
Evaluation of Biological Activities of Twenty Flavones and In Silico Docking Study

 ,
,  , and
, and 
Abstract
:1. Introduction
2. Results and Discussion
2.1. Antioxidant Activity
2.2. Biological Activity
2.2.1. Anti-α-Amylase Activity
2.2.2. Effects of Flavonoids on the Inhibition of SOD and XOD
2.2.3. Anti-AChE Activity
2.2.4. Anti-Inflammatory Activity
2.2.5. Cytotoxic Activity against Cancer Cell Lines
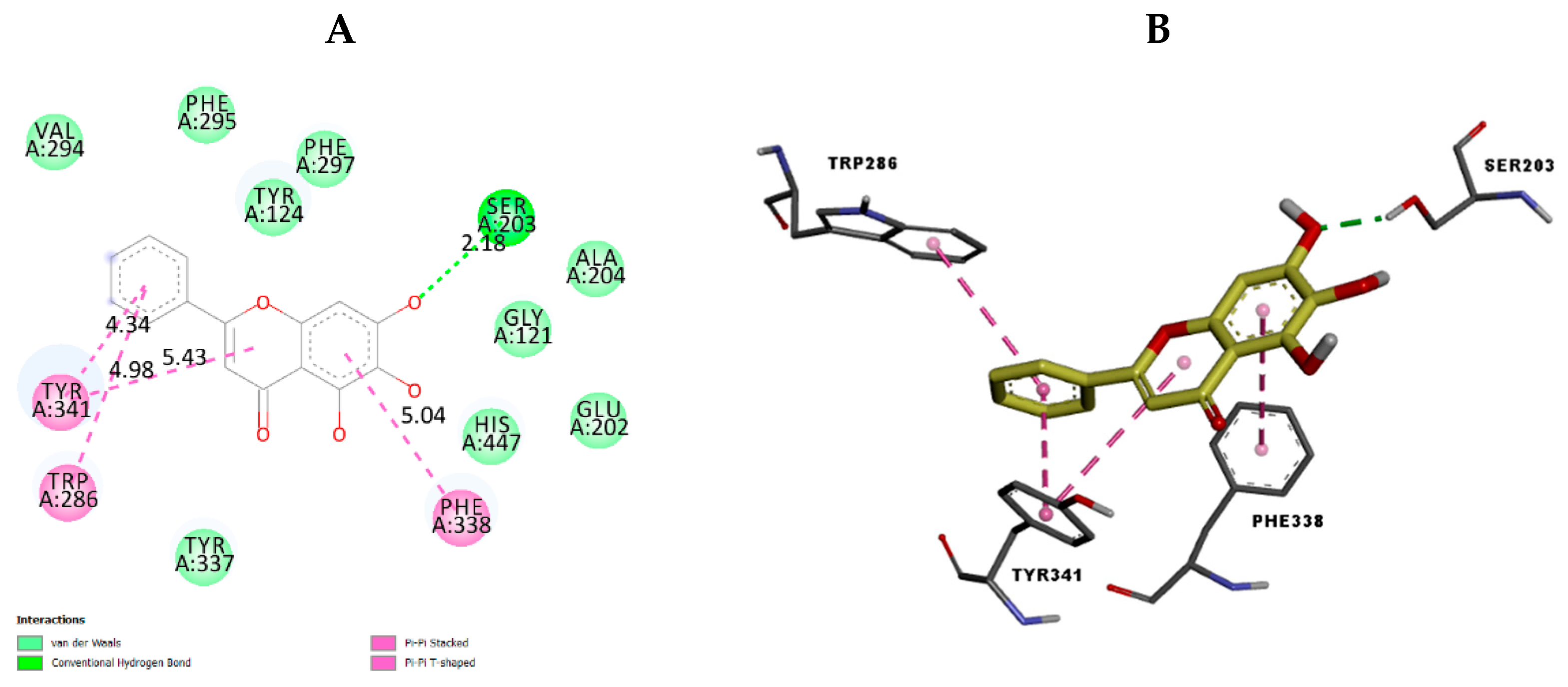
2.3. In Silico Docking Study
3. Materials and Methods
3.1. Preparation of Molecules
3.2. Antioxidant Activity and Biological Activities
3.2.1. Spectrophotometry
Free Radical Scavenging Activity
Anti-α-Amylase Activity
Anti-Xanthine Oxidase Activity
Anti-Superoxide Dismutase Activity
Anti-Cholinesterase Activity
Anti-Inflammatory Activity
Cytotoxic Activity
3.2.2. Molecular Docking
3.2.3. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Pietta, P.G. Flavonoids as antioxidants. J. Nat. Prod. 2000, 63, 1035–1042. [Google Scholar] [CrossRef]
- Sak, K. Cytotoxicity of dietary flavonoids on different human cancer types. Pharm. Rev. 2014, 8, 122–146. [Google Scholar] [CrossRef] [Green Version]
- Agarwal, A.A.; Jadhav, P.R.; Deshmukh, Y.A. Prescribing pattern and efficacy of anti-diabetic drugs in maintaining optimal glycemic levels in diabetic patients. J. Basic Clin. Pharm. 2014, 5, 79–83. [Google Scholar] [CrossRef] [Green Version]
- Standl, E.; Schnell, O. Alpha-glucosidase inhibitors 2012 - cardiovascular considerations and trial evaluation. Diab. Vasc. Dis. Res. 2012, 9, 163–169. [Google Scholar] [CrossRef]
- Gao, J.; Xu, P.; Wang, Y.; Wang, Y.; Hochstetter, D. Combined Effects of Green Tea Extracts, Green Tea Polyphenols or Epigallocatechin Gallate with Acarbose on Inhibition against α-Amylase and α-Glucosidase In Vitro. Molecules 2013, 18, 11614–11623. [Google Scholar] [CrossRef] [Green Version]
- Kee, K.T.; Koh, M.; Oong, L.X.; Ng, K. Screening culinary herbs for antioxidant and α-glucosidase inhibitory activities. Int. J. Food Sci. Technol. 2013, 48, 1884–1891. [Google Scholar] [CrossRef]
- Yang, S.-F.; Yang, W.-E.; Kuo, W.-H.; Chang, H.-R.; Chu, S.-C.; Hsieh, Y.-S. Antimetastatic potentials of flavones on oral cancer cell via an inhibition of matrix-degrading proteases. Arch. Oral Biol. 2008, 53, 287–294. [Google Scholar] [CrossRef]
- Costantino, L.; Albasini, A.; Rastelli, G.; Benvenuti, S. Activity of polyphenolic crude extracts as scavengers of superoxide radicals and inhibitors of xanthine oxidase. Planta Med. 1992, 58, 342–344. [Google Scholar] [CrossRef]
- Bohnen, N.I.; Albin, R.L. The cholinergic system and Parkinson disease. Behav. Brain Res. 2011, 221, 564–573. [Google Scholar] [CrossRef] [Green Version]
