
Diagonal Interactions between Glutamate and Arginine Analogs with Varying Side-Chain Lengths in a β-Hairpin


Abstract
:1. Introduction
2. Results
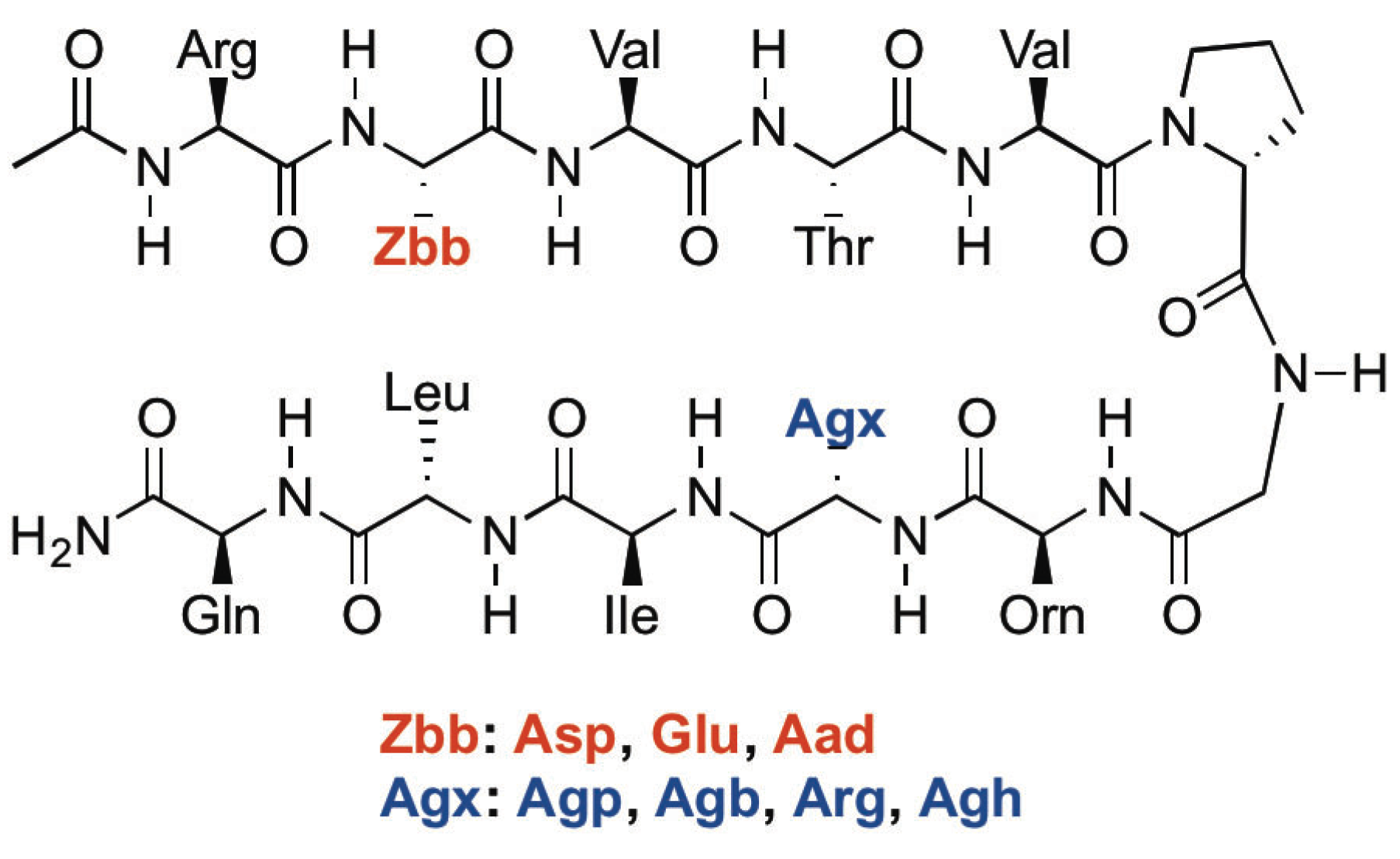
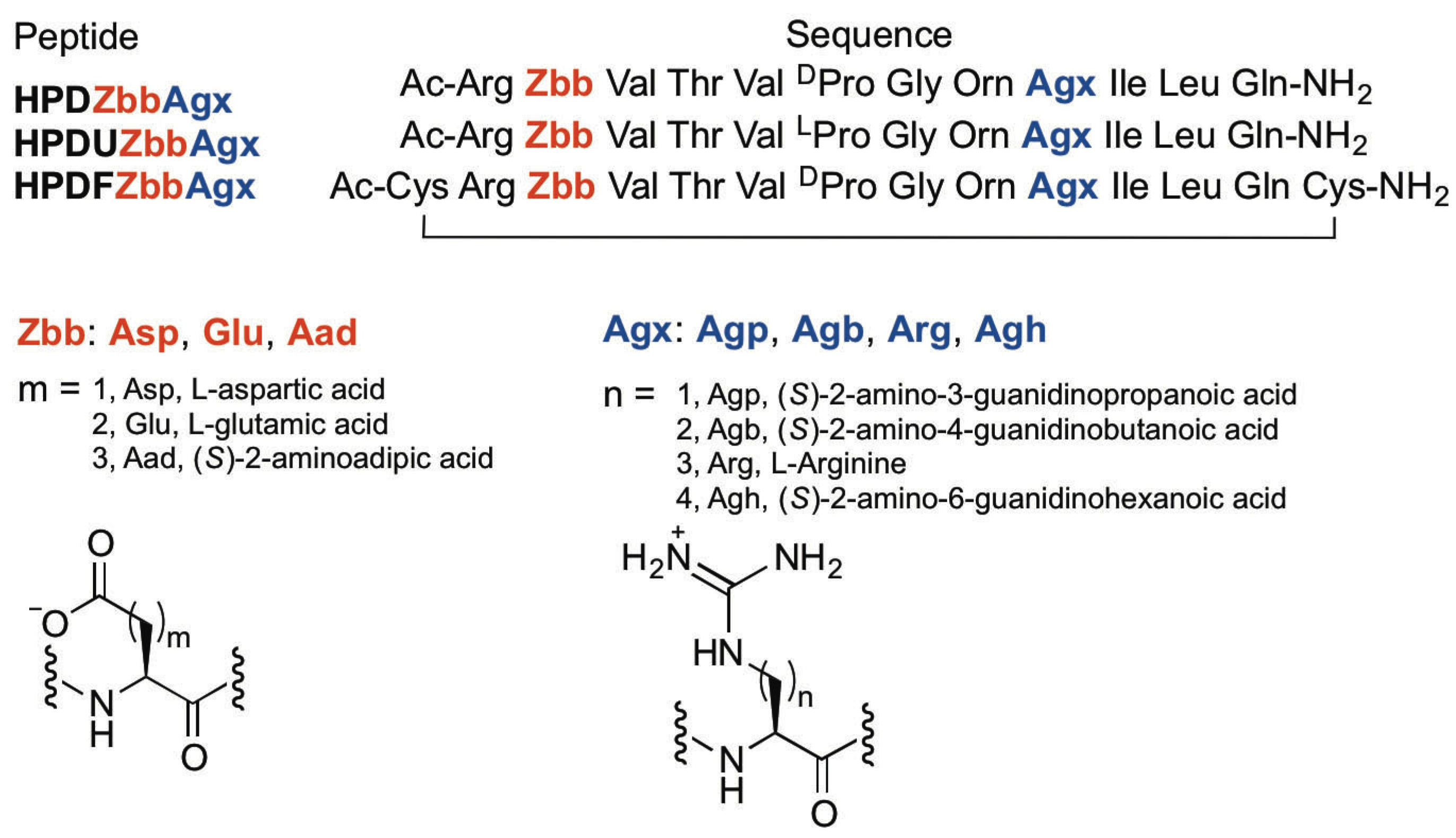
2.1. Peptide Design
2.2. Peptide Synthesis and Purification
2.3. NMR Characterization
2.4. Fraction Folded Population and Folding Free Energy
2.5. Diagonal Zbb2-Agx9 Ion-Pairing Interaction Energy
3. Discussion
4. Materials and Methods
4.1. Peptide Synthesis and Purification
4.2. Nuclear Magnetic Resonance Spectroscopy
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Chou, P.Y.; Fasman, G.D. Conformational parameters for amino acids in helical, β-sheet, and random coil regions calculated from proteins. Biochemistry 1974, 13, 211–222. [Google Scholar] [CrossRef]
- Muñoz, V.; Serrano, L. Intrinsic secondary structure propensities of the amino acids, using statistical φ–ψ matrices: Comparison with experimental scales. Proteins 1994, 20, 301–311. [Google Scholar] [CrossRef]
- Kuo, L.-H.; Li, J.-H.; Kuo, H.-T.; Hung, C.-Y.; Tsai, H.-Y.; Chiu, W.-C.; Wu, C.-H.; Wang, W.-R.; Yang, P.-A.; Yao, Y.-C.; et al. Effect of charged amino acid side chain length at non-hydrogen bonded strand positions on β-hairpin stability. Biochemistry 2013, 52, 7785–7797. [Google Scholar] [CrossRef] [PubMed]
- Hardy, J.; Allsop, D. Amyloid deposition as the central event in the aetiology of Alzheimer’s disease. Trends Pharmacol. Sci. 1991, 12, 383–388. [Google Scholar] [CrossRef]
- Bartzokis, G.; Lu, P.H.; Mintz, J. Human brain myelination and amyloid beta deposition in Alzheimer’s disease. Alzheimers Dement. 2007, 3, 122–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scherzinger, E.; Lurz, R.; Turmaine, M.; Mangiarini, L.; Hollenbach, B.; Hasenbank, R.; Bates, G.P.; Davies, S.W.; Lehrach, H.; Wanker, E.E. Huntingtin-encoded polyglutamine expansions form amyloid-like protein aggregates in vitro and in vivo. Cell 1997, 90, 549–558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Irvine, G.B.; El-Agnaf, O.M.; Shankar, G.M.; Walsh, D.M. Protein aggregation in the brain: The molecular basis for Alzheimer’s and Parkinson’s diseases. Mol. Med. 2008, 14, 451–464. [Google Scholar] [CrossRef]
