
Anthocyanins: Potential Therapeutic Approaches towards Obesity and Diabetes Mellitus Type 2


 ,
,
Abstract
:1. Introduction
2. Phytochemicals
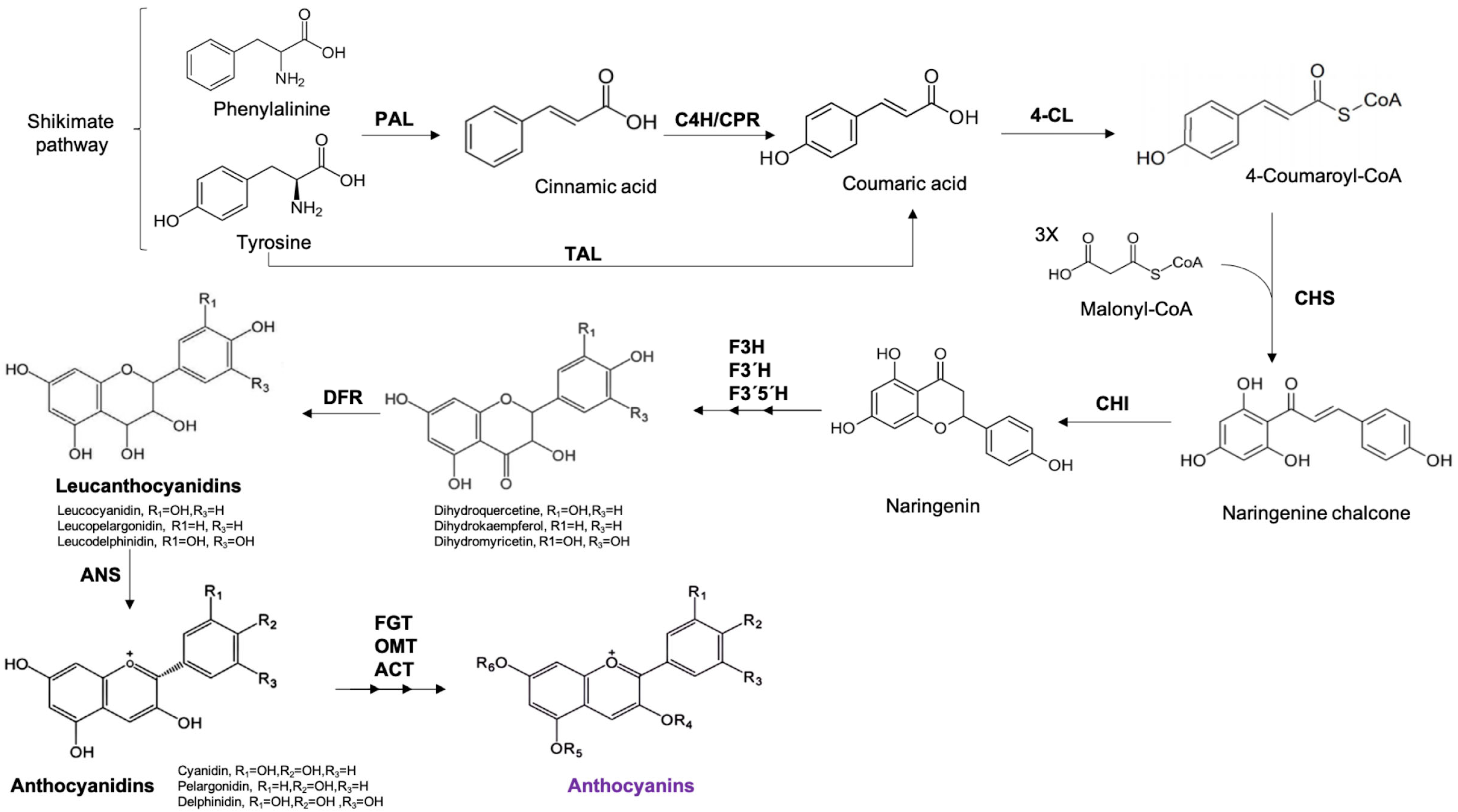
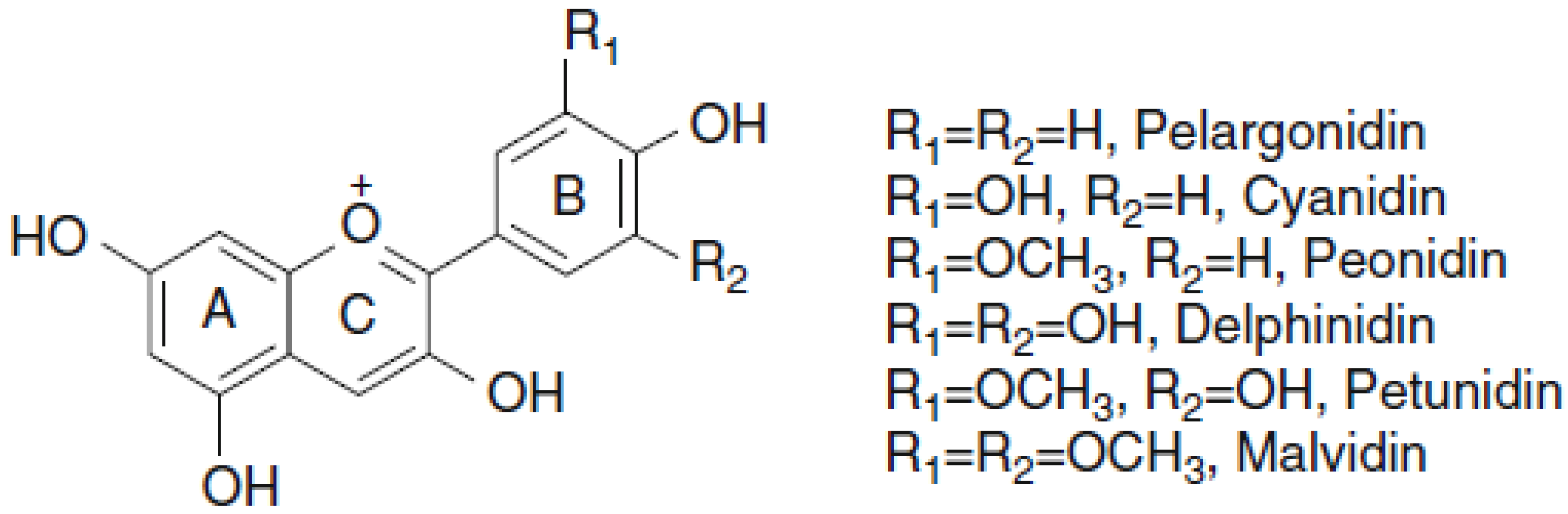
Structure, Classification, and Function of Anthocyanins
Anthocyanins in Pigmented Corn as an Example of Functional Food
3. Mechanisms of Absorption, Metabolism, and Bioavailability of Polyphenols
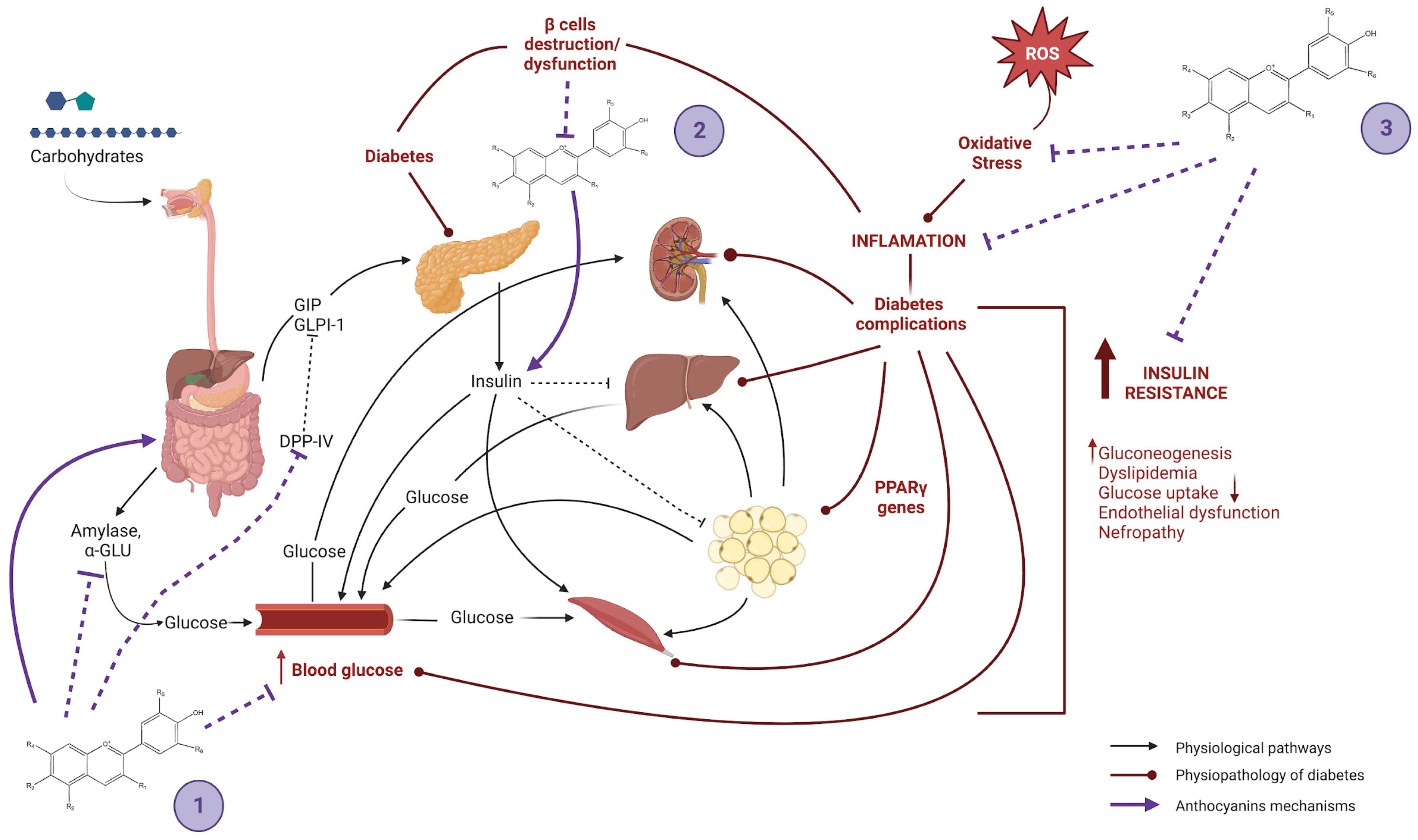
4. Chemical Nature, Reported Mechanism of Action and Assigned Effects
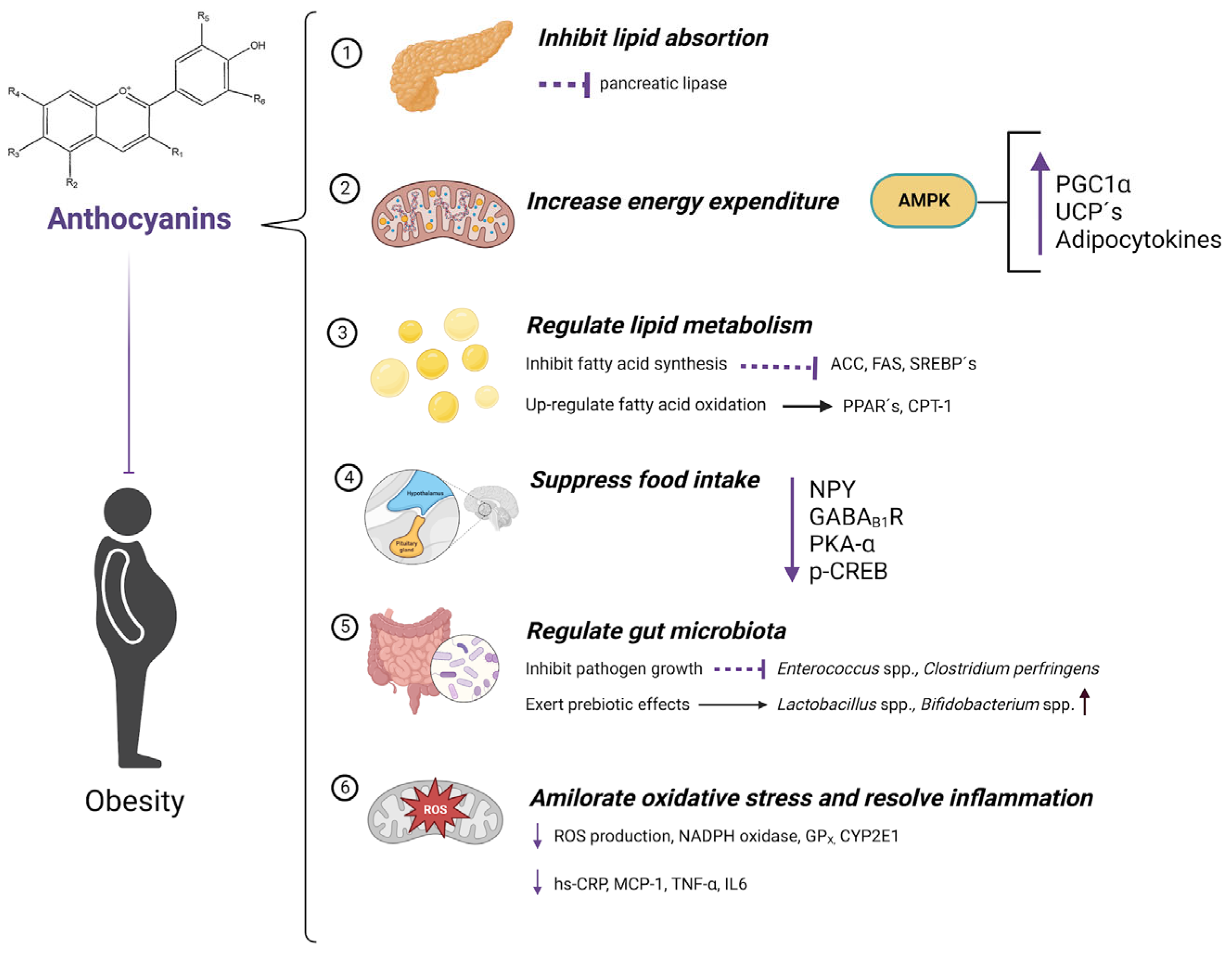
4.1. Reduction of Adipogenesis
4.2. Antioxidant Properties
4.3. Anti-Inflammatory Activity
5. Discussion
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Abdel-Aal, E.-S.M.; Young, J.C.; Rabalski, I. Anthocyanin Composition in Black, Blue, Pink, Purple, and Red Cereal Grains. J. Agric. Food Chem. 2006, 54, 4696–4704. [Google Scholar] [CrossRef]
- Abubakar, S.M.; Ukeyima, M.T.; Spencer, J.P.E.; Lovegrove, J.A. Acute Effects of Hibiscus Sabdariffa Calyces on Postprandial Blood Pressure, Vascular Function, Blood Lipids, Biomarkers of Insulin Resistance and Inflammation in Humans. Nutrients 2019, 11, 341. [Google Scholar] [CrossRef] [Green Version]
- Arendt, E.K.; Zannini, E. Cereal Grains for the Food and Beverage Industries, 1st ed.; Elsevier: Berkeley, CA, USA, 2013. [Google Scholar]
- Călinoiu, L.F.; Vodnar, D.C. Whole Grains and Phenolic Acids: A Review on Bioactivity, Functionality, Health Benefits and Bioavailability. Nutrients 2018, 10, 1615. [Google Scholar] [CrossRef] [Green Version]
- Castañeda-Ovando, A.; de Lourdes Pacheco-Hernández, M.; Páez-Hernández, M.E.; Rodríguez, J.A.; Galán-Vidal, C.A. Chemical Studies of Anthocyanins: A Review. Food Chem. 2009, 113, 859–871. [Google Scholar] [CrossRef]
- Castro-Acosta, M.L.; Stone, S.G.; Mok, J.E.; Mhajan, R.K.; Fu, C.-I.; Lenihan-Geels, G.N.; Corpe, C.P.; Hall, W.L. Apple and Blackcurrant Polyphenol-Rich Drinks Decrease Postprandial Glucose, Insulin and Incretin Response to a High-Carbohydrate Meal in Healthy Men and Women. J. Nutr. Biochem. 2017, 49, 53–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cevallos-Casals, B.A.; Cisneros-Zevallos, L. Stoichiometric and Kinetic Studies of Phenolic Antioxidants from Andean Purple Corn and Red-Fleshed Sweetpotato. J. Agric. Food Chem. 2003, 51, 3313–3319. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-F.; Shibu, M.A.; Fan, M.-J.; Chen, M.-C.; Viswanadha, V.P.; Lin, Y.-L.; Lai, C.-H.; Lin, K.-H.; Ho, T.-J.; Kuo, W.-W.; et al. Purple Rice Anthocyanin Extract Protects Cardiac Function in STZ-Induced Diabetes Rat Hearts by Inhibiting Cardiac Hypertrophy and Fibrosis. J. Nutr. Biochem. 2016, 31, 98–105. [Google Scholar] [CrossRef]