- Jaiswal, J.V.; Wadegaonkar, P.A.; Hajare, S.W. The Bioflavonoid Galangin Suppresses the Growth of Ehrlich Ascites Carcinoma in Swiss Albino Mice: A Molecular Insight. Appl. Biochem. Biotechnol. 2012, 167, 1325–1339. [Google Scholar] [CrossRef]
- Shen, S.-C.; Ko, C.H.; Tseng, S.-W.; Tsai, S.-H.; Chen, Y.-C. Structurally related antitumor effects of flavanones in vitro and in vivo: Involvement of caspase 3 activation, p21 gene expression, and reactive oxygen species production. Toxicol. Appl. Pharmacol. 2004, 197, 84–95. [Google Scholar] [CrossRef]
- Attoub, S.; Hassan, A.H.; Vanhoecke, B.; Iratni, R.; Takahashi, T.; Gaben, A.-M.; Bracke, M.; Awad, S.; John, A.; Kamalboor, H.A.; et al. Inhibition of cell survival, invasion, tumor growth and histone deacetylase activity by the dietary flavonoid luteolin in human epithelioid cancer cells. Eur. J. Pharmacol. 2011, 651, 18–25. [Google Scholar] [CrossRef]
- Jeong, J.C.; Kim, M.S.; Kim, T.H.; Kim, Y.K. Kaempferol induces cell death through ERK and Akt-dependent down-regulation of XIAP and survivin in human glioma cells. Neurochem. Res. 2009, 34, 991–1001. [Google Scholar] [CrossRef]
- Li, Y.-L.; Gan, G.-P.; Zhang, H.-Z.; Wu, H.-Z.; Li, C.-L.; Huang, Y.-P.; Liu, Y.-W.; Liu, J.-W. A flavonoid glycoside isolated from Smilax china L. rhizome in vitro anticancer effects on human cancer cell lines. J. Ethnopharmacol. 2007, 113, 115–124. [Google Scholar] [CrossRef]
- Tokalov, S.V.; Abramyuk, A.M.; Abolmaali, N.D. Protection of p53 wild type cells from taxol by genistein in the combined treatment of lung cancer. Nutr. Cancer 2010, 62, 795–801. [Google Scholar] [CrossRef]
- Choi, S.-M.; Kim, B.C.; Cho, Y.-H.; Choi, K.-H.; Chang, J.; Park, M.-S.; Kim, M.-K.; Cho, K.-H.; Kim, J.-K. Effects of Flavonoid Compounds on β-amyloid-peptide-induced Neuronal Death in Cultured Mouse Cortical Neurons. Chonnam. Med. J. 2014, 50, 45–51. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.J.; Oh, Y.C.; Cho, W.K.; Ma, J.Y. Antioxidant and Anti-Inflammatory Activity Determination of One Hundred Kinds of Pure Chemical Compounds Using Offline and Online Screening HPLC Assay. Evid.-Based Complement. Altern. Med. 2015, 2015, 165457. [Google Scholar] [CrossRef] [Green Version]
- Ng, K. Evaluation of α-Amylase and α-Glucosidase Inhibitory Activity of Flavonoids. Int. J. Food Nutr. Sci. 2015, 2, 1–6. [Google Scholar] [CrossRef]
- Piparo, E.L.; Scheib, H.; Frei, N.; Williamson, G.; Grigorov, M.; Chou, C.J. Flavonoids for Controlling Starch Digestion: Structural Requirements for Inhibiting Human α-Amylase. J. Med. Chem. 2008, 51, 3555–3561. [Google Scholar] [CrossRef]
