

LncRNA JHDM1D-AS1 Is a Key Biomarker for Progression and Modulation of Gemcitabine Sensitivity in Bladder Cancer Cells

 , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. JHDM1D-AS1 and JHDM1D Expression in Bladder Tumor Samples
2.2. Cell Viability and Morphology
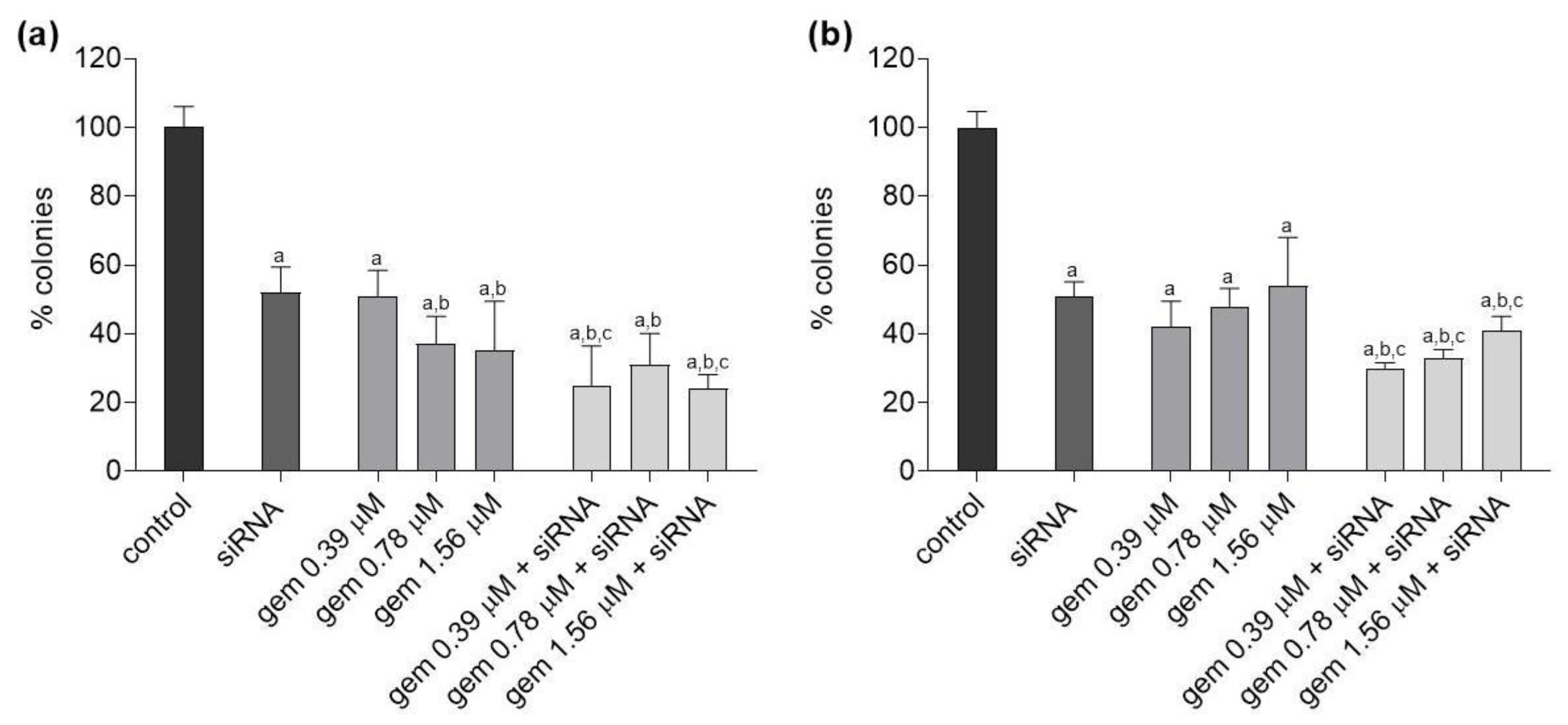
2.3. Clonogenic Survival
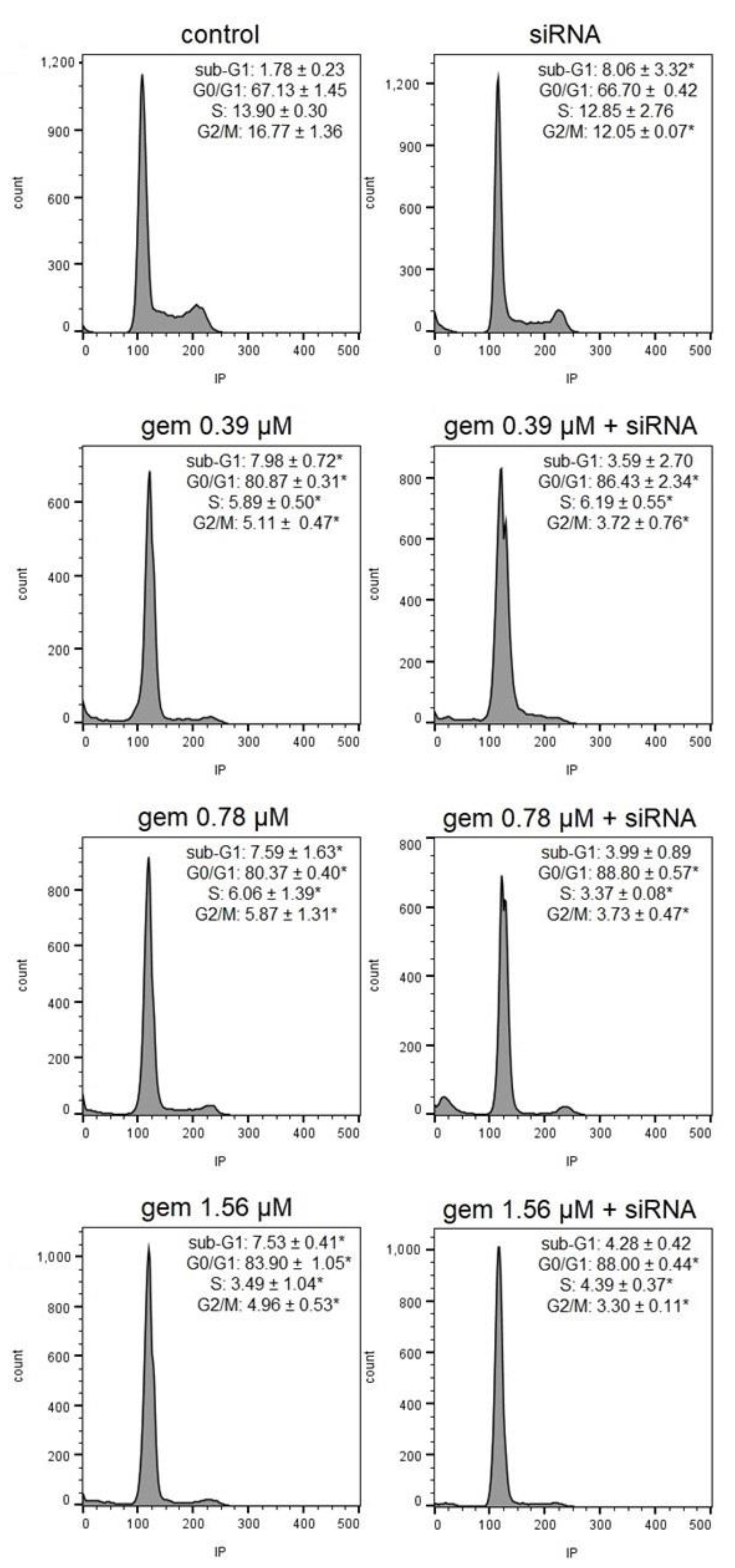
2.4. Cell Cycle Progression
2.5. Cell Migration
3. Discussion
4. Materials and Methods
4.1. Clinical Samples
4.2. Cell Lines, Culture Conditions, and Reagents
4.3. Expression and Knockdown Assays
4.4. Cytotoxicity Assay (XTT)
4.5. Clonogenic Survival Assay
4.6. Cell Morphology
4.7. Wound Healing Assay
4.8. Cell Cycle Progression
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Mushtaq, J.; Thurairaja, R.; Nair, R. Bladder cancer. Surgery 2019, 37, 529–537. [Google Scholar] [CrossRef]
- Sanli, O.; Dobruch, J.; Knowles, M.A.; Burger, M.; Alemozaffar, M.; Nielsen, M.E.; Lotan, Y. Bladder cancer. Nat. Rev. Dis. Prim. 2017, 3, 17022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saginala, K.; Barsouk, A.; Aluru, J.S.; Rawla, P.; Padala, S.A.; Barsouk, A. Epidemiology of Bladder Cancer. Med. Sci. 2020, 8, 15. [Google Scholar] [CrossRef] [Green Version]
- Chang, S.S.; Boorjian, S.A.; Chou, R.; Clark, P.E.; Daneshmand, S.; Konety, B.R.; Pruthi, R.; Quale, D.Z.; Ritch, C.R.; Seigne, J.D.; et al. Diagnosis and Treatment of Non-Muscle Invasive Bladder Cancer: AUA/SUO Guideline. J. Urol. 2016, 196, 1021–1029. [Google Scholar] [CrossRef]
- Yu, C.; Hequn, C.; Jinbo, C.; Feng, Z.; Xiongbing, Z.; Jian, D. Gemcitabine/cisplatin versus methotrexate/vinblastine/doxorubicin/cisplatin for muscle-invasive bladder cancer: A systematic review and meta-analysis. J. Cancer Res. Ther. 2018, 14, 1260–1265. [Google Scholar] [CrossRef]
- Schlack, K.; Boegemann, M.; Steinestel, J.; Schrader, A.J.; Krabbe, L.M. The safety and efficacy of gemcitabine for the treatment of bladder cancer. Expert Rev. Anticancer Ther. 2016, 16, 255–271. [Google Scholar] [CrossRef]
- Brunton, L.L.; Hilal-Dandan, R.; Knollmann, B.C. As Bases Farmacológicas da Terapêutica de Goodman e Gilman, 13th ed.; AMGH: Goderich, ON, Cananda, 2018. [Google Scholar]