- Cueva, C.; Gil-Sánchez, I.; Ayuda-Durán, B.; González-Manzano, S.; González-Paramás, A.; Santos-Buelga, C.; Bartolomé, B.; Moreno-Arribas, M. An Integrated View of the Effects of Wine Polyphenols and Their Relevant Metabolites on Gut and Host Health. Molecules 2017, 22, 99. [Google Scholar] [CrossRef]
- de Pascual-Teresa, S.; Sanchez-Ballesta, M.T. Anthocyanins: From Plant to Health. Phytochem. Rev. 2008, 7, 281–299. [Google Scholar] [CrossRef]
- del Pozo-Insfran, D.; Brenes, C.H.; Saldivar, S.O.S.; Talcott, S.T. Polyphenolic and Antioxidant Content of White and Blue Corn (Zea Mays L.) Products. Food Res. Int. 2006, 39, 696–703. [Google Scholar] [CrossRef]
- Enaru, B.; Drețcanu, G.; Pop, T.D.; Stǎnilǎ, A.; Diaconeasa, Z. Anthocyanins: Factors Affecting Their Stability and Degradation. Antioxidants 2021, 10, 1967. [Google Scholar] [CrossRef] [PubMed]
- Escribano-Bailón, M.T.; Santos-Buelga, C.; Rivas-Gonzalo, J.C. Anthocyanins in Cereals. J. Chromatogr. A 2004, 1054, 129–141. [Google Scholar] [CrossRef]
- Esposito, D.; Damsud, T.; Wilson, M.; Grace, M.H.; Strauch, R.; Li, X.; Lila, M.A.; Komarnytsky, S. Black Currant Anthocyanins Attenuate Weight Gain and Improve Glucose Metabolism in Diet-Induced Obese Mice with Intact, but Not Disrupted, Gut Microbiome. J. Agric. Food Chem. 2015, 63, 6172–6180. [Google Scholar] [CrossRef] [PubMed]
- Fardet, A. New Hypotheses for the Health-Protective Mechanisms of Whole-Grain Cereals: What Is beyond Fibre? Nutr. Res. Rev. 2010, 23, 65–134. [Google Scholar] [CrossRef] [Green Version]
- Faria, A.; Pestana, D.; Azevedo, J.; Martel, F.; de Freitas, V.; Azevedo, I.; Mateus, N.; Calhau, C. Absorption of Anthocyanins through Intestinal Epithelial Cells—Putative Involvement of GLUT2. Mol. Nutr. Food Res. 2009, 53, 1430–1437. [Google Scholar] [CrossRef] [PubMed]
- Forouhi, N.G.; Wareham, N.J. Epidemiology of Diabetes. Medicine 2014, 42, 698–702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Francavilla, A.; Joye, I.J. Anthocyanins in Whole Grain Cereals and Their Potential Effect on Health. Nutrients 2020, 12, 2922. [Google Scholar] [CrossRef] [PubMed]