- Höppener, J.W.M.; Ahrén, B.; Lips, C.J.M. Islet amyloid and type 2 diabetes mellitus. N. Engl. J. Med. 2000, 343, 411–419. [Google Scholar] [CrossRef]
- Westermark, P.; Andersson, A.; Westermark, G.T. Islet amyloid polypeptide, islet amyloid, and diabetes mellitus. Physiol. Rev. 2011, 91, 795–826. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Tan, J.; Pei, R.; Ye, S. Acidic environment significantly alters aggregation pathway of human islet amyloid polypeptide at negative lipid membrane. Langmuir 2020, 36, 1530–1537. [Google Scholar] [CrossRef]
- Cheng, Y.-S.; Chen, Z.-T.; Liao, T.-Y.; Lin, C.; Shen, H.C.-H.; Wang, Y.-H.; Chang, C.-W.; Liu, R.-S.; Chen, R.P.-Y.; Tu, P.-H. An intranasally delivered peptide drug ameliorates cognitive decline in Alzheimer transgenic mice. EMBO Mol. Med. 2017, 9, 703–715. [Google Scholar] [CrossRef]
- Watkins, A.M.; Arora, P.S. Anatomy of β-strands at protien-protein interfaces. ACS Chem. Biol. 2014, 9, 1747–1754. [Google Scholar] [CrossRef]
- Blosser, S.L.; Sawyer, N.; Maksimovic, I.; Ghosh, B.; Arora, P.S. Covalent and noncovalent targeting of the Tcf4/β-catenin strand interface with β-hairpin mimics. ACS Chem. Biol. 2021, 16, 1518–1525. [Google Scholar] [CrossRef] [PubMed]
- Pauling, L.; Corey, R.B. The pleated sheet, a new layer configuration of polypeptide chains. Proc. Natl. Acad. Sci. USA 1951, 37, 251–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blanco, F.J.; Jimenez, M.A.; Herranz, J.; Rico, M.; Santoro, J.; Nieto, J.L. NMR evidence of a short linear peptide that folds into a β-hairpin in aqueous solution. J. Am. Chem. Soc. 1993, 115, 5887–5888. [Google Scholar] [CrossRef]
- Wouters, M.A.; Curmi, P.M. An analysis of side chain interactions and pair correlations within antiparallel β-sheets: The differences between backbone hydrogen-bonded and non-hydrogen-bonded residue pairs. Proteins 1995, 22, 119–131. [Google Scholar] [CrossRef] [PubMed]
- Cootes, A.P.; Curmi, P.M.; Cunningham, R.; Donnelly, C.; Torda, A.E. The dependence of amino acid pair correlations on structural environment. Proteins 1998, 32, 175–189. [Google Scholar] [CrossRef]