- Gutschner, T.; Diederichs, S. The hallmarks of cancer. RNA Biol. 2012, 9, 703–719. [Google Scholar] [CrossRef] [Green Version]
- Jarroux, J.; Morillon, A.; Pinskaya, M. History, Discovery, and Classification of lncRNAs. Adv. Exp. Med. Biol. 2017, 1008, 1–46. [Google Scholar] [CrossRef]
- Li, C.H.; Chen, Y. Targeting long non-coding RNAs in cancers: Progress and prospects. Int. J. Biochem. Cell Biol. 2013, 45, 1895–1910. [Google Scholar] [CrossRef]
- Forrest, M.E.; Khalil, A.M. Review: Regulation of the cancer epigenome by long non-coding RNAs. Cancer Lett. 2017, 407, 106–112. [Google Scholar] [CrossRef]
- Lima, A.P.B.; da Silva, G.N. Long Non-Coding RNA and Chemoresistance in Bladder Cancer—A Mini Review. Cancer Investig. 2023, 41, 164–172. [Google Scholar] [CrossRef]
- Flippot, R.; Beinse, G.; Boilève, A.; Vibert, J.; Malouf, G.G. Long non-coding RNAs in genitourinary malignancies: A whole new world. Nat. Rev. Urol. 2019, 16, 484–504. [Google Scholar] [CrossRef]
- Wang, F.; Li, X.; Xie, X.; Zhao, L.; Chen, W. UCA1, a non-protein-coding RNA up-regulated in bladder carcinoma and embryo, influencing cell growth and promoting invasion. FEBS Lett. 2008, 582, 1919–1927. [Google Scholar] [CrossRef] [Green Version]
- Luo, J.; Chen, J.; Li, H.; Yang, Y.; Yun, H.; Yang, S.; Mao, X. LncRNA UCA1 promotes the invasion and EMT of bladder cancer cells by regulating the miR-143/HMGB1 pathway. Oncol. Lett. 2017, 14, 5556–5562. [Google Scholar] [CrossRef] [Green Version]
- Fan, Y.; Shen, B.; Tan, M.; Mu, X.; Qin, Y.; Zhang, F.; Liu, Y. Long non-coding RNA UCA1 increases chemoresistance of bladder cancer cells by regulating Wnt signaling. FEBS J. 2014, 281, 1750–1758. [Google Scholar] [CrossRef]
- Ying, L.; Chen, Q.; Wang, Y.; Zhou, Z.; Huang, Y.; Qiu, F. Upregulated MALAT-1 contributes to bladder cancer cell migration by inducing epithelial-to-mesenchymal transition. Mol. Biosyst. 2012, 8, 2289–2294. [Google Scholar] [CrossRef]
- Liu, P.; Li, X.; Cui, Y.; Chen, J.; Li, C.; Li, Q.; Li, H.; Zhang, X.; Zu, X. LncRNA-MALAT1 mediates cisplatin resistance via miR-101-3p/VEGF-C pathway in bladder cancer. Acta Biochim. Biophys. Sin. 2019, 51, 1148–1157. [Google Scholar] [CrossRef]
