 3.03 | C15H14O6 | 290.0790 | 290.0798 | 289.0726 | 2.75 | 5.76 ± 0.38 a | 5.91 ± 0.44 a | 4.95 ± 0.34 a | 5.81 ± 0.44 a | 0.2036 | |
| F_5 | Procyanidin dimer B2 † | 3.12 | C30H26O12 | 578.1424 | 578.1434 | 577.1361 | 1.67 | 0.66 ± 0.16 a | 1.20 ± 0.36 a | 0.36 ± 0.10 a | 0.76 ± 0.22 a | 0.0868 | |
| Flavonols | F_6 | Quercetin hexoside-rhamnoside | 3.87 | C27H30O16 | 610.1534 | 610.1537 | 609.1464 | 0.56 | 0.63 ± 0.13 a | 0.94 ± 0.19 a | 0.57 ± 0.12 a | 0.62 ± 0.13 a | 0.1812 |
| F_7 | Quercetin hexoside | 4.26 | C21H20O12 | 464.0955 | 464.0964 | 463.0891 | 1.90 | 3.78 ± 0.07 b | 5.30 ± 0.62 b | 3.03 ± 0.01 b | 3.53 ± 0.07 a | 0.0076 | |
| F_8 | Kaempferol pentosyl-hexoside | 4.30 | C26H28O15 | 580.1428 | 580.1434 | 579.1361 | 0.96 | 4.47 ± 0.04 b | 10.04 ± 0.44 a | 4.73 ± 0.03 b | 4.59 ± 0.00 b | <0.0001 | |
| F_9 | Kaempferol hexoside | 5.26 | C21H20O11 | 448.1006 | 448.1015 | 447.0942 | 2.00 | 217.70 ± 2.18 b | 256.42 ± 4.95 a | 215.44 ± 2.79 b | 215.25 ± 2.38 b | 0.0006 | |
| F_10 | Kaempferol acetyl-hexoside | 6.28 | C23H22O12 | 490.1111 | 490.1116 | 489.1043 | 0.97 | 3.81 ± 0.48 b | 10.26 ± 1.33 a | 4.97 ± 0.64 b | 4.06 ± 0.54 b | 0.0041 | |
| F_11 | Kaempferol malonyl-hexoside | 6.33 | C24H22O14 | 534.1010 | 534.1008 | 533.0935 | −0.36 | 0.30 ± 0.04 b | 0.65 ± 0.14 a | 0.36 ± 0.05 ab | 0.31 ± 0.07 ab | 0.0408 | |
| F_12 | Quercetin rhamnoside | 6.52 | C21H20O11 | 448.1006 | 448.1013 | 447.0940 | 1.68 | 6.43 ± 0.70 a | 9.12 ± 1.02 a | 6.27 ± 0.68 a | 6.86 ± 0.72 a | 0.0649 | |
| F_13 | Quercetin acetyl-rhamnoside | 8.84 | C23H22O12 | 490.1111 | 490.1108 | 489.1035 | −0.69 | 0.24 ± 0.03 a | 0.21 ± 0.01ab | 0.13 ± 0.00 b | 0.20 ± 0.04 ab | 0.0460 | |
| F_14 | Quercetin † | 8.99 | C15H10O7 | 302.0427 | 302.0424 | 301.0351 | −0.98 | 0.13 ± 0.02 a | 0.11 ± 0.01 ab | 0.07 ± 0.01 b | 0.09 ± 0.01 ab | 0.0290 | |
| F_15 | Kaempferol † | 11.12 | C15H10O6 | 286.0477 | 286.0469 | 285.0396 | −3.03 | 17.55 ± 1.51 a | 7.34 ± 0.57 b | 6.56 ± 0.50 b | 8.04 ± 0.63 b | 0.0008 | |
| Flavones | F_16 | Luteolin apiosyl-hexoside | 4.30 | C26H28O15 | 580.1428 | 580.1434 | 579.1361 | 0.96 | 4.53 ± 0.00 c | 9.48 ± 0.10 a | 4.80 ± 0.03 b | 4.23 ± 0.00 d | <0.0001 |
| F_17 | Luteolin † | 6.25 | C15H10O6 | 286.0477 | 286.0479 | 285.0406 | 0.65 | 0.07 ± 0.01 b | 0.14 ± 0.02 a | 0.08 ± 0.01 b | 0.08 ± 0.01 b | 0.0070 | |
| F_18 | (Iso)rhoifolin | 8.85 | C27H30O14 | 578.1636 | 578.1614 | 577.1541 | −3.72 | 0.12 ± 0.03 a | 0.19 ± 0.03 a | 0.10 ± 0.02 a | 0.13 ± 0.02 a | 0.1160 | |
| F_19 | Hydroxyluteolin | 8.99 | C15H10O7 | 302.0427 | 302.0424 | 301.0351 | −0.98 | 0.12 ± 0.01 a | 0.10 ± 0.01 ab | 0.06 ± 0.01 b | 0.08 ± 0.01 ab | 0.0290 | |
| F_20 | Sinensetin | 11.02 | C20H20O7 | 372.1209 | 372.1208 | 371.1135 | −0.34 | 2.34 ± 0.25 a | 0.37 ± 0.05 c | 0.31 ± 0.04 c | 0.96 ± 0.11 b | 0.0004 | |