- Narayanan, C.; Dias, C.L. Hydrophobic interactions and hydrogen bonds in β-sheet formation. J. Chem. Phys. 2013, 139, 115103. [Google Scholar] [CrossRef] [Green Version]
- Searle, M.S.; Griffiths-Jones, S.R.; Skinner-Smith, H. Energetics of weak interactions in a β-hairpin peptide: Electrostatic and hydrophobic contributions to stability from lysine salt bridges. J. Am. Chem. Soc. 1999, 121, 11615–11620. [Google Scholar] [CrossRef]
- Tatko, C.D.; Waters, M.L. Selective aromatic interactions in β-hairpin peptides. J. Am. Chem. Soc. 2002, 124, 9372–9373. [Google Scholar] [CrossRef]
- Kiehna, S.E.; Waters, M.L. Sequence dependence of β-hairpin structure: Comparison of a salt bridge and an aromatic interaction. Protein Sci. 2003, 12, 2657–2667. [Google Scholar] [CrossRef] [PubMed]
- Ramirez-Alvarado, M.; Blanco, F.J.; Serrano, L. Elongation of the BH8 β-hairpin peptide: Electrostatic interactions in β-hairpin formation and stability. Protein Sci. 2001, 10, 1381–1392. [Google Scholar] [CrossRef] [PubMed]
- Kuo, H.-T.; Fang, C.-J.; Tsai, H.-Y.; Yang, M.-F.; Chang, H.-C.; Liu, S.-L.; Kuo, L.-H.; Wang, W.-R.; Yang, P.-A.; Huang, S.-J.; et al. Effect of charged amino acid side chain length on lateral cross-strand interations between carboxylate-containing residues and lysine analogues in a β-hairpin. Biochemistry 2013, 52, 9212–9222. [Google Scholar] [CrossRef] [PubMed]
- Kuo, H.-T.; Liu, S.-L.; Chiu, W.-C.; Fang, C.-J.; Chang, H.-C.; Wang, W.-R.; Yang, P.-A.; Li, J.-H.; Huang, S.-J.; Huang, S.-L.; et al. Effect of charged amino acid side chain length on lateral cross-strand interactions between carboxylate- and guanidinium-containing residues in a β-hairpin. Amino Acids 2015, 47, 885–898. [Google Scholar] [CrossRef]
- Huang, C.-H.; Wong, T.W.; Yu, C.-H.; Chang, J.-Y.; Huang, S.-J.; Huang, S.-L.; Cheng, R.P. Swapping the positions in a cross-strand lateral ion-pairing interaction between ammonium- and carboxylate-containing residues in a β-hairpin. Molecules 2021, 26, 1346. [Google Scholar] [CrossRef]