- Gani, A.; SM, W.; FA, M. Whole-Grain Cereal Bioactive Compounds and Their Health Benefits: A Review. J. Food Process Technol. 2012, 3. [Google Scholar] [CrossRef] [Green Version]
- Gowd, V.; Jia, Z.; Chen, W. Anthocyanins as Promising Molecules and Dietary Bioactive Components against Diabetes—A Review of Recent Advances. Trends Food Sci. Technol. 2017, 68, 1–13. [Google Scholar] [CrossRef]
- Guo, X.; Yang, B.; Tan, J.; Jiang, J.; Li, D. Associations of Dietary Intakes of Anthocyanins and Berry Fruits with Risk of Type 2 Diabetes Mellitus: A Systematic Review and Meta-Analysis of Prospective Cohort Studies. Eur. J. Clin. Nutr. 2016, 70, 1360–1367. [Google Scholar] [CrossRef]
- Haeusler, R.A.; Camastra, S.; Astiarraga, B.; Nannipieri, M.; Anselmino, M.; Ferrannini, E. Decreased Expression of Hepatic Glucokinase in Type 2 Diabetes. Mol. Metab. 2015, 4, 222–226. [Google Scholar] [CrossRef] [PubMed]
- Hershey, M.S.; Sotos-Prieto, M.; Ruiz-Canela, M.; Martinez-Gonzalez, M.A.; Cassidy, A.; Moffatt, S.; Kales, S.N. Anthocyanin Intake and Physical Activity: Associations with the Lipid Profile of a US Working Population. Molecules 2020, 25, 4398. [Google Scholar] [CrossRef]
- Hidalgo, J.; Teuber, S.; Morera, F.; Ojeda, C.; Flores, C.; Hidalgo, M.; Núñez, L.; Villalobos, C.; Burgos, R. Delphinidin Reduces Glucose Uptake in Mice Jejunal Tissue and Human Intestinal Cells Lines through FFA1/GPR40. Int. J. Mol. Sci. 2017, 18, 750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Q.; Xu, J. Profiles of Carotenoids, Anthocyanins, Phenolics, and Antioxidant Activity of Selected Color Waxy Corn Grains during Maturation. J. Agric. Food Chem. 2011, 59, 2026–2033. [Google Scholar] [CrossRef] [PubMed]
- International Diabetes Federation. IDF Diabetes Atlas, 8th ed.; IDF: Brussels, Belgium, 2017. [Google Scholar]
- Jayaprakasa, B.; Vareed, S.K.; Olson, L.K.; Nair, M.G. Insulin Secretion by Bioactive Anthocyanins and Anthocyanidins Present in Fruits. J. Agric. Food Chem. 2005, 53, 28–31. [Google Scholar] [CrossRef]
- Johnson, M.H.; de Mejia, E.G. Phenolic Compounds from Fermented Berry Beverages Modulated Gene and Protein Expression To Increase Insulin Secretion from Pancreatic β-Cells in Vitro. J. Agric. Food Chem. 2016, 64, 2569–2581. [Google Scholar] [CrossRef]
- Kamonpatana, K.; Giusti, M.M.; Chitchumroonchokchai, C.; MorenoCruz, M.; Riedl, K.M.; Kumar, P.; Failla, M.L. Susceptibility of Anthocyanins to Ex Vivo Degradation in Human Saliva. Food Chem. 2012, 135, 738–747. [Google Scholar] [CrossRef] [Green Version]
- Kianbakht, S.; Abasi, B.; Hashem Dabaghian, F. Improved Lipid Profile in Hyperlipidemic Patients Taking Vaccinium Arctostaphylos Fruit Hydroalcoholic Extract: A Randomized Double-Blind Placebo-Controlled Clinical Trial. Phytother. Res. 2013, 28, 432–436. [Google Scholar] [CrossRef]
- Kumar, A.; Bharti, S.K.; Kumar, A. Therapeutic Molecules against Type 2 Diabetes: What We Have and What Are We Expecting? Pharmacol. Rep. 2017, 69, 959–970. [Google Scholar] [CrossRef]
- Li, J.; Kang, M.-K.; Kim, J.-K.; Kim, J.-L.; Kang, S.-W.; Lim, S.S.; Kang, Y.-H. Purple Corn Anthocyanins Retard Diabetes-Associated Glomerulosclerosis in Mesangial Cells and Db/Db Mice. Eur. J. Nutr. 2011, 51, 961–973. [Google Scholar] [CrossRef]
- Lim, K.; Ma, M.; Dolan, K.D. Effects of Spray Drying on Antioxidant Capacity and Anthocyanidin Content of Blueberry By-Products. J. Food Sci. 2011, 76, H156–H164. [Google Scholar] [CrossRef]
- Luna-Vital, D.A.; de Mejia, E.G. Anthocyanins from Purple Corn Activate Free Fatty Acid-Receptor 1 and Glucokinase Enhancing in Vitro Insulin Secretion and Hepatic Glucose Uptake. PLoS ONE 2018, 13, e0200449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luna-Vital, D.; Weiss, M.; de Mejia, E.G. Anthocyanins from Purple Corn Ameliorated Tumor Necrosis Factor-α-Induced Inflammation and Insulin Resistance in 3T3-L1 Adipocytes via Activation of Insulin Signaling and Enhanced GLUT4 Translocation. Mol. Nutr. Food Res. 2017, 61, 1700362. [Google Scholar] [CrossRef] [PubMed]
- McGhie, T.K.; Walton, M.C. The Bioavailability and Absorption of Anthocyanins: Towards a Better Understanding. Mol. Nutr. Food Res. 2007, 51, 702–713. [Google Scholar] [CrossRef] [PubMed]
- Nash, R.J.; Kato, A.; Yu, C.-Y.; Fleet, G.W. Iminosugars as Therapeutic Agents: Recent Advances and Promising Trends. Future Med. Chem. 2011, 3, 1513–1521. [Google Scholar] [CrossRef] [PubMed]
- Noratto, G.D.; Chew, B.P.; Atienza, L.M. Red Raspberry (Rubus Idaeus L.) Intake Decreases Oxidative Stress in Obese Diabetic (Db/Db) Mice. Food Chem. 2017, 227, 305–314. [Google Scholar] [CrossRef]
- Park, E.; Edirisinghe, I.; Wei, H.; Vijayakumar, L.P.; Banaszewski, K.; Cappozzo, J.C.; Burton-Freeman, B. A Dose-Response Evaluation of Freeze-Dried Strawberries Independent of Fiber Content on Metabolic Indices in Abdominally Obese Individuals with Insulin Resistance in a Randomized, Single-Blinded, Diet-Controlled Crossover Trial. Mol. Nutr. Food Res. 2016, 60, 1099–1109. [Google Scholar] [CrossRef]
- Paulsmeyer, M.; Chatham, L.; Becker, T.; West, M.; West, L.; Juvik, J. Survey of Anthocyanin Composition and Concentration in Diverse Maize Germplasms. J. Agric. Food Chem. 2017, 65, 4341–4350. [Google Scholar] [CrossRef]
- Pimpão, R.C.; Ventura, M.R.; Ferreira, R.B.; Williamson, G.; Santos, C.N. Phenolic Sulfates as New and Highly Abundant Metabolites in Human Plasma after Ingestion of a Mixed Berry Fruit Purée. Br. J. Nutr. 2015, 113, 454–463. [Google Scholar] [CrossRef] [Green Version]
- Riaz, M.; Zia-Ul-Haq, M.; Saad, B. Biosynthesis and Stability of Anthocyanins. In Anthocyanins and Human Health: Biomolecular and Therapeutic Aspects; Springer: Berlin/Heidelberg, Germany, 2016; pp. 71–86. [Google Scholar] [CrossRef]
- Richter, C.K.; Skulas-Ray, A.C.; Gaugler, T.L.; Lambert, J.D.; Proctor, D.N.; Kris-Etherton, P.M. Incorporating Freeze-Dried Strawberry Powder into a High-Fat Meal Does Not Alter Postprandial Vascular Function or Blood Markers of Cardiovascular Disease Risk: A Randomized Controlled Trial. Am. J. Clin. Nutr. 2016, 105, 313–322. [Google Scholar] [CrossRef] [Green Version]
- Saikia, S.; Dutta, H.; Saikia, D.; Mahanta, C.L. Quality Characterisation and Estimation of Phytochemicals Content and Antioxidant Capacity of Aromatic Pigmented and Non-Pigmented Rice Varieties. Food Res. Int. 2012, 46, 334–340. [Google Scholar] [CrossRef]
- Moreno, Y.S.; Sanchez, G.S.; Hernandez, D.R.; Lobato, N.R. Characterization of Anthocyanin Extracts from Maize Kernels. J. Chromatogr. Sci. 2005, 43, 483–487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schell, J.; Betts, N.M.; Lyons, T.J.; Basu, A. Raspberries Improve Postprandial Glucose and Acute and Chronic Inflammation in Adults with Type 2 Diabetes. Ann. Nutr. Metab. 2019, 74, 165–174. [Google Scholar] [CrossRef] [PubMed]
- Silvester, A.J.; Aseer, K.R.; Yun, J.W. Dietary Polyphenols and Their Roles in Fat Browning. J. Nutr. Biochem. 2019, 64, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Soltani, R.; Hakimi, M.; Asgary, S.; Ghanadian, S.M.; Keshvari, M.; Sarrafzadegan, N. Evaluation of the Effects of Vaccinium Arctostaphylos L. Fruit Extract on Serum Lipids and Hs-CRP Levels and Oxidative Stress in Adult Patients with Hyperlipidemia: A Randomized, Double-Blind, Placebo-Controlled Clinical Trial. Evid. Based Complement. Altern. Med. 2014, 2014, 217451. [Google Scholar] [CrossRef] [Green Version]
- Stalmach, A.; Edwards, C.A.; Wightman, J.D.; Crozier, A. Gastrointestinal Stability and Bioavailability of (Poly)Phenolic Compounds Following Ingestion of Concord Grape Juice by Humans. Mol. Nutr. Food Res. 2012, 56, 497–509. [Google Scholar] [CrossRef]
- Suantawee, T.; Elazab, S.; Hsu, W.; Yao, S.; Cheng, H.; Adisakwattana, S. Cyanidin Stimulates Insulin Secretion and Pancreatic β-Cell Gene Expression through Activation of l-Type Voltage-Dependent Ca2+ Channels. Nutrients 2017, 9, 814. [Google Scholar] [CrossRef]
- Talagavadi, V.; Rapisarda, P.; Galvano, F.; Pelicci, P.; Giorgio, M. Cyanidin-3-O-β-Glucoside and Protocatechuic Acid Activate AMPK/MTOR/S6K Pathway and Improve Glucose Homeostasis in Mice. J. Funct. Foods 2016, 21, 338–348. [Google Scholar] [CrossRef]
- Talavéra, S.; Felgines, C.; Texier, O.; Besson, C.; Manach, C.; Lamaison, J.-L.; Rémésy, C. Anthocyanins Are Efficiently Absorbed from the Small Intestine in Rats. J. Nutr. 2004, 134, 2275–2279. [Google Scholar] [CrossRef]
- Tsujihata, Y.; Ito, R.; Suzuki, M.; Harada, A.; Negoro, N.; Yasuma, T.; Momose, Y.; Takeuchi, K. TAK-875, an Orally Available G Protein-Coupled Receptor 40/Free Fatty Acid Receptor 1 Agonist, Enhances Glucose-Dependent Insulin Secretion and Improves Both Postprandial and Fasting Hyperglycemia in Type 2 Diabetic Rats. J. Pharmacol. Exp. Ther. 2011, 339, 228–237. [Google Scholar] [CrossRef] [PubMed]
- Wahyuningsih, S.; Wulandari, L.; Wartono, M.W.; Munawaroh, H.; Ramelan, A.H. The Effect of PH and Color Stability of Anthocyanin on Food Colorant. IOP Conf. Ser. Mater. Sci. Eng. 2017, 193, 012047. [Google Scholar] [CrossRef] [Green Version]
- Williamson, G. Possible Effects of Dietary Polyphenols on Sugar Absorption and Digestion. Mol. Nutr. Food Res. 2013, 57, 48–57. [Google Scholar] [CrossRef] [PubMed]
- Xiao, D.; Zhu, L.; Edirisinghe, I.; Fareed, J.; Brailovsky, Y.; Burton-Freeman, B. Attenuation of Postmeal Metabolic Indices with Red Raspberries in Individuals at Risk for Diabetes: A Randomized Controlled Trial. Obesity 2019, 27, 542–550. [Google Scholar] [CrossRef] [PubMed]
- Yan, F.; Zhang, J.; Zhang, L.; Zheng, X. Mulberry Anthocyanin Extract Regulates Glucose Metabolism by Promotion of Glycogen Synthesis and Reduction of Gluconeogenesis in Human HepG2 Cells. Food Funct. 2016, 7, 425–433. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Ling, W.; Guo, H.; Song, F.; Ye, Q.; Zou, T.; Li, D.; Zhang, Y.; Li, G.; Xiao, Y.; et al. Anti-Inflammatory Effect of Purified Dietary Anthocyanin in Adults with Hypercholesterolemia: A Randomized Controlled Trial. Nutr. Metab. Cardiovasc. Dis. 2013, 23, 843–849. [Google Scholar] [CrossRef]
- Žilić, S.; Serpen, A.; Akıllıoğlu, G.; Gökmen, V.; Vančetović, J. Phenolic Compounds, Carotenoids, Anthocyanins, and Antioxidant Capacity of Colored Maize (Zea Mays L.) Kernels. J. Agric. Food Chem. 2012, 60, 1224–1231. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Author | Study Design | Population | Dosage | Study Duration | Main Findings |
|---|---|---|---|---|---|
| María S. Hershey 2020 [50] | Cross-sectional ECA | 249 men Age: 47 ± 7.6 | Range from 6.05 to 120.8 mg/per ration | 2 years | High intake of anthocyanins is associated with > HDL. |
| Zhu, 2013 [51] | Randomized, double-blind, parallel | 73 participants, per group Age: 40–65 | 320 mg/day | 24 weeks | Anthocyanin supplementation decreased TC, TG, LDL compared to placebo > HDL levels. |
| Soltani, 2014 (Soltani, [52] | Randomized, double-blind, parallel | 25 participants, per group Age: 48 ± 16 | 90 mg/per day | 4 weeks | Anthocyanin supplementation decreased TC, TG, LDL compared to placebo. >HDL levels. |
| Kianbakht 2013 [53] | Randomized, double-blind, parallel | 40 participants, per group Age: 51.3 ± 15.27 | 259.68 mg/per day | 2 months | Anthocyanin supplementation decreased TC, TG, LDL compared to placebo. >HDL levels. |
| Schell et al.,2019 [54] | Cross-sectional, randomized, controlled | 25 participants (5 men, 20 women) Age: 54 ± 4.2 Obese diabetic adults | 225 mg | 4 h. | Lower postprandial glucose levels were reported at 2 and 4 h with the anthocyanin group vs. control. IL6/TNF α decreased at postprandial 4 h. |
| Park et al., 2016 [55] | Cross-sectional, randomized, controlled, double-blind, 4 groups | 21 participants (5 men 16 women) Age: 39.8 ± 13.8. Obese adults with insulin resistance | 42, 88 and 155 mg | 6 h. | The 6-hour postprandial insulin concentration in the group with the highest beverage concentration was significantly reduced. |
| Richter et al., 2016 [56] | Cross-sectional, randomized, controlled, double-blind, | 30 participants (17 men 13 women) Age: 28 ± 2 Healthy, overweight, and obese adults | 163 mg | 4 h | The intervention did not improve TG, glucose, or insulin. |
| Abubakar et al., 2019 [57] | Cross-sectional, randomized, controlled, double-blind, | 25 male participants. Age: 49 ± 2 Adults: CVD 1–10% in 10 years | 150 mg | 4 h. | Hibiscus extract improves postprandial vascular function, useful in reducing endothelial dysfunction and cardiovascular risk. |
| Xiao et al., 2019 [58] | Cross-sectional, three-arm, single blind randomized, controlled trial | 21 participants (12 men, 9 women) Age: 34 ± 12 With IR | 73, 146 mg | 24 h | Significant reduction at 2 h in the area under the curve and a reduction in peak insulin and glucose at 2 h. |
| Castro-Acosta et al., 2017 [59] | Cross-sectional, randomized, controlled, double-blind, | 22 participants (13 men 9 women) Age: 45.4 Healthy | 150, 300 and 600 mg | 2 h | A reduction in postprandial blood glucose, insulin, and incretin secretion was observed. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Franco-San Sebastián, D.; Alaniz-Monreal, S.; Rabadán-Chávez, G.; Vázquez-Manjarrez, N.; Hernández-Ortega, M.; Gutiérrez-Salmeán, G. Anthocyanins: Potential Therapeutic Approaches towards Obesity and Diabetes Mellitus Type 2. Molecules 2023, 28, 1237. https://doi.org/10.3390/molecules28031237
Franco-San Sebastián D, Alaniz-Monreal S, Rabadán-Chávez G, Vázquez-Manjarrez N, Hernández-Ortega M, Gutiérrez-Salmeán G. Anthocyanins: Potential Therapeutic Approaches towards Obesity and Diabetes Mellitus Type 2. Molecules. 2023; 28(3):1237. https://doi.org/10.3390/molecules28031237
Chicago/Turabian StyleFranco-San Sebastián, Denise, Samary Alaniz-Monreal, Griselda Rabadán-Chávez, Natalia Vázquez-Manjarrez, Marcela Hernández-Ortega, and Gabriela Gutiérrez-Salmeán. 2023. "Anthocyanins: Potential Therapeutic Approaches towards Obesity and Diabetes Mellitus Type 2" Molecules 28, no. 3: 1237. https://doi.org/10.3390/molecules28031237
APA StyleFranco-San Sebastián, D., Alaniz-Monreal, S., Rabadán-Chávez, G., Vázquez-Manjarrez, N., Hernández-Ortega, M., & Gutiérrez-Salmeán, G. (2023). Anthocyanins: Potential Therapeutic Approaches towards Obesity and Diabetes Mellitus Type 2. Molecules, 28(3), 1237. https://doi.org/10.3390/molecules28031237