Anti-Trypanosomal Bufadienolides from the Oocytes of the Toad Rhinella alata (Anura, Bufonidae)


 , , ,
, , ,
Abstract
:
1. Introduction
2. Results and Discussion
2.1. Isolation and Structural Elucidation of Bufadienolides
2.2. Anti-Trypanosomal Activity and Mammalian Cytotoxicity