- Syud, F.A.; Stanger, H.E.; Gellman, S.H. Interstrand side chain-side chain interactions in a designed β-hairpin: Significance of both lateral and diagonal pairings. J. Am. Chem. Soc. 2001, 123, 8667–8677. [Google Scholar] [CrossRef]
- Chothia, C. Conformation of twisted β-pleated sheets in proteins. J. Mol. Biol. 1973, 75, 295–302. [Google Scholar] [CrossRef]
- Tatko, C.D.; Waters, M.L. The geometry and efficacy of cation-π interactions in a diagonal position of a designed β-hairpin. Protein Sci. 2003, 12, 2443–2452. [Google Scholar] [CrossRef]
- Hughes, R.M.; Waters, M.L. Influence of N-methylation on a cation-π interaction produces a remarkably stable β-haripin peptide. J. Am. Chem. Soc. 2005, 127, 6518–6519. [Google Scholar] [CrossRef]
- Hughes, R.M.; Waters, M.L. Arginine methylation in a β-hairpin peptide: Implications for Arg-π interactions, ΔCp°, and the cold denatured state. J. Am. Chem. Soc. 2006, 128, 12735–12742. [Google Scholar] [CrossRef]
- Hughes, R.M.; Bensoff, M.L.; Waters, M.L. Effects of Chain Length and N-Methylation on a Cation-π Interaction in a β-Hairpin Peptide. Chem. Eur. J. 2007, 13, 5753–5764. [Google Scholar] [CrossRef] [PubMed]
- Tatko, C.D.; Waters, M.L. Comparison of C-H⋯π and hydrophobic interactions in a β-hairpin peptide: Impact on stability and specificity. J. Am. Chem. Soc. 2004, 126, 2028–2034. [Google Scholar] [CrossRef] [PubMed]
- Laughrey, Z.R.; Kiehna, S.E.; Riemen, A.J.; Waters, M.L. Carbohydrate-π interactions: What are they worth? J. Am. Chem. Soc. 2008, 130, 14625–14633. [Google Scholar] [CrossRef] [Green Version]
- Kiehna, S.E.; Laughrey, Z.R.; Waters, M.L. Evaluation of a carbohydrate-π interaction in a peptide model system. Chem. Commun. 2007, 39, 4026–4028. [Google Scholar] [CrossRef]
- Chang, J.-Y.; Pan, Y.-J.; Huang, P.-Y.; Sun, Y.-T.; Yu, C.-H.; Ning, Z.-J.; Huang, S.-L.; Huang, S.-J.; Cheng, R.P. The effects of charged amino acid side-chain length on diagonal cross-strand interactions between carboxylate- and ammonium-containing residues in a β-hairpin. Molecules 2022, 27, 4172. [Google Scholar] [CrossRef] [PubMed]