- Wei, D.; Tang, J.; Bai, W.; Wang, Y.; Zhang, Z. Ameliorative effects of baicalein on an amyloid-β induced Alzheimer’s disease rat model: A proteomics study. Curr. Alzheimer. Res. 2014, 11, 869–881. [Google Scholar] [CrossRef]
- Li-Weber, M.; Rhoads, C.P.; Abelson, P.H.; Newman, D.J. New therapeutic aspects of flavones: The anticancer properties of Scutellaria and its main active constituents Wogonin, Baicalein and Baicalin. Cancer Treat. Rev. 2009, 35, 57–68. [Google Scholar] [CrossRef]
- Lebeau, A.; Esclaire, F.; Rostène, W.; Pélaprat, D. Baicalein protects cortical neurons from beta-amyloid (25-35) induced toxicity. Neuroreport 2001, 12, 2199–2202. [Google Scholar] [CrossRef]
- Chan, F.L.; Choi, H.L. Induction of apoptosis in prostate cancer cell lines by a flavonoid, baicalin. Cancer Lett. 2000, 160, 219–228. [Google Scholar] [CrossRef]
- Ghotbi, G.; Mahdavi, M.; Najafi, Z.; Moghadam, F.H.; Hamzeh-Mivehroud, M.; Davaran, S.; Dastmalchi, S. Design, synthesis, biological evaluation, and docking study of novel dual-acting thiazole-pyridiniums inhibiting acetylcholinesterase and β-amyloid aggregation for Alzheimer’s disease. Bioorg. Chem. 2020, 103, 104186. [Google Scholar] [CrossRef]
- Savithri, K.; Kumar, B.C.V.; Vivek, H.K.; Revanasiddappa, H.D. Synthesis and Characterization of Cobalt(III) and Copper(II) Complexes of 2-((E)-(6-Fluorobenzo[d]thiazol-2-ylimino) methyl)-4-chlorophenol: DNA Binding and Nuclease Studies—SOD and Antimicrobial Activities. Int. J. Spectrosc. 2018, 2018, 8759372. [Google Scholar] [CrossRef]
- Joshi, G.; Sharma, M.; Kalra, S.; Gavande, N.S.; Singh, S.; Kumar, R. Design, synthesis, biological evaluation of 3,5-diaryl-4,5-dihydro-1H-pyrazole carbaldehydes as non-purine xanthine oxidase inhibitors: Tracing the anticancer mechanism via xanthine oxidase inhibition. Bioorg. Chem. 2021, 107, 104620. [Google Scholar] [CrossRef]
- Chaudhry, F.; Shahid, W.; Al-Rashida, M.; Ashraf, M.; Munawar, M.A.; Khan, M.A. Synthesis of imidazole-pyrazole conjugates bearing aryl spacer and exploring their enzyme inhibition potentials. Bioorg. Chem. 2021, 108, 104686. [Google Scholar] [CrossRef]
- Ramasubbu, N.; Paloth, V.; Luo, Y.; Brayer, G.D.; Levine, M.J. Structure of Human Salivary α-Amylase at 1.6 Å Resolution: Implications for its Role in the Oral Cavity. Acta Crystallogr. Sect. D Biol. Crystallogr. 1996, 52, 435–446. [Google Scholar] [CrossRef]
- Williams, L.K.; Li, C.; Withers, S.G.; Brayer, G.D. Order and disorder: Differential structural impacts of myricetin and ethyl caffeate on human amylase, an antidiabetic target. J. Med. Chem. 2012, 55, 10177–10186. [Google Scholar] [CrossRef]