| F_21 | Scutellarein | 11.12 | C15H10O6 | 286.0477 | 286.0469 | 285.0396 | −3.03 | 16.18 ± 1.39 a | 6.77 ± 0.53 b | 6.05 ± 0.46 b | 7.41 ± 0.58 b | 0.0008 | |
| Isoflavones | F_22 | Genistein malonyl-hexoside (malonylgenistin) | 3.49 | C24H22O13 | 518.1060 | 518.1051 | 517.0978 | −1.81 | 1.37 ± 0.18 a | 2.15 ± 0.29 a | 1.45 ± 0.19 a | 1.83 ± 0.25 a | 0.0853 |
| F_23 | Daidzein hexoside (daidzin) | 3.98 | C21H20O9 | 416.1107 | 416.1124 | 415.1051 | 3.91 | 0.03 ± 0.00 a | 0.05 ± 0.01 a | ND | ND | 0.0010 | |
| F_24 | Tetrahydroxyisoflavanone | 5.51 | C15H12O6 | 288.0634 | 288.0636 | 287.0563 | 0.80 | 6.67 ± 0.46 a | 4.24 ± 0.32 b | 3.42 ± 0.25 b | 4.35 ± 0.30 b | 0.0027 | |
| F_25 | Genistein † | 5.75 | C15H10O5 | 270.0528 | 270.0531 | 269.0458 | 1.06 | 0.11 ± 0.01 ab | 0.13 ± 0.02 a | 0.07 ± 0.01 b | 0.09 ± 0.01 ab | 0.0213 | |
| F_26 | Daidzein † | 7.63 | C15H10O4 | 254.0579 | 254.0574 | 253.0501 | −2.12 | 0.15 ± 0.01 ab | 0.20 ± 0.03 a | 0.10 ± 0.01 b | 0.14 ± 0.01 ab | 0.0205 | |
| F_27 | Trihydroxyisoflavan | 8.24 | C15H14O4 | 258.0892 | 258.0888 | 257.0816 | −1.44 | 0.06 ± 0.01 ab | 0.08 ± 0.01 a | 0.05 ± 0.00 b | ND | 0.0006 | |
| Flavanones | F_28 | Eriodictyol | 5.52 | C15H12O6 | 288.0634 | 288.0636 | 287.0563 | 0.80 | 6.45 ± 0.44 a | 4.11 ± 0.31 b | 3.31 ± 0.25 b | 4.21 ± 0.29 b | 0.0027 |
| F_29 | Naringenin hexoside | 5.75 | C21H22O10 | 434.1213 | 434.1221 | 433.1148 | 1.77 | 0.25 ± 0.04 a | 0.34 ± 0.06 a | 0.19 ± 0.03 a | 0.19 ± 0.04 a | 0.0687 | |
| F_30 | Naringenin † | 10.95 | C15H12O5 | 272.0685 | 272.0676 | 271.0603 | −3.36 | 0.25 ± 0.02 b | 0.23 ± 0.02 b | 0.20 ± 0.02 b | 0.43 ± 0.04 a | 0.0031 | |
| Hydroxybenzoic acids | PA_1 | Vanillic acid † | 1.79 | C8H8O4 | 168.0423 | 168.0417 | 167.0344 | −3.26 | 0.49 ± 0.03 c | 0.78 ± 0.06 a | 0.50 ± 0.04 b | 0.63 ± 0.04 ab | 0.0063 |
| PA_2 | Dihydroxybenzoic acid | 1.91 | C7H6O4 | 154.0266 | 154.0263 | 153.0190 | −2.04 | 0.29 ± 0.00 b | 0.42 ± 0.02 a | 0.21 ± 0.01 c | 0.36 ± 0.02 a | 0.0007 | |
| PA_3 | Hydroxybenzoic acid hexoside | 2.10 | C13H16O8 | 300.0845 | 300.0841 | 299.0768 | −1.36 | 0.32 ± 0.02 a | 0.79 ± 0.26 a | 0.30 ± 0.02 a | 0.49 ± 0.14 a | 0.0889 | |
| Hydroxycinnamic acids | PA_4 | Ferulic acid † | 3.96 | C10H10O4 | 194.0579 | 194.0579 | 193.0507 | 0.10 | 0.08 ± 0.01 b | 0.12 ± 0.01 a | 0.09 ± 0.01 b | 0.13 ± 0.01 a | 0.0072 |
| Oligosaccharides | O_1 | Raffinose † | 0.58 | C18H32O16 | 504.1690 | 504.1705 | 503.1632 | 2.84 | 1.98 ± 0.25 a | 1.96 ± 0.23 a | 1.74 ± 0.22 a | 1.91 ± 0.27 a | 0.7628 |
| O_2 | Stachyose | 1.27 | C24H42O21 | 666.2219 | 666.2217 | 665.2144 | −0.26 | 4.21 ± 0.65 a | 4.31 ± 0.71 a | 4.97 ± 0.73 a | 4.99 ± 0.76 a | 0.6086 | |