- Cheng, R.P.; Weng, Y.-J.; Wang, W.-R.; Koyack, M.J.; Suzuki, Y.; Wu, C.-H.; Yang, P.-A.; Hsu, H.-C.; Kuo, H.-T.; Girinath, P.; et al. Helix formation and capping energetics of arginine analogs with varying side chain length. Amino Acids 2012, 43, 195–206. [Google Scholar] [CrossRef]
- Kuo, H.-T.; Yang, P.-A.; Wang, W.-R.; Hsu, H.-C.; Wu, C.-H.; Ting, Y.-T.; Weng, M.-H.; Kuo, L.-H.; Cheng, R.P. Effect of side chain length on intrahelical interactions between carboxylate- and guanidinium-containing amino acids. Amino Acids 2014, 46, 1867–1883. [Google Scholar] [CrossRef]
- Chang, J.-Y.; Li, N.-Z.; Wang, W.-M.; Liu, C.-T.; Yu, C.-H.; Chen, Y.-C.; Lu, D.; Lin, P.-H.; Huang, C.-H.; Kono, O.; et al. Longer charged amino acids favor β-strand formation in hairpin peptides. J. Pept. Sci. 2021, 27, e3333. [Google Scholar] [CrossRef]
- Stanger, H.E.; Gellman, S.H. Rules for antiparallel β-sheet design: D-Pro-Gly is superior to L-Asn-Gly for β-hairpin nucleation. J. Am. Chem. Soc. 1998, 120, 4236–4237. [Google Scholar] [CrossRef]
- Atherton, E.; Fox, H.; Harkiss, D.; Logan, C.J.; Sheppard, R.C.; Williams, B.J. A mild procedure for solid phase peptide synthesis: Use of fluorenylmethoxycarbonylamino-acids. J. Chem. Soc. Chem. Commun. 1978, 13, 537–539. [Google Scholar] [CrossRef]
- Fields, G.B.; Noble, R.L. Solid phase peptide synthesis utilizing 9-fluorenylmethoxycarbonyl amino acids. Int. J. Pept. Protein Res. 1990, 35, 161–214. [Google Scholar] [CrossRef] [PubMed]
- Feichtinger, K.; Sings, H.L.; Baker, T.J.; Matthews, K.; Goodman, M. Triurethane-protected guanidines and triflyldiurethane-protected guanidines: New reagents for guanidinylation reactions. J. Org. Chem. 1998, 63, 8432–8439. [Google Scholar] [CrossRef]
- Volkmer-Engert, R.; Landgraf, C.; Schneider-Mergener, J. Charcoal surface-assisted catalysis of intramolecular disulfide bond formation in peptides. J. Pept. Res. 1998, 51, 365–369. [Google Scholar] [CrossRef] [PubMed]
- Bax, A.; Davis, D.G. MLEV-17-based two-dimensional homonuclear magnetization transfer spectroscopy. J. Magn. Reson. 1985, 65, 355–360. [Google Scholar] [CrossRef]
- Aue, W.P.; Bartholdi, E.; Ernst, R.R. Two-dimensional spectroscopy. Application to nuclear magnetic resonance. J. Chem. Phys. 1976, 64, 2229–2246. [Google Scholar] [CrossRef] [Green Version]