- El Euch, S.K.; Bouajila, J.; Bouzouita, N. Chemical composition, biological and cytotoxic activities of Cistus salviifolius flower buds and leaves extracts. Industrial Crops and Products. Ind. Crops Prod. 2015, 76, 1100–1105. [Google Scholar] [CrossRef]
- Cheung, J.; Rudolph, M.; Burshteyn, F.; Cassidy, M.; Gary, E.; Love, J.; Franklin, M.; Height, J. Structures of human acetylcholinesterase in complex with pharmacologically important ligands. J. Med. Chem. 2012, 55, 10282–10286. [Google Scholar] [CrossRef]
- Hough, M.A.; Hasnain, S.S. Crystallographic structures of bovine copper-zinc superoxide dismutase reveal asymmetry in two subunits: Functionally important three and five coordinate copper sites captured in the same crystal. J. Mol. Biol. 1999, 287, 579–592. [Google Scholar] [CrossRef]
- Enroth, C.; Eger, B.T.; Okamoto, K.; Nishino, T.; Nishino, T.; Pai, E.F. Crystal structures of bovine milk xanthine dehydrogenase and xanthine oxidase: Structure-based mechanism of conversion. Proc. Natl. Acad. Sci. USA 2000, 97, 10723–10728. [Google Scholar] [CrossRef] [Green Version]
- Gilbert, N.C.; Rui, Z.; Neau, D.B.; Waight, M.T.; Bartlett, S.G.; Boeglin, W.E.; Brash, A.R.; Newcomer, M.E. Conversion of human 5-lipoxygenase to a 15-lipoxygenase by a point mutation to mimic phosphorylation at Serine-663. FASEB J. 2012, 26, 3222–3229. [Google Scholar] [CrossRef] [Green Version]
- Caner, S.; Brayer, G.D. Human Pancreatic Alpha-Amylase in Complex with Mini-Montbretin A. Can. Inst. Health Res. (CIHR) 2016. [Google Scholar] [CrossRef]
- Mseddi, K.; Alimi, F.; Noumi, E.; Veettil, V.N.; Deshpande, S.; Adnan, M.; Hamdi, A.; Elkahoui, S.; Alghamdi, A.; Kadri, A.; et al. Thymus musilii Velen. as a promising source of potent bioactive compounds with its pharmacological properties: In vitro and in silico analysis. Arabian Journal of Chemistry. Arab. J. Chem. 2020, 13, 6782–6801. [Google Scholar] [CrossRef]
- Morris, G.M.; Huey, R.; Olson, A.J. Using AutoDock for Ligand-Receptor Docking. Curr. Protoc. Bioinform. 2008, 24, 8.14.1–8.14.40. [Google Scholar] [CrossRef]
- Pedretti, A.; Villa, L.; Vistoli, G.J. VEGA—An open platform to develop chemo-bio-informatics applications, using plug-in architecture and script programming. Comput.-Aided Mol. Des. 2004, 18, 167–173. [Google Scholar] [CrossRef]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization and multithreading. J. Comput. Chem. 2010, 31, 455. [Google Scholar] [CrossRef] [Green Version]
- Electronic Lab Notebooks—Cloud-Based Data Capture and Knowledge Sharing—BIOVIA—Dassault Systèmes®. Available online: https://www.3ds.com/products-services/biovia/products/laboratory-informatics/electronic-lab-notebooks/ (accessed on 1 July 2022).








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
 Flavonoids’ Basic Structure | |||
|---|---|---|---|
| N° | Name and Structure | N° | Name and Structure |
| M1 |  Chrysin (or 5,7-dihydroxyflavone) | M11 |  5-hydroxy-3′-methoxyflavone |
| M2 |  Icariin | M12 |  4′,5-dihydroxy-7-methoxyflavone |
| M3 |  6-hydroxyflavone | M13 |  7,3′-dihydroxyflavone |
| M4 |  5-hydroxyflavone | M14 |  5,3′-dihydroxyflavone |
| M5 |  7-hydroxyflavone | M15 |  5-hydroxy-7-((3-methylbenzyl)oxy)-2-phenyl-4h-chromen-4-one |
| M6 |  trihydroxyethylrutin | M16 |  3′-hydroxy-6-methylflavone |
| M7 |  Baicalein (or 5, 6, 7-trihydroxyflavone) | M17 |  6-hydroxy-4′-methylflavone |
| M8 |  3′,5′-dihydroxyflavone | M18 |  7-hydroxy-3′,4′,5′-trimethoxy-α-naphthoflavone |
| M9 |  3′-hydroxy-b-naphthoflavone | M19 |  Diosmin (or 3′, 5,7-trihydroxy-4′-methoxyflavone 7-rutinoside) |
| M10 |  3′-hydroxy-a-naphthoflavone | M20 |  myricitrin dihydrate |
| Antioxidant | ||||
|---|---|---|---|---|
| Molecule | DPPH Assay | ABTS Assay | ||
| % | IC50 | % | IC50 | |
| M1 | na | >100 | 60.1 ± 3.1 | >50 |
| M2 | na | >100 | 87.8 ± 0.9 | 20.1 ± 0.1 |
| M3 | na | >100 | 95.0 ± 1.2 | 24.0 ± 0.1 |
| M4 | na | >100 | 16.0 ± 1.5 | >100 |
| M5 | na | >100 | 5.9 ± 0.4 | >100 |
| M6 | na | >100 | 96.0 ± 0.8 | 24.5 ± 0.1 |
| M7 | 96.4 ± 0.1 | 5.2 ± 0.2 | 99.4 ± 0.3 | 6.3 ± 0.1 |
| M8 | 20.3 ± 4.1 | >100 | 100.1 ± 0.4 | 7.3 ± 0.1 |
| M9 | na | >100 | na | >100 |
| M10 | na | >100 | 11.0 ± 1.0 | >100 |
| M11 | na | >100 | 17.7 ± 0.7 | >100 |
| M12 | na | >100 | 30.3 ± 0.6 | >100 |
| M13 | na | >100 | 89.9 ± 0.2 | 24.5 ± 0.1 |
| M14 | na | >100 | 16.9 ± 0.3 | >100 |
| M15 | na | >100 | 25.6 ± 0.7 | >100 |
| M16 | na | >100 | 56.7 ± 0.7 | >50 |
| M17 | na | >100 | 65.5 ± 0.6 | >50 |
| M18 | 40.8 ± 0.6 | >100 | 50.7 ± 0.7 | >50 |
| M19 | na | >100 | 66.6 ± 0.9 | >50 |
| M20 | na | >100 | 20.3 ± 3.2 | >100 |
| Vitamin C | 2.3 ± 0.2 | 2.2 ± 0.2 | ||
| Anti ACHE | Anti SOD | Anti XOD | Anti 15-Lipoxygenase | Anti α-Amylase | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Molecule | % | IC50 | % | IC50 | % | IC50 | % | IC50 | % | IC50 |
| M1 | 45.8 ± 1.3 | >100 | 41.5 ± 1.8 | >100 | na | >100 | na | >100 | 59.7 ± 2.0 | 36.6 ± 0.2 |
| M2 | 19.3 ± 0.3 | >100 | 61.7 ± 1.1 | >50 | na | >100 | na | >100 | 58.1 ± 2.2 | 14.1 ± 2.1 |
| M3 | 40.1 ± 2.8 | >100 | 54.8 ± 3.2 | >50 | na | >100 | na | >100 | 90.3 ± 3.2 | 10.9 ± 0.1 |