- Li, C.; Cui, Y.; Liu, L.F.; Ren, W.B.; Li, Q.Q.; Zhou, X.; Li, Y.L.; Li, Y.; Bai, X.Y.; Zu, X.B. High Expression of Long Noncoding RNA MALAT1 Indicates a Poor Prognosis and Promotes Clinical Progression and Metastasis in Bladder Cancer. Clin. Genitourin. Cancer 2017, 15, 570–576. [Google Scholar] [CrossRef]
- Lee, K.-H.; Kim, B.-C.; Jeong, S.-H.; Jeong, C.W.; Ku, J.H.; Kim, H.H.; Kwak, C. Histone Demethylase KDM7A Regulates Androgen Receptor Activity, and Its Chemical Inhibitor TC-E 5002 Overcomes Cisplatin-Resistance in Bladder Cancer Cells. Int. J. Mol. Sci. 2020, 21, 5658. [Google Scholar] [CrossRef]
- Lee, K.H.; Hong, S.; Kang, M.; Jeong, C.W.; Ku, J.H.; Kim, H.H.; Kwak, C. Histone demethylase KDM7A controls androgen receptor activity and tumor growth in prostate cancer. Int. J. Cancer 2018, 143, 2849–2861. [Google Scholar] [CrossRef] [Green Version]
- Meng, Z.; Liu, Y.; Wang, J.; Fan, H.; Fang, H.; Li, S.; Yuan, L.; Liu, C.; Peng, Y.; Zhao, W.; et al. Histone demethylase KDM7A is required for stem cell maintenance and apoptosis inhibition in breast cancer. J. Cell Physiol. 2020, 235, 932–943. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Liu, Y.; Pu, Y.-S.; Ma, Y.; Wang, J.-H.; Liu, E.-Q. JHDM1D-AS1 aggravates the development of gastric cancer through miR-450a-2-3p-PRAF2 axis. Life Sci. 2021, 265, 118805. [Google Scholar] [CrossRef] [PubMed]
- Yao, G.; Chen, K.; Qin, Y.; Niu, Y.; Zhang, X.; Xu, S.; Zhang, C.; Feng, M.; Wang, K. Long Non-coding RNA JHDM1D-AS1 Interacts with DHX15 Protein to Enhance Non-Small-Cell Lung Cancer Growth and Metastasis. Mol. Ther. Nucleic Acids 2019, 18, 831–840. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Han, L.; Roebuck, P.; Diao, L.; Liu, L.; Yuan, Y.; Weinstein, J.N.; Liang, H. TANRIC: An Interactive Open Platform to Explore the Function of lncRNAs in Cancer. Cancer Res. 2015, 75, 3728–3737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, X.; Liu, Q.; Chen, X.; Chen, S.; Yang, J.; Liu, Q.; Cheng, Y. Screening of tumor grade-related mRNAs and lncRNAs for Esophagus Squamous Cell Carcinoma. J. Clin. Lab Anal. 2021, 35, e23797. [Google Scholar] [CrossRef]