- Jeener, J.; Meier, B.H.; Bachmann, P.; Ernst, R.R. Investigation of exchange processes by two-dimensional NMR spectroscopy. J. Chem. Phys. 1979, 71, 4546–4553. [Google Scholar] [CrossRef]
- Ramirez-Alvarado, M.; Blanco, F.J.; Serrano, L. De novo design and structural analysis of a model β-hairpin peptide system. Nat. Struct. Mol. Biol. 1996, 3, 604–612. [Google Scholar] [CrossRef]
- Russell, S.J.; Blandl, T.; Skelton, N.J.; Cochran, A.G. Stability of cyclic β-hairpins: Asymmetric contributions from side chains of a hydrogen-bonded cross-strand residue pair. J. Am. Chem. Soc. 2003, 125, 388–395. [Google Scholar] [CrossRef]
- Kim, Y.; Prestegard, J.H. Measurement of vicinal couplings from cross peaks in COSY spectra. J. Magn. Reson. 1989, 84, 9–13. [Google Scholar] [CrossRef]
- Ciani, B.; Jourdan, M.; Searle, M.S. Stabilization of β-hairpin peptides by salt bridges: Role of preorganization in the energetic contribution of weak interactions. J. Am. Chem. Soc. 2003, 125, 9038–9047. [Google Scholar] [CrossRef]
- Horovitz, A. Double-mutant cycles: A powerful tool for analyzing protein structure and function. Fold. Des. 1996, 1, R121–R126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yao, J.; Dyson, H.J.; Wright, P.E. Chemical shift dispersion and secondary structure prediction in unfolded and partly folded proteins. FEBS Lett. 1997, 419, 285–289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bothner-By, A.A.; Stephens, R.L.; Lee, J.; Warren, C.D.; Jeanloz, R.W. Structure determination of a tetrasaccharide: Transient nuclear Overhauser effects in the rotating frame. J. Am. Chem. Soc. 1984, 106, 811–813. [Google Scholar]
- Piotto, M.; Saudek, V.; Sklenar, V. Gradient-tailored excitation for single-quantum NMR spectroscopy of aqueous solutions. J. Biomol. NMR 1992, 2, 661–665. [Google Scholar]
- Sklenar, V.; Piotto, M.; Leppik, R.; Saudek, V. Gradient-tailored water suppression for 1H-15N HSQC experiments optimized to retain full sensitivity. J. Magn. Reson. Ser. A 1993, 102, 241–245. [Google Scholar]