| O_3 | Verbascose | 3.13 | C30H52O26 | 828.2747 | 828.2707 | 827.2634 | −4.79 | 0.11 ± 0.05 a | 0.17 ± 0.07 a | ND | 0.14 ± 0.08 a | 0.1405 | |
| Saponins | S_1 | Soyasaponin Ba (V) | 11.54 | C48H78O19 | 958.5137 | 958.5114 | 957.5041 | −2.47 | 48.17 ± 1.95 a | 51.13 ± 0.60 a | 48.52 ± 1.87 a | 53.40 ± 0.55 a | 0.0575 |
| S_2 | Soyasaponin Bb (I) † | 11.58 | C48H78O18 | 942.5188 | 942.5166 | 941.5093 | −2.34 | 11.11 ± 3.75 a | 12.78 ± 4.33 a | 11.46 ± 3.77 a | 11.85 ± 4.00 a | 0.975 | |
| S_3 | Soyasaponin ag | 11.68 | C54H84O22 | 1084.5454 | 1084.5450 | 1083.5378 | −0.36 | 18.21 ± 0.65 b | 18.78 ± 0.07 b | 20.69 ± 0.29 a | 19.49 ± 0.15 ab | 0.0098 | |
| S_4 | Soyasaponin Bd | 11.71 | C48H76O19 | 956.4981 | 956.4968 | 955.4895 | −1.31 | 2.07 ± 0.55 a | 5.66 ± 1.52 a | 2.34 ± 0.61 a | 2.92 ± 0.79 a | 0.0578 | |
| S_5 | Soyasaponin bg | 11.74 | C54H84O21 | 1068.5505 | 1068.5499 | 1067.5427 | −0.54 | 3.16 ± 0.96 a | 3.69 ± 1.48 a | 4.27 ± 1.26 a | 3.71 ± 1.12 a | 0.8414 | |
| S_6 | Soyasaponin gg | 11.80 | C48H74O17 | 922.4926 | 922.4919 | 921.4846 | −0.80 | 2.30 ± 0.97 a | 2.35 ± 1.00 a | 2.84 ± 1.02 a | 2.53 ± 1.02 a | 0.9443 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Herrera, M.D.; Pérez-Ramírez, I.F.; Reynoso-Camacho, R.; Reveles-Torres, L.R.; Servín-Palestina, M.; Granados-López, A.J.; Reyes-Estrada, C.A.; López, J.A. Chemometric Evaluation of RI-Induced Phytochemicals in Phaseolus vulgaris Seeds Indicate an Improvement on Liver Enzymes in Obese Rats. Molecules 2023, 28, 7983. https://doi.org/10.3390/molecules28247983
Herrera MD, Pérez-Ramírez IF, Reynoso-Camacho R, Reveles-Torres LR, Servín-Palestina M, Granados-López AJ, Reyes-Estrada CA, López JA. Chemometric Evaluation of RI-Induced Phytochemicals in Phaseolus vulgaris Seeds Indicate an Improvement on Liver Enzymes in Obese Rats. Molecules. 2023; 28(24):7983. https://doi.org/10.3390/molecules28247983
Chicago/Turabian StyleHerrera, Mayra Denise, Iza Fernanda Pérez-Ramírez, Rosalía Reynoso-Camacho, Luis Roberto Reveles-Torres, Miguel Servín-Palestina, Angelica Judith Granados-López, Claudia Araceli Reyes-Estrada, and Jesús Adrián López. 2023. "Chemometric Evaluation of RI-Induced Phytochemicals in Phaseolus vulgaris Seeds Indicate an Improvement on Liver Enzymes in Obese Rats" Molecules 28, no. 24: 7983. https://doi.org/10.3390/molecules28247983
APA StyleHerrera, M. D., Pérez-Ramírez, I. F., Reynoso-Camacho, R., Reveles-Torres, L. R., Servín-Palestina, M., Granados-López, A. J., Reyes-Estrada, C. A., & López, J. A. (2023). Chemometric Evaluation of RI-Induced Phytochemicals in Phaseolus vulgaris Seeds Indicate an Improvement on Liver Enzymes in Obese Rats. Molecules, 28(24), 7983. https://doi.org/10.3390/molecules28247983