- Yang, X.; Wang, G.; Wang, Y.; Zhou, J.; Yuan, H.; Li, X.; Liu, Y.; Wang, B. Histone demethylase KDM7A reciprocally regulates adipogenic and osteogenic differentiation via regulation of C/EBPalpha and canonical Wnt signalling. J. Cell Mol. Med. 2019, 23, 2149–2162. [Google Scholar] [CrossRef]
- Osawa, T.; Muramatsu, M.; Wang, F.; Tsuchida, R.; Kodama, T.; Minami, T.; Shibuya, M. Increased expression of histone demethylase JHDM1D under nutrient starvation suppresses tumor growth via down-regulating angiogenesis. Proc. Natl. Acad. Sci. USA 2011, 108, 20725–20729. [Google Scholar] [CrossRef] [Green Version]
- Tee, A.E.; Liu, B.; Song, R.; Li, J.; Pasquier, E.; Cheung, B.B.; Jiang, C.; Marshall, G.M.; Haber, M.; Norris, M.D.; et al. The long noncoding RNA MALAT1 promotes tumor-driven angiogenesis by up-regulating pro-angiogenic gene expression. Oncotarget 2016, 7, 8663–8675. [Google Scholar] [CrossRef]
- Kondo, A.; Nonaka, A.; Shimamura, T.; Yamamoto, S.; Yoshida, T.; Kodama, T.; Aburatani, H.; Osawa, T. Long Noncoding RNA JHDM1D-AS1 Promotes Tumor Growth by Regulating Angiogenesis in Response to Nutrient Starvation. Mol. Cell. Biol. 2017, 37, e00125-17. [Google Scholar] [CrossRef] [Green Version]
- Lobo, N.; Mount, C.; Omar, K.; Nair, R.; Thurairaja, R.; Khan, M.S. Landmarks in the treatment of muscle-invasive bladder cancer. Nat. Rev. Urol. 2017, 14, 565–574. [Google Scholar] [CrossRef]
- Malek, E.; Jagannathan, S.; Driscoll, J.J. Correlation of long non-coding RNA expression with metastasis, drug resistance and clinical outcome in cancer. Oncotarget 2014, 5, 8027. [Google Scholar] [CrossRef] [Green Version]
- Pan, J.; Li, X.; Wu, W.; Xue, M.; Hou, H.; Zhai, W.; Chen, W. Long non-coding RNA UCA1 promotes cisplatin/gemcitabine resistance through CREB modulating miR-196a-5p in bladder cancer cells. Cancer Lett. 2016, 382, 64–76. [Google Scholar] [CrossRef]
- An, Q.; Zhou, L.; Xu, N. Long noncoding RNA FOXD2-AS1 accelerates the gemcitabine-resistance of bladder cancer by sponging miR-143. Biomed. Pharmacother. 2018, 103, 415–420. [Google Scholar] [CrossRef]
- Munshi, A.; Hobbs, M.; Meyn, R.E. Clonogenic cell survival assay. Methods Mol. Med. 2005, 110, 21–28. [Google Scholar] [CrossRef]
- da Silva, G.N.; de Camargo, E.A.; Salvadori, D.M. Toxicogenomic activity of gemcitabine in two TP53-mutated bladder cancer cell lines: Special focus on cell cycle-related genes. Mol. Biol. Rep. 2012, 39, 10373–10382. [Google Scholar] [CrossRef]
- Riahi, R.; Yang, Y.; Zhang, D.D.; Wong, P.K. Advances in Wound-Healing Assays for Probing Collective Cell Migration. SLAS Technol. 2012, 17, 59–65. [Google Scholar] [CrossRef]
- Dhamija, S.; Diederichs, S. From junk to master regulators of invasion: lncRNA functions in migration, EMT and metastasis. Int. J. Cancer 2016, 139, 269–280. [Google Scholar] [CrossRef]
- Montano, R.; Thompson, R.; Chung, I.; Hou, H.; Khan, N.; Eastman, A. Sensitization of human cancer cells to gemcitabine by the Chk1 inhibitor MK-8776: Cell cycle perturbation and impact of administration schedule in vitro and in vivo. BMC Cancer 2013, 13, 604. [Google Scholar] [CrossRef] [Green Version]
- Pandey, S.; Bourn, J.; Cekanova, M. Mutations of p53 decrease sensitivity to the anthracycline treatments in bladder cancer cells. Oncotarget 2018, 9, 28514. [Google Scholar] [CrossRef]
- Earl, J.; Rico, D.; Carrillo-de-Santa-Pau, E.; Rodriguez-Santiago, B.; Mendez-Pertuz, M.; Auer, H.; Gomez, G.; Grossman, H.B.; Pisano, D.G.; Schulz, W.A.; et al. The UBC-40 Urothelial Bladder Cancer cell line index: A genomic resource for functional studies. BMC Genom. 2015, 16, 403. [Google Scholar] [CrossRef] [Green Version]
- Rieger, K.M.; Little, A.F.; Swart, J.M.; Kastrinakis, W.V.; Fitzgerald, J.M.; Hess, D.T.; Libertino, J.A.; Summerhayes, I.C. Human bladder carcinoma cell lines as indicators of oncogenic change relevant to urothelial neoplastic progression. Br. J. Cancer 1995, 72, 683–690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersen, C.L.; Jensen, J.L.; Orntoft, T.F. Normalization of real-time quantitative reverse transcription-PCR data: A model-based variance estimation approach to identify genes suited for normalization, applied to bladder and colon cancer data sets. Cancer Res. 2004, 64, 5245–5250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lima, A.P.B.; Almeida, T.C.; Barros, T.M.B.; Rocha, L.C.M.; Garcia, C.C.M.; da Silva, G.N. Toxicogenetic and antiproliferative effects of chrysin in urinary bladder cancer cells. Mutagenesis 2020, 35, 361–371. [Google Scholar] [CrossRef]
- da Silva, G.N.; de Castro Marcondes, J.P.; de Camargo, E.A.; da Silva Passos Junior, G.A.; Sakamoto-Hojo, E.T.; Salvadori, D.M. Cell cycle arrest and apoptosis in TP53 subtypes of bladder carcinoma cell lines treated with cisplatin and gemcitabine. Exp. Biol. Med. 2010, 235, 814–824. [Google Scholar] [CrossRef]
- Almeida, T.C.; Guerra, C.C.C.; De Assis, B.L.G.; de Oliveira Aguiar Soares, R.D.; Garcia, C.C.M.; Lima, A.A.; da Silva, G.N. Antiproliferative and toxicogenomic effects of resveratrol in bladder cancer cells with different TP53 status. Environ. Mol. Mutagen. 2019, 60, 740–751. [Google Scholar] [CrossRef]
- Akoglu, H. User’s guide to correlation coefficients. Turk. J. Emerg. Med. 2018, 18, 91–93. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pereira, I.O.A.; da Silva, G.N.; Almeida, T.C.; Lima, A.P.B.; Sávio, A.L.V.; Leite, K.R.M.; Salvadori, D.M.F. LncRNA JHDM1D-AS1 Is a Key Biomarker for Progression and Modulation of Gemcitabine Sensitivity in Bladder Cancer Cells. Molecules 2023, 28, 2412. https://doi.org/10.3390/molecules28052412
Pereira IOA, da Silva GN, Almeida TC, Lima APB, Sávio ALV, Leite KRM, Salvadori DMF. LncRNA JHDM1D-AS1 Is a Key Biomarker for Progression and Modulation of Gemcitabine Sensitivity in Bladder Cancer Cells. Molecules. 2023; 28(5):2412. https://doi.org/10.3390/molecules28052412
Chicago/Turabian StylePereira, Isadora Oliveira Ansaloni, Glenda Nicioli da Silva, Tamires Cunha Almeida, Ana Paula Braga Lima, André Luiz Ventura Sávio, Katia Ramos Moreira Leite, and Daisy Maria Fávero Salvadori. 2023. "LncRNA JHDM1D-AS1 Is a Key Biomarker for Progression and Modulation of Gemcitabine Sensitivity in Bladder Cancer Cells" Molecules 28, no. 5: 2412. https://doi.org/10.3390/molecules28052412
APA StylePereira, I. O. A., da Silva, G. N., Almeida, T. C., Lima, A. P. B., Sávio, A. L. V., Leite, K. R. M., & Salvadori, D. M. F. (2023). LncRNA JHDM1D-AS1 Is a Key Biomarker for Progression and Modulation of Gemcitabine Sensitivity in Bladder Cancer Cells. Molecules, 28(5), 2412. https://doi.org/10.3390/molecules28052412