- Syud, F.A.; Espinosa, J.F.; Gellman, S.H. NMR-based quantification of β-sheet populations in aqueous solution through use of reference peptides for the folded and unfolded states. J. Am. Chem. Soc. 1999, 121, 11577–11578. [Google Scholar]



{kind=link}
{kind=link}
| Zbb2 | Agx9 | |||
|---|---|---|---|---|
| Agp | Agb | Arg | Agh | |
| Asp | 21 ± 4 | 19 ± 4 | 17 ± 5 | 16 ± 6 |
| Glu | 31 ± 2 | 41 ± 4 | 36 ± 4 | 35 ± 4 |
| Aad | 30 ± 3 | 51 ± 4 | 37 ± 5 | 39 ± 5 |
| Zbb2 | Agx9 | |||
|---|---|---|---|---|
| Agp | Agb | Arg | Agh | |
| Asp | 0.789 ± 0.143 | 0.869 ± 0.163 | 0.942 ± 0.212 | 0.993 ± 0.261 |
| Glu | 0.473 ± 0.058 | 0.229 ± 0.098 | 0.334 ± 0.110 | 0.383 ± 0.103 |
| Aad | 0.496 ± 0.094 | −0.020 ± 0.099 | 0.332 ± 0.123 | 0.272 ± 0.113 |
| Zbb2 | Agx9 | |||
|---|---|---|---|---|
| Agp | Agb | Arg | Agh | |
| Asp | −0.453 ± 0.227 | −0.074 ± 0.208 | −0.185 ± 0.258 | −0.061 ± 0.124 |
| Glu | −0.196 ± 0.055 | −0.141 ± 0.070 | −0.221 ± 0.145 | −0.098 ± 0.042 |
| Aad | 0.061 ± 0.056 | −0.156 ± 0.045 | 0.011 ± 0.172 | 0.026 ± 0.054 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, N.-Z.; Yu, C.-H.; Wu, J.-Y.; Huang, S.-J.; Huang, S.-L.; Cheng, R.P. Diagonal Interactions between Glutamate and Arginine Analogs with Varying Side-Chain Lengths in a β-Hairpin. Molecules 2023, 28, 2888. https://doi.org/10.3390/molecules28072888
Li N-Z, Yu C-H, Wu J-Y, Huang S-J, Huang S-L, Cheng RP. Diagonal Interactions between Glutamate and Arginine Analogs with Varying Side-Chain Lengths in a β-Hairpin. Molecules. 2023; 28(7):2888. https://doi.org/10.3390/molecules28072888
Chicago/Turabian StyleLi, Nian-Zhi, Chen-Hsu Yu, Jhuan-Yu Wu, Shing-Jong Huang, Shou-Ling Huang, and Richard P. Cheng. 2023. "Diagonal Interactions between Glutamate and Arginine Analogs with Varying Side-Chain Lengths in a β-Hairpin" Molecules 28, no. 7: 2888. https://doi.org/10.3390/molecules28072888
APA StyleLi, N. -Z., Yu, C. -H., Wu, J. -Y., Huang, S. -J., Huang, S. -L., & Cheng, R. P. (2023). Diagonal Interactions between Glutamate and Arginine Analogs with Varying Side-Chain Lengths in a β-Hairpin. Molecules, 28(7), 2888. https://doi.org/10.3390/molecules28072888