| M4 | 42.3 ± 2.1 | >100 | 25.9 ± 2.5 | >100 | na | >100 | na | >100 | 58.4 ± 1.4 | 37.9 ± 0.8 |
| M5 | 47.7 ± 0.7 | >100 | 74.3 ± 3.02 | 36.5 ± 1.2 | na | >100 | na | >100 | 90.3 ± 3.2 | 1.2 ± 0.1 |
| M6 | 33.7 ± 3.7 | >100 | 53.7 ± 2.1 | >50 | na | >100 | na | >100 | 49.4 ± 5.1 | >100 |
| M7 | 62.0 ± 1.9 | 10.2 ± 3.0 | 71.2 ± 2.3 | 31.5 ± 0.5 | na | >100 | 68.8 ± 2.9 | 38.5 ± 2.1 | 96.9 ± 1.7 | 3.4 ± 0.1 |
| M8 | 49.3 ± 2.2 | >100 | 52.3 ± 0.9 | >50 | 75.3 ± 1.6 | 2.3 ± 0.1 | na | >100 | 104.8 ± 3.5 | 14.9 ± 0.3 |
| M9 | 47.9 ± 4.0 | >100 | 19.8 ± 2.9 | >100 | na | >100 | na | >100 | 86.6 ± 2.3 | 2.0 ± 0.2 |
| M10 | 54.7 ± 1.9 | >50 | 39.4 ± 1.9 | >100 | 58.8 ± 0.3 | >50 | na | >100 | 69.4 ± 1.6 | 2.3 ± 0.1 |
| M11 | 52.6 ± 4.2 | >50 | 9.4 ± 3.1 | >100 | na | >100 | na | >100 | 124.8 ± 15.3 | 8.3 ± 0.2 |
| M12 | 51.0 ± 0.3 | >50 | 42.1 ± 5.3 | >100 | 58.2 ± 3.1 | 1.2 ± 0.1 | na | >100 | 111.7 ± 0.8 | 9.0 ± 0.1 |
| M13 | 63.9 ± 3.3 | >50 | 56.6 ± 1.7 | >50 | 64.2 ± 1.2 | 0.9 ± 0.2 | na | >100 | 91.7 ± 4.9 | 1.4 ± 0.1 |
| M14 | 55.0 ± 1.9 | >50 | 5.7 ± 2.5 | >100 | 71.3 ± 4.7 | 2.0 ± 0.1 | na | >100 | 109.6 ± 1.7 | 6.7 ± 0.9 |
| M15 | 55.3 ± 2.7 | >50 | 26.5 ± 3.3 | >100 | na | >100 | na | >100 | 46.2 ± 4.4 | >100 |
| M16 | 59.1 ± 1.5 | >50 | 27.3 ± 3.2 | >100 | 83.5 ± 3.8 | 2.0 ± 0.1 | na | >100 | na | >100 |
| M17 | 50.8 ± 4.1 | >50 | 80.9 ± 0.2 | >50 | 67.7 ± 2.1 | 0.8 ± 0.1 | na | >100 | na | >100 |
| M18 | 43.3 ± 1.8 | >100 | 32.7 ± 3.9 | >100 | 19.4 ± 1.0 | >100 | na | >100 | na | >100 |
| M19 | 37.7 ± 4.6 | >100 | 57.8 ± 2.4 | >50 | 45.3 ± 3.2 | >100 | na | >100 | 60.6 ± 4.8 | 3.7 ± 0.3 |
| M20 | 51.8 ± 3.5 | >50 | 42.4 ± 1.1 | >100 | 30.9 ± 2.5 | >100 | na | >100 | na | >100 |
| Galantamin | 1.4 ± 0.3 | |||||||||
| NDGA | 2.5 ± 0.4 | |||||||||
| Acarbose | 1.5 ± 0.1 | |||||||||
| Allopurinol | 1.3 ± 0.1 | |||||||||
| Cancer Cell Line | ||||
|---|---|---|---|---|
| Molecule | HCT116 | MCF7 | OVCAR-3 | SKOV-3 |
| M1 | >50 | 35.9 ± 0.8 | >50 | >50 |
| M2 | >50 | >100 | >50 | >50 |
| M3 | >50 | >50 | 44.7 ± 1.2 | >50 |
| M4 | >50 | >100 | >50 | >50 |
| M5 | >50 | >50 | >50 | >50 |
| M6 | >50 | >50 | >50 | >50 |
| M7 | 38.5 ± 1.3 | 33.1 ± 1.4 | >50 | 15.6 ± 0.1 |
| M8 | 33.1 ± 0.1 | 40.4 ± 0.7 | >50 | >50 |
| M9 | >50 | >50 | >50 | >50 |
| M10 | >50 | >50 | >50 | >50 |
| M11 | >50 | >100 | >50 | >50 |
| M12 | 38.5 ± 1.2 | >50 | 47.0 ± 1.4 | 44.7 ± 1.2 |
| M13 | 39.3 ± 0.8 | 38.5 ± 1.2 | >50 | 42.1 ± 0.7 |
| M14 | 4.6 ± 0.2 | >50 | >50 | >50 |
| M15 | >50 | >50 | 45.6 ± 1.4 | >50 |
| M16 | >100 | >100 | >50 | 42.1 ± 1.5 |
| M17 | 40.4 ± 1.2 | >50 | >50 | 43.8 ± 0.9 |
| M18 | 45.0 ± 0.9 | >50 | >50 | 37.3 ± 1.0 |
| M19 | >50 | >50 | >50 | 40.1 ± 0.9 |
| M20 | >50 | >50 | >50 | >50 |
| Tamoxifen | 1.0 ± 0.2 | 1.0 ± 0.1 | 1.4 ± 0.3 | 1.3 ± 0.1 |
| Compound | 4EY7 | 1CB4 | 1FIQ | 3V99 | 5e0F |
|---|---|---|---|---|---|
| M1 | −10.4 | −7.8 | −10.0 | −7.6 | −8.5 |
| M2 | −8.2 | −8.6 | −7.3 | −8.7 | −8.6 |
| M3 | −10.2 | −7.0 | −9.6 | −7.3 | −8.5 |
| M4 | −10.3 | −7.2 | −9.8 | −7.7 | −8.7 |
| M5 | −10.4 | −7.9 | −9.6 | −7.6 | −8.5 |
| M6 | −9.0 | −8.2 | −6.5 | −8.6 | −7.8 |
| M7 | −10.6 | −7.4 | −10.2 | −7.5 | −8.3 |
| M8 | −10.3 | −8.1 | −10.0 | −7.5 | −9.1 |
| M9 | −12.4 | −8.1 | −10.2 | −8.8 | −10.2 |
| M10 | −11.2 | −8.1 | −10.3 | −8.7 | −10.3 |
| M11 | −10.5 | −7.5 | −9.8 | −7.7 | −8.7 |
| M12 | −10.4 | −7.9 | −9.5 | −7.4 | −9.0 |
| M13 | −10.5 | −8.0 | −10.1 | −7.7 | −8.8 |
| M14 | −10.3 | −7.7 | −9.9 | −7.6 | −8.9 |
| M15 | −11.6 | −8.2 | −10.9 | −9.6 | −9.7 |
| M16 | −10.9 | −7.8 | −9.7 | −7.9 | −9.3 |
| M17 | −10.2 | −7.2 | −9.3 | −7.3 | −8.3 |
| M18 | −10.4 | −7.4 | −7.6 | −8.0 | −8.9 |
| M19 | −10.9 | −8.9 | −8.8 | −8.8 | −9.3 |
| M20 | −8.1 | −8.8 | −6.2 | −8.3 | −8.3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Belaiba, M.; Aldulaijan, S.; Messaoudi, S.; Abedrabba, M.; Dhouib, A.; Bouajila, J. Evaluation of Biological Activities of Twenty Flavones and In Silico Docking Study. Molecules 2023, 28, 2419. https://doi.org/10.3390/molecules28062419
Belaiba M, Aldulaijan S, Messaoudi S, Abedrabba M, Dhouib A, Bouajila J. Evaluation of Biological Activities of Twenty Flavones and In Silico Docking Study. Molecules. 2023; 28(6):2419. https://doi.org/10.3390/molecules28062419
Chicago/Turabian StyleBelaiba, Meriam, Sarah Aldulaijan, Sabri Messaoudi, Manef Abedrabba, Adnene Dhouib, and Jalloul Bouajila. 2023. "Evaluation of Biological Activities of Twenty Flavones and In Silico Docking Study" Molecules 28, no. 6: 2419. https://doi.org/10.3390/molecules28062419
APA StyleBelaiba, M., Aldulaijan, S., Messaoudi, S., Abedrabba, M., Dhouib, A., & Bouajila, J. (2023). Evaluation of Biological Activities of Twenty Flavones and In Silico Docking Study. Molecules, 28(6), 2419. https://doi.org/10.3390/molecules28062419