
Preparation, Structural Characterisation, and Bioactivities of Fructans: A Review

Abstract
:1. Introduction
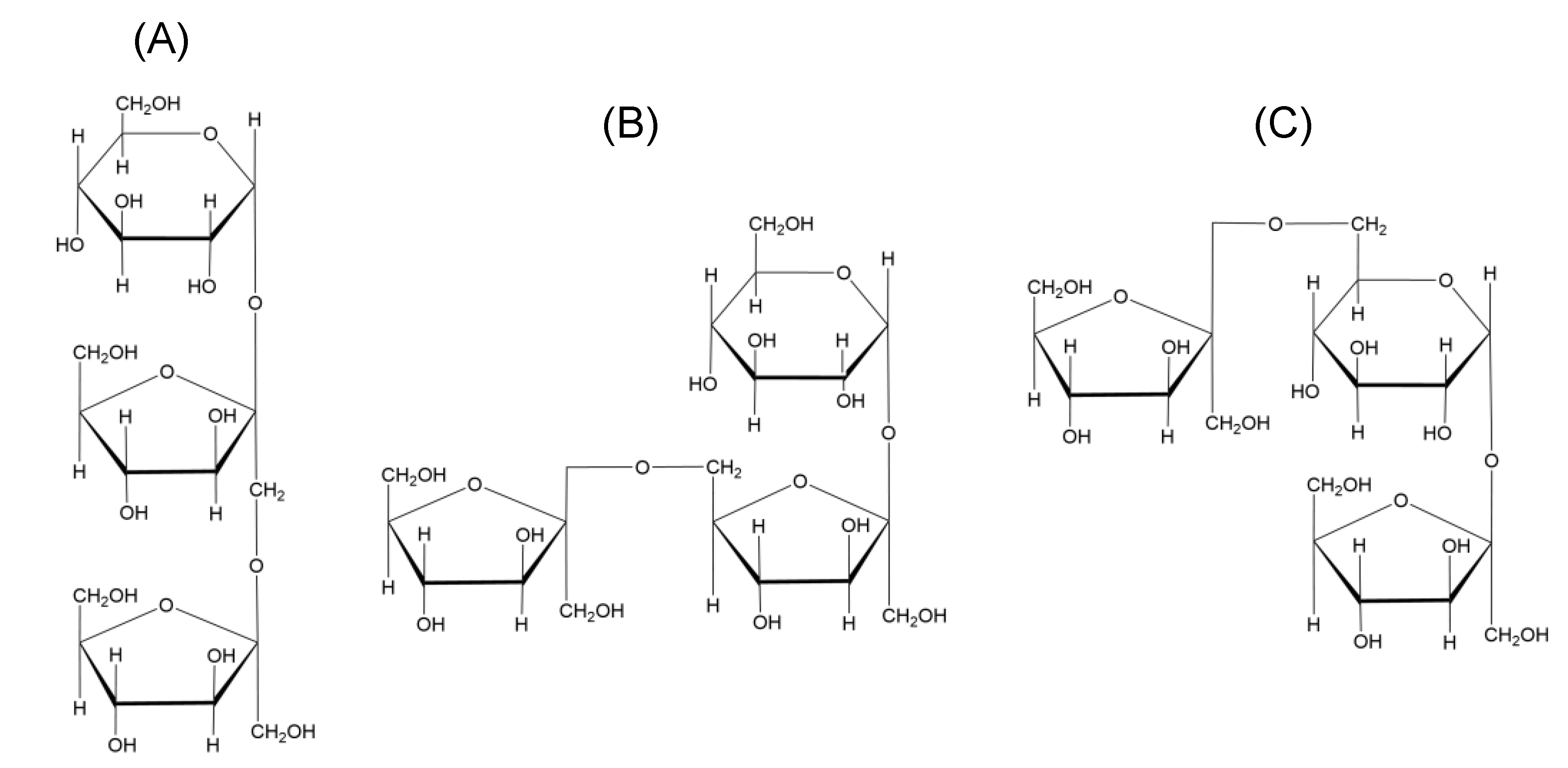
2. Occurrence and Chemistry in Plants
3. Metabolism in Plants
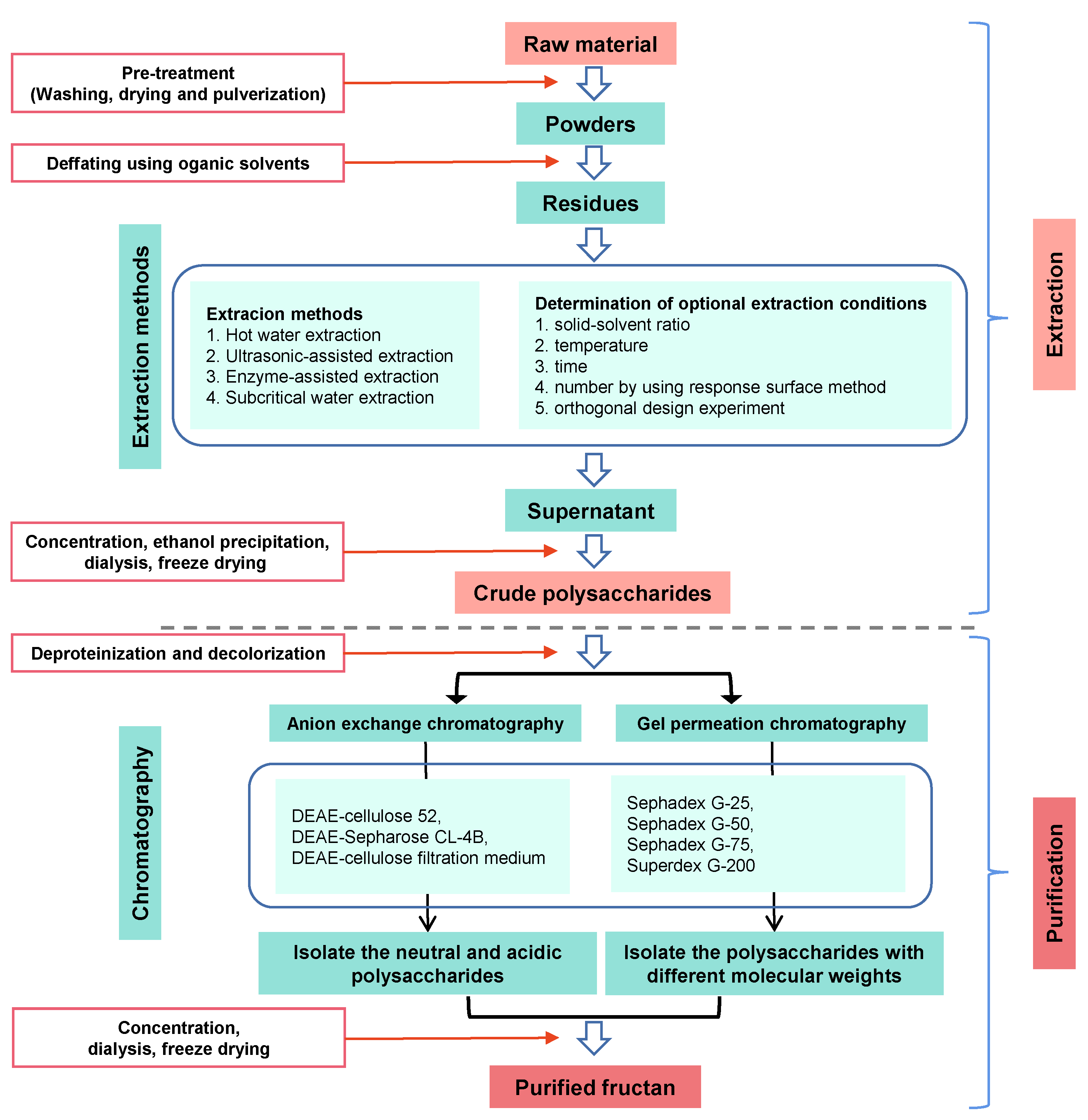
4. Extraction and Purification of Fructans
4.1. Extraction
4.2. Purification
5. Characterisation
5.1. MW

{kind=link}
{kind=link}
{kind=link}
| Source | Extraction Procedure | Structural Characteristic | Biological Activity | Ref. |
|---|---|---|---|---|
| Agave | Mixture of β(2→1) and β(2→6)-Fruf | Antioxidant | [42] | |
| Agave tequilana | Commercial product | Mixture of β(2→1) and β(2→6)-Fruf, DP 7-45 | Antiinflammation | [43] |
| Anemarrhena asphodeloides | Hot water (80 °C) extraction | Backbone (2→6)-linked β-D-Fruf, MW 2.72 kDa | Neuroprotective and immunoregulatory | [39] |
| Atractylodes chinensis | Enzymatic auxiliary-ultrasonic extraction | Backbone (2→1)-linked β-D-Fruf, MW 11.2 kDa | Antitumor | [44] |
| Atractylodes macrocephala | Hot water (80 °C) extraction | α-D-Glcp-(1→(2-β-D-Fruf-1)7 | Anti-weightlessness bone loss | [45] |
| Atractylodis macrocephalae | 0.2 mol/L NaOH (100 °C) extraction | Backbone (2→1)-linked β-D-Fruf, MW 3.438 kDa | Immunoregulatory | [46] |
| Asparagus | Hot water (80 °C) extraction | Backbone (2→1)-linked β-D-Fruf, DP > 25 | Prebiotic | [47] |
| Asparagus cochinchinensis | Hot water extraction | Backbone (2→1)-linked β-D-Fruf, MW 2.69 kDa | Regulates gut microbiota | [48] |
| Codonopsis pilosula | Hot water (85 °C) extraction | Backbone (2→1)-linked β-D-Fruf, MW 3.6 kDa | Anti-gastric ulcer | [49] |
| Codonopsis pilosula | Ultrasonic extraction (90 °C) | Backbone (2→1)-linked β-D-Fruf, DP 16-31 | Prebiotic | [41] |
| Codonopsis tangshen | Hot water extraction | Backbone (2→1)-linked β-D-Fruf, MW 3.95 kD | Antioxidant and prebiotic | [50] |
| Jerusalem artichoke | Hot water (70 °C) extraction | Backbone (2→1)-linked β-D-Fruf, MW 2.6 kD | Antitumor | [51] |
| Lobelia chinensis | Hot water extraction | α-D-Glcp-(1→(1-β-D-Fruf-2)15 linkage, MW 2.6 kDa | Antiobesity | [52] |
| Platycodon grandiflorum | Hot water (90 °C) extraction | Backbone (2→1)-linked β-D-Fruf, MW 12.1 kDa | Regulates gut microbiota | [53] |
| Platycodon grandiflorus | Hot water extraction | Backbone (2→1)-linked β-D-Fruf, DP 2–7 | Prebiotic and immunoregulatory | [37] |
5.2. Monosaccharide Composition
5.3. Quantification
5.4. Linkage Analysis
5.5. Structural Analysis
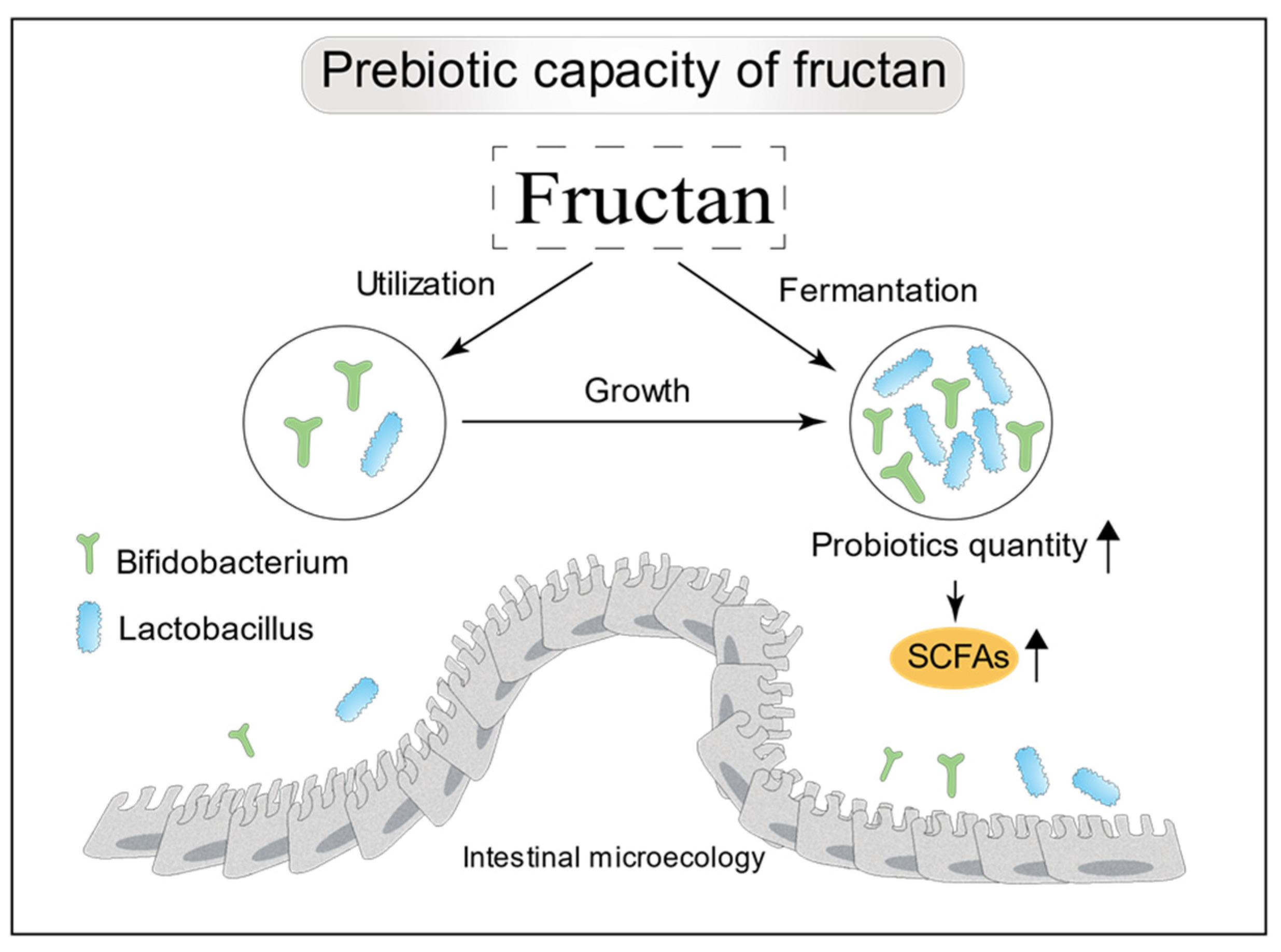
6. Bioactivity
7. Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Aldrete-Herrera, P.I.; López, M.G.; Medina-Torres, L.; Ragazzo-Sánchez, J.A.; Calderón-Santoyo, M.; González-Ávila, M.; Ortiz-Basurto, R.I. Physicochemical composition and apparent degree of polymerization of fructans in five wild agave varieties: Potential industrial use. Foods 2019, 8, 404. [Google Scholar] [CrossRef] [PubMed]
- Meng, Y.; Xu, Y.; Chang, C.; Qiu, Z.; Hu, J.; Wu, Y.; Zhang, B.; Zheng, G. Extraction, characterization and anti-inflammatory activities of an inulin-type fructan from Codonopsis pilosula. Int. J. Biol. Macromol. 2020, 163, 1677–1686. [Google Scholar] [CrossRef] [PubMed]
- Espinosa-Andrews, H.; Urías-Silvas, J.E.; Morales-Hernández, N. The role of agave fructans in health and food applications: A review. Trends Food Sci. Technol. 2021, 114, 585–598. [Google Scholar] [CrossRef]
- Veenstra, L.D.; Santantonio, N.; Jannink, J.-L.; Sorrells, M.E. Influence of genotype and environment on wheat grain fructan content. Crop Sci. 2019, 59, 190–198. [Google Scholar] [CrossRef]
- Van Laere, A.; Van Den Ende, W. Inulin metabolism in dicots: Chicory as a model system. Plant Cell Environ. 2002, 25, 803–813. [Google Scholar] [CrossRef]
- Jirayucharoensak, R.; Khuenpet, K.; Jittanit, W.; Sirisansaneeyakul, S. Physical and chemical properties of powder produced from spray drying of inulin component extracted from Jerusalem artichoke tuber powder. Dry. Technol. 2019, 37, 1215–1227. [Google Scholar] [CrossRef]
- Chikkerur, J.; Samanta, A.K.; Kolte, A.P.; Dhali, A.; Roy, S. Production of short chain fructo-oligosaccharides from inulin of chicory root using fungal endoinulinase. Appl. Biochem. Biotechnol. 2020, 191, 695–715. [Google Scholar] [CrossRef]
- Aisara, J.; Wongputtisin, P.; Deejing, S.; Maneewong, C.; Unban, K.; Khanongnuch, C.; Kosma, P.; Blaukopf, M.; Kanpiengjai, A. Potential of iInulin-fructooligosaccharides extract produced from red onion (Allium cepa var. viviparum (Metz) Mansf.) as an alternative prebiotic product. Plants 2021, 10, 2401. [Google Scholar] [CrossRef]
- Viera-Alcaide, I.; Hamdi, A.; Guillén-Bejarano, R.; Rodríguez-Arcos, R.; Espejo-Calvo, J.A.; Jiménez-Araujo, A. Asparagus roots: From an agricultural by-product to a valuable source of fructans. Foods 2022, 11, 652. [Google Scholar] [CrossRef]
- Apolinário, A.C.; de Carvalho, E.M.; de Lima Damasceno, B.P.G.; da Silva, P.C.D.; Converti, A.; Pessoa, A.; da Silva, J.A. Extraction, isolation and characterization of inulin from Agave sisalana boles. Ind. Crop. Prod. 2017, 108, 355–362. [Google Scholar] [CrossRef]
- Verspreet, J.; Hansen, A.H.; Harrison, S.J.; Vergauwen, R.; Van den Ende, W.; Courtin, C.M. Building a fructan LC–MS2 library and its application to reveal the fine structure of cereal grain fructans. Carbohydr. Polym. 2017, 174, 343–351. [Google Scholar] [CrossRef]
- Verspreet, J.; Hansen, A.H.; Dornez, E.; Delcour, J.A.; Van den Ende, W.; Harrison, S.J.; Courtin, C.M. LC-MS analysis reveals the presence of graminan- and neo-type fructans in wheat grains. J. Cereal Sci. 2015, 61, 133–138. [Google Scholar] [CrossRef]
- Szwengiel, A.; Nkongha, G.L. Influence of acid depolymerization parameters on levan molar mass distribution and its utilization by bacteria. Carbohydr. Polym. 2019, 206, 371–379. [Google Scholar] [CrossRef]
- Ispiryan, L.; Kuktaite, R.; Zannini, E.; Arendt, E.K. Fundamental study on changes in the FODMAP profile of cereals, pseudo-cereals, and pulses during the malting process. Food Chem. 2021, 343, 128549. [Google Scholar] [CrossRef]
- Mudannayake, D.C.; Jayasena, D.D.; Wimalasiri, K.M.S.; Ranadheera, C.S.; Ajlouni, S. Inulin fructans—food applications and alternative plant sources: A review. Int. J. Food Sci. Technol. 2022, 57, 5764–5780. [Google Scholar] [CrossRef]
- Muir, J.G.; Shepherd, S.J.; Rosella, O.; Rose, R.; Barrett, J.S.; Gibson, P.R. Fructan and free fructose content of common Australian vegetables and fruit. J. Agric. Food. Chem. 2007, 55, 6619–6627. [Google Scholar] [CrossRef]
- Dunn, S.; Datta, A.; Kallis, S.; Law, E.; Myers, C.E.; Whelan, K. Validation of a food frequency questionnaire to measure intakes of inulin and oligofructose. Eur. J. Clin. Nutr. 2011, 65, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Ueno, K.; Sonoda, T.; Yoshida, M.; Kawakami, A.; Shiomi, N.; Onodera, S. Decreased expression of fructosyltransferase genes in asparagus roots may contribute to efficient fructan degradation during asparagus spear harvesting. Plant Physiol. Biochem. 2020, 156, 192–200. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, G.; Zhou, Z.; Zong, Y.; Wang, L.; Yang, S.; Zhang, Y.; Sun, X. Transcriptome analysis for genes involved in fructan biosynthesis in the Jerusalem artichoke (Helianthus tuberosus L.). Biotechnol. Biotec. Eq. 2022, 36, 472–480. [Google Scholar] [CrossRef]
- Márquez-López, R.E.; Loyola-Vargas, V.M.; Santiago-García, P.A. Interaction between fructan metabolism and plant growth regulators. Planta 2022, 255, 49. [Google Scholar] [CrossRef]
- Ueno, K.; Sonoda, T.; Yoshida, M.; Shiomi, N.; Onodera, S. Purification, characterization, and functional analysis of a novel 6G&1-FEH mainly hydrolyzing neokestose from asparagus. J. Exp. Bot. 2018, 69, 4295–4308. [Google Scholar] [PubMed]
- Van den Ende, W. Different evolutionary pathways to generate plant fructan exohydrolases. J. Exp. Bot. 2022, 73, 4620–4623. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.-H.; Xiong, W.-P.; Huang, C.; Li, X.-M.; Wang, Q.-Y.; Huang, R.-B. Engineering better catalytic activity and acidic adaptation into Kluyveromyces marxianus exoinulinase using site-directed mutagenesis. J. Sci. Food Agric. 2021, 101, 2472–2482. [Google Scholar] [CrossRef] [PubMed]
- Rigui, A.P.; Gaspar, M.; Oliveira, V.F.; Purgatto, E.; Carvalho, M.A.M.d. Endogenous hormone concentrations correlate with fructan metabolism throughout the phenological cycle in Chrysolaena obovata. Ann. Bot. 2015, 115, 1163–1175. [Google Scholar] [CrossRef] [PubMed]
- Portes, M.T.; Figueiredo-Ribeiro, R.d.C.L.; de Carvalho, M.A.M. Low temperature and defoliation affect fructan-metabolizing enzymes in different regions of the rhizophores of Vernonia herbacea. J. Plant Physiol. 2008, 165, 1572–1581. [Google Scholar] [CrossRef]
- Abeynayake, S.W.; Etzerodt, T.P.; Jonavičienė, K.; Byrne, S.; Asp, T.; Boelt, B. Fructan metabolism and changes in fructan composition during cold acclimation in perennial ryegrass. Front. Plant Sci. 2015, 6, 329. [Google Scholar] [CrossRef]
- Zeaiter, Z.; Regonesi, M.E.; Cavini, S.; Labra, M.; Sello, G.; Di Gennaro, P. Extraction and characterization of inulin-type fructans from Artichoke wastes and their effect on the growth of intestinal bacteria associated with health. Biomed Res. Int. 2019, 2019, 1083952. [Google Scholar] [CrossRef]
- Fu, Y.-P.; Li, L.-X.; Zhang, B.-Z.; Paulsen, B.S.; Yin, Z.-Q.; Huang, C.; Feng, B.; Chen, X.-F.; Jia, R.-R.; Song, X.; et al. Characterization and prebiotic activity in vitro of inulin-type fructan from Codonopsis pilosula roots. Carbohydr. Polym. 2018, 193, 212–220. [Google Scholar] [CrossRef]
- Lu, X.; Li, N.; Qiao, X.; Qiu, Z.; Liu, P. Effects of thermal treatment on polysaccharide degradation during black garlic processing. LWT 2018, 95, 223–229. [Google Scholar] [CrossRef]
- Petkova, N.; Ivanov, I.; Vrancheva, R.; Denev, P.; Pavlov, A. Ultrasound and microwave-assisted extraction of Elecampane (Inula helenium) Roots. Nat. Prod. Commun. 2017, 12, 1934578X1701200207. [Google Scholar] [CrossRef]
- Shalini, R.; Krishna, J.; Sankaranarayanan, M.; Antony, U. Enhancement of fructan extraction from garlic and fructooligosaccharide purification using an activated charcoal column. LWT 2021, 148, 111703. [Google Scholar]
- Wang, H.; Ma, J.-X.; Zhou, M.; Si, J.; Cui, B.-K. Current advances and potential trends of the polysaccharides derived from medicinal mushrooms sanghuang. Front. Microbiol. 2022, 13, 2762. [Google Scholar] [CrossRef]
- Luiz-Santos, N.; Prado-Ramírez, R.; Arriola-Guevara, E.; Camacho-Ruiz, R.-M.; Moreno-Vilet, L. Performance evaluation of tight ultrafiltration membrane systems at pilot scale for agave fructans fractionation and purification. Membranes 2020, 10, 261. [Google Scholar] [CrossRef]
- Ostermann-Porcel, M.V.; Rinaldoni, A.N.; Campderrós, M.E. Assessment of Jerusalem artichoke as a source for the production of gluten-free flour and fructan concentrate by ultrafiltration. Appl. Food Res. 2022, 2, 100201. [Google Scholar] [CrossRef]
- Sun, P.; Liao, S.-G.; Yang, R.-Q.; Lu, C.-L.; Ji, K.-L.; Cao, D.-H.; Hu, H.-B.; Lu, J.-M.; Song, X.-Z.; Wu, M.; et al. Aspidopterys obcordata vine inulin fructan affects urolithiasis by modifying calcium oxalate crystallization. Carbohydr. Polym. 2022, 294, 119777. [Google Scholar] [CrossRef]
- Young, I.D.; Nepogodiev, S.A.; Black, I.M.; Le Gall, G.; Wittmann, A.; Latousakis, D.; Visnapuu, T.; Azadi, P.; Field, R.A.; Juge, N.; et al. Lipopolysaccharide associated with β-2,6 fructan mediates TLR4-dependent immunomodulatory activity in vitro. Carbohydr. Polym. 2022, 277, 118606. [Google Scholar] [CrossRef]
- Pang, D.-J.; Huang, C.; Chen, M.-L.; Chen, Y.-L.; Fu, Y.-P.; Paulsen, B.S.; Rise, F.; Zhang, B.-Z.; Chen, Z.-L.; Jia, R.-Y.; et al. Characterization of inulin-type fructan from Platycodon grandiflorus and study on Its prebiotic and immunomodulating activity. Molecules 2019, 24, 1199. [Google Scholar] [CrossRef]
- Zhang, Y.-y.; Zhuang, D.; Wang, H.-y.; Liu, C.-y.; Lv, G.-p.; Meng, L.-j. Preparation, characterization, and bioactivity evaluation of oligosaccharides from Atractylodes lancea (Thunb.) DC. Carbohydr. Polym. 2022, 277, 118854. [Google Scholar] [CrossRef]
- Zhang, S.; Zhang, Q.; An, L.; Zhang, J.; Li, Z.; Zhang, J.; Li, Y.; Tuerhong, M.; Ohizumi, Y.; Jin, J.; et al. A fructan from Anemarrhena asphodeloides Bunge showing neuroprotective and immunoregulatory effects. Carbohydr. Polym. 2020, 229, 115477. [Google Scholar] [CrossRef]
- Gan, L.; Jiang, G.; Li, X.; Zhang, S.; Tian, Y.; Peng, B. Structural elucidation and physicochemical characteristics of a novel high-molecular-weight fructan from halotolerant Bacillus sp. SCU-E108. Food Chem. 2021, 365, 130496. [Google Scholar] [CrossRef]
- Li, J.; Zhang, X.; Cao, L.; Ji, J.; Gao, J. Three inulin-type fructans from Codonopsis pilosula (Franch.) Nannf. Roots and their prebiotic activity on Bifidobacterium longum. Molecules 2018, 23, 3123. [Google Scholar] [CrossRef] [PubMed]
- Franco-Robles, E.; Ramírez-Emiliano, J.; López, M.G. Agave fructans and oligofructose decrease oxidative stress in brain regions involved in learning and memory of overweight mice. Nat. Prod. Res. 2019, 33, 1527–1530. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Lainez, C.; Akkerman, R.; Oerlemans, M.M.P.; Logtenberg, M.J.; Schols, H.A.; Silva-Lagos, L.A.; López-Velázquez, G.; de Vos, P. β(2→6)-Type fructans attenuate proinflammatory responses in a structure dependent fashion via Toll-like receptors. Carbohydr. Polym. 2022, 277, 118893. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Chen, D.; Liu, C.; Wu, X.-Z.; Dong, C.-X.; Zhou, J. Structural characterization and anti-tumor effects of an inulin-type fructan from Atractylodes chinensis. Int. J. Biol. Macromol. 2016, 82, 765–771. [Google Scholar] [CrossRef]
- Wang, R.; Shan, H.; Zhang, G.; Li, Q.; Wang, J.; Yan, Q.; Li, E.; Diao, Y.; Wei, L. An inulin-type fructan (AMP1-1) from Atractylodes macrocephala with anti-weightlessness bone loss activity. Carbohydr. Polym. 2022, 294, 119742. [Google Scholar] [CrossRef]
- Li, X.-J.; Xiao, S.-J.; Chen, J.; Xu, H.-R. Inulin-type fructans obtained from Atractylodis Macrocephalae by water/alkali extraction and immunoregulatory evaluation. Int. J. Biol. Macromol. 2023, 230, 123212. [Google Scholar] [CrossRef]
- Hamdi, A.; Viera-Alcaide, I.; Guillén-Bejarano, R.; Rodríguez-Arcos, R.; Muñoz, M.J.; Moreno, J.M.M.; Jiménez-Araujo, A. Asparagus fructans as emerging prebiotics. Foods 2023, 12, 81. [Google Scholar] [CrossRef]
- Sun, Q.; Zhu, L.; Li, Y.; Cui, Y.; Jiang, S.; Tao, N.; Chen, H.; Zhao, Z.; Xu, J.; Dong, C. A novel inulin-type fructan from Asparagus cochinchinensis and its beneficial impact on human intestinal microbiota. Carbohydr. Polym. 2020, 247, 116761. [Google Scholar] [CrossRef]
- Li, J.; Wang, T.; Zhu, Z.; Yang, F.; Cao, L.; Gao, J. Structure features and anti-gastric ulcer effects of inulin-type fructan CP-A from the roots of Codonopsis pilosula (Franch.) Nannf. Molecules 2017, 22, 2258. [Google Scholar] [CrossRef]
- Zou, Y.-F.; Zhang, Y.-Y.; Zhu, Z.-K.; Fu, Y.-P.; Paulsen, B.S.; Huang, C.; Feng, B.; Li, L.-X.; Chen, X.-F.; Jia, R.-Y.; et al. Characterization of inulin-type fructans from two species of Radix Codonopsis and their oxidative defense activation and prebiotic activities. J. Sci. Food Agric. 2021, 101, 2491–2499. [Google Scholar] [CrossRef]
- Shao, T.; Yuan, P.; Zhang, W.; Dou, D.; Wang, F.; Hao, C.; Liu, C.; Han, J.; Chen, K.; Wang, G. Preparation and characterization of sulfated inulin-type fructans from Jerusalem artichoke tubers and their antitumor activity. Carbohydr. Res. 2021, 509, 108422. [Google Scholar] [CrossRef]
- Zhang, X.; Hu, P.; Zhang, X.; Li, X. Chemical structure elucidation of an inulin-type fructan isolated from Lobelia chinensis lour with anti-obesity activity on diet-induced mice. Carbohydr. Polym. 2020, 240, 116357. [Google Scholar] [CrossRef]
- Shan, S.; Xiong, Y.; Guo, J.; Liu, M.; Gao, X.; Fu, X.; Zeng, D.; Song, C.; Zhang, Y.; Cheng, D.; et al. Effect of an inulin-type fructan from Platycodon grandiflorum on the intestinal microbiota in rats exposed to PM2.5. Carbohydr. Polym. 2022, 283, 119147. [Google Scholar] [CrossRef]
- Ji, X.; Hou, C.; Shi, M.; Yan, Y.; Liu, Y. An insight into the research concerning Panax ginseng C. A. Meyer polysaccharides: A review. Food Rev. Int. 2022, 38, 1149–1165. [Google Scholar] [CrossRef]
- Rodríguez-Rodríguez, R.; Barajas-Álvarez, P.; Morales-Hernández, N.; Camacho-Ruíz, R.M.; Espinosa-Andrews, H. Physical properties and prebiotic activities (Lactobacillus spp.) of gelatine-based gels formulated with agave fructans and agave syrups as sucrose and glucose substitutes. Molecules 2022, 27, 4902. [Google Scholar] [CrossRef]
- Lopes, S.M.S.; Francisco, M.G.; Higashi, B.; de Almeida, R.T.R.; Krausová, G.; Pilau, E.J.; Gonçalves, J.E.; Gonçalves, R.A.C.; Oliveira, A.J.B.d. Chemical characterization and prebiotic activity of fructo-oligosaccharides from Stevia rebaudiana (Bertoni) roots and in vitro adventitious root cultures. Carbohydr. Polym. 2016, 152, 718–725. [Google Scholar] [CrossRef]
- Ji, X.; Peng, B.; Ding, H.; Cui, B.; Nie, H.; Yan, Y. Purification, structure and biological activity of pumpkin polysaccharides: A review. Food Rev. Int. 2021. [Google Scholar] [CrossRef]
- Ruiz-Matute, A.I.; Hernadnez-Hernandez, O.; Rodriguez-Sanchez, S.; Sanz, M.L.; Martinz-Castro, I. Derivatization of carbohydrates for GC and GC-MS analyses. J. Chromatogr. B 2011, 879, 1226–1240. [Google Scholar] [CrossRef]
- Ispiryan, L.; Heitmann, M.; Hoehnel, A.; Zannini, E.; Arendt, E.K. Optimization and validation of an HPAEC-PAD method for the quantification of FODMAPs in cereals and cereal-based products. J. Agric. Food. Chem. 2019, 67, 4384–4392. [Google Scholar] [CrossRef]
- Tobaruela, E.d.C.; Santos, A.d.O.; Almeida-Muradian, L.B.d.; Araujo, E.d.S.; Lajolo, F.M.; Menezes, E.W. Application of dietary fiber method AOAC 2011.25 in fruit and comparison with AOAC 991.43 method. Food Chem. 2018, 238, 87–93. [Google Scholar] [CrossRef]
- Zhang, J.; Chen, H.; Luo, L.; Zhou, Z.; Wang, Y.; Gao, T.; Yang, L.; Peng, T.; Wu, M. Structures of fructan and galactan from Polygonatum cyrtonema and their utilization by probiotic bacteria. Carbohydr. Polym. 2021, 267, 118219. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Shi, C.; Shi, S.; Wang, H.; Yan, J.; Wang, S. An inulin-type fructan isolated from Artemisia japonica and its anti-arthritic effects. J. Funct. Food. 2017, 29, 29–36. [Google Scholar] [CrossRef]
- Li, J.; Wang, Y.; Zhang, X.; Cao, L.; Ji, J.; Zheng, Q.; Gao, J. Isolation and structural identification of a novel fructan from Radix Codonopsis. J. Carbohydr. Chem. 2020, 39, 163–174. [Google Scholar] [CrossRef]
- De Vries, J.; Le Bourgot, C.; Calame, W.; Respondek, F. Effects of β-fructans fiber on bowel function: A systematic review and meta-analysis. Nutrients 2019, 11, 91. [Google Scholar] [CrossRef]
- Healey, G.; Murphy, R.; Butts, C.; Brough, L.; Whelan, K.; Coad, J. Habitual dietary fibre intake influences gut microbiota response to an inulin-type fructan prebiotic: A randomised, double-blind, placebo-controlled, cross-over, human intervention study. Br. J. Nutr. 2018, 119, 176–189. [Google Scholar] [CrossRef]
- Zhang, J.; Li, Z.; Zhou, L.; Bao, J.; Xu, J. The modifications of a fructan from Anemarrhena asphodeloides Bunge and their antioxidant activities. Int. J. Biol. Macromol. 2020, 164, 4435–4443. [Google Scholar] [CrossRef]
- Caleffi, E.R.; Krausová, G.; Hyršlová, I.; Paredes, L.L.R.; dos Santos, M.M.; Sassaki, G.L.; Gonçalves, R.A.C.; de Oliveira, A.J.B. Isolation and prebiotic activity of inulin-type fructan extracted from Pfaffia glomerata (Spreng) Pedersen roots. Int. J. Biol. Macromol. 2015, 80, 392–399. [Google Scholar] [CrossRef]
- Chen, B.; Shi, Y.; Zhang, K.; Chang, Y.; Fu, P.; Liu, P.; Zhang, S. Inulin reduces liver triacylglycerol by increasing lipid droplet lipolysis in fat-loaded mice. Food Res. Int. 2023, 163, 112226. [Google Scholar] [CrossRef]
- Li, T.; Hou, X.; Huang, Y.; Wang, C.; Chen, H.; Yan, C. In vitro and in silico anti-osteoporosis activities and underlying mechanisms of a fructan, ABW90-1, from Achyranthes bidentate. Carbohydr. Polym. 2022, 276, 118730. [Google Scholar] [CrossRef]
- Ojwach, J.; Adetunji, A.I.; Mutanda, T.; Mukaratirwa, S. Oligosaccharides production from coprophilous fungi: An emerging functional food with potential health-promoting properties. Biotechnol. Rep. 2022, 33, e00702. [Google Scholar] [CrossRef]
- Rivière, A.; Selak, M.; Geirnaert, A.; Abbeele, P.V.d.; Vuyst, L.D. Complementary mechanisms for degradation of inulin-type fructans and arabinoxylan oligosaccharides among Bifidobacterial strains suggest bacterial cooperation. Appl. Environ. Microbiol. 2018, 84, e02893-17. [Google Scholar] [CrossRef]
- Deroover, L.; Verspreet, J.; Luypaerts, A.; Vandermeulen, G.; Courtin, C.M.; Verbeke, K. Wheat bran does not affect postprandial plasma short-chain fatty acids from 13C-inulin fermentation in healthy subjects. Nutrients 2017, 9, 83. [Google Scholar] [CrossRef]
- Belobrajdic, D.P.; Jenkins, C.L.D.; Christophersen, C.T.; Bird, A.R. Cereal fructan extracts alter intestinal fermentation to reduce adiposity and increase mineral retention compared to oligofructose. Eur. J. Nutr. 2019, 58, 2811–2821. [Google Scholar] [CrossRef]
- Bouhnik, Y.; Vahedi, K.; Achour, L.; Attar, A.; Salfati, J.; Pochart, P.; Marteau, P.; Flouria, B.; Bornet, F.; Rambaud, J.-C. Short-chain fructo-oligosaccharide administration dose-dependently increases fecal bifidobacteria in healthy humans. J. Nutr. 1999, 129, 113–116. [Google Scholar] [CrossRef]
- Zheng, L.-X.; Chen, X.-Q.; Cheong, K.-L. Current trends in marine algae polysaccharides: The digestive tract, microbial catabolism, and prebiotic potential. Int. J. Biol. Macromol. 2020, 151, 344–354. [Google Scholar] [CrossRef]
- Liu, F.; Prabhakar, M.; Ju, J.; Long, H.; Zhou, H.W. Effect of inulin-type fructans on blood lipid profile and glucose level: A systematic review and meta-analysis of randomized controlled trials. Eur. J. Clin. Nutr. 2017, 71, 9–20. [Google Scholar] [CrossRef]
- Salto, R.; Girón, M.D.; Ortiz-Moral, C.; Manzano, M.; Vílchez, J.D.; Reche-Perez, F.J.; Bueno-Vargas, P.; Rueda, R.; Lopez-Pedrosa, J.M. Dietary complex and slow digestive carbohydrates prevent fat deposits during catch-up growth in rats. Nutrients 2020, 12, 2568. [Google Scholar] [CrossRef]
- Cheong, K.-L.; Yu, B.; Chen, J.; Zhong, S. A comprehensive review of the cardioprotective effect of marine algae polysaccharide on the gut microbiota. Foods 2022, 11, 3550. [Google Scholar] [CrossRef]
- Forman, H.J.; Zhang, H. Targeting oxidative stress in disease: Promise and limitations of antioxidant therapy. Nat. Rev. Drug Discov. 2021, 20, 689–709. [Google Scholar] [CrossRef]
- Hu, Y.-C.; Hu, J.-L.; Li, J.; Wang, J.; Zhang, X.-Y.; Wu, X.-Y.; Li, X.; Guo, Z.-B.; Zou, L.; Wu, D.-T. Physicochemical characteristics and biological activities of soluble dietary fibers isolated from the leaves of different quinoa cultivars. Food Res. Int. 2023, 163, 112166. [Google Scholar] [CrossRef]
- Liu, W.; Wang, J.; Zhang, Z.; Xu, J.; Xie, Z.; Slavin, M.; Gao, X. In vitro and in vivo antioxidant activity of a fructan from the roots of Arctium lappa L. Int. J. Biol. Macromol. 2014, 65, 446–453. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Qiu, Z.; Dong, H.; Ma, C.; Qiao, Y.; Zheng, Z. Structural characterization and antioxidant activities of one neutral polysaccharide and three acid polysaccharides from the roots of Arctium lappa L.: A comparison. Int. J. Biol. Macromol. 2021, 182, 187–196. [Google Scholar] [CrossRef] [PubMed]
- Yao, W.; Qiu, H.-M.; Cheong, K.-L.; Zhong, S. Advances in anti-cancer effects and underlying mechanisms of marine algae polysaccharides. Int. J. Biol. Macromol. 2022, 221, 472–485. [Google Scholar] [CrossRef] [PubMed]
- Han, K.; Nam, J.; Xu, J.; Sun, X.; Huang, X.; Animasahun, O.; Achreja, A.; Jeon, J.H.; Pursley, B.; Kamada, N.; et al. Generation of systemic antitumour immunity via the in situ modulation of the gut microbiome by an orally administered inulin gel. Nat. Biomed. Eng. 2021, 5, 1377–1388. [Google Scholar] [CrossRef]
- Mazraeh, R.; Azizi-Soleiman, F.; Jazayeri, S.; Noori, S.M.A. Effect of inulin-type fructans in patients undergoing cancer treatments: A systematic review. Pak. J. Med. Sci. 2019, 35, 575–580. [Google Scholar] [CrossRef]



| Type of Fructan | Linear Bond | Formula | Source | Type of Kestose |
|---|---|---|---|---|
| Linear inulin | β(2→1) | G1–2F1–2Fn | Jerusalem artichoke [6], chicory [7] | 1-kestose |
| Inulin neoseries | β(2→1) | mF2–1F2–6G1-2F1–2Fn | Onion [8], asparagus [9], agave [10] | 6G-kestose |
| Linear levan | β(2→6) | G1–2F6–2Fn | Rye grass [11] | 6-kestose |
| Mixed levan | β(2→1) β(2→6) | G1–2F1 (6–2)F m–2F | Wheat [12], rye, barley [13] | 1-kestose 6-kestose |
| Levan neoseries | β(2→1) β(2→6) | F2(6–2)Fm–6G1–2F1 (6–2)F n–2F | Oat [14] | 6G-kestose |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, M.; Cheong, K.-L. Preparation, Structural Characterisation, and Bioactivities of Fructans: A Review. Molecules 2023, 28, 1613. https://doi.org/10.3390/molecules28041613
Wang M, Cheong K-L. Preparation, Structural Characterisation, and Bioactivities of Fructans: A Review. Molecules. 2023; 28(4):1613. https://doi.org/10.3390/molecules28041613
Chicago/Turabian StyleWang, Min, and Kit-Leong Cheong. 2023. "Preparation, Structural Characterisation, and Bioactivities of Fructans: A Review" Molecules 28, no. 4: 1613. https://doi.org/10.3390/molecules28041613
APA StyleWang, M., & Cheong, K. -L. (2023). Preparation, Structural Characterisation, and Bioactivities of Fructans: A Review. Molecules, 28(4), 1613. https://doi.org/10.3390/molecules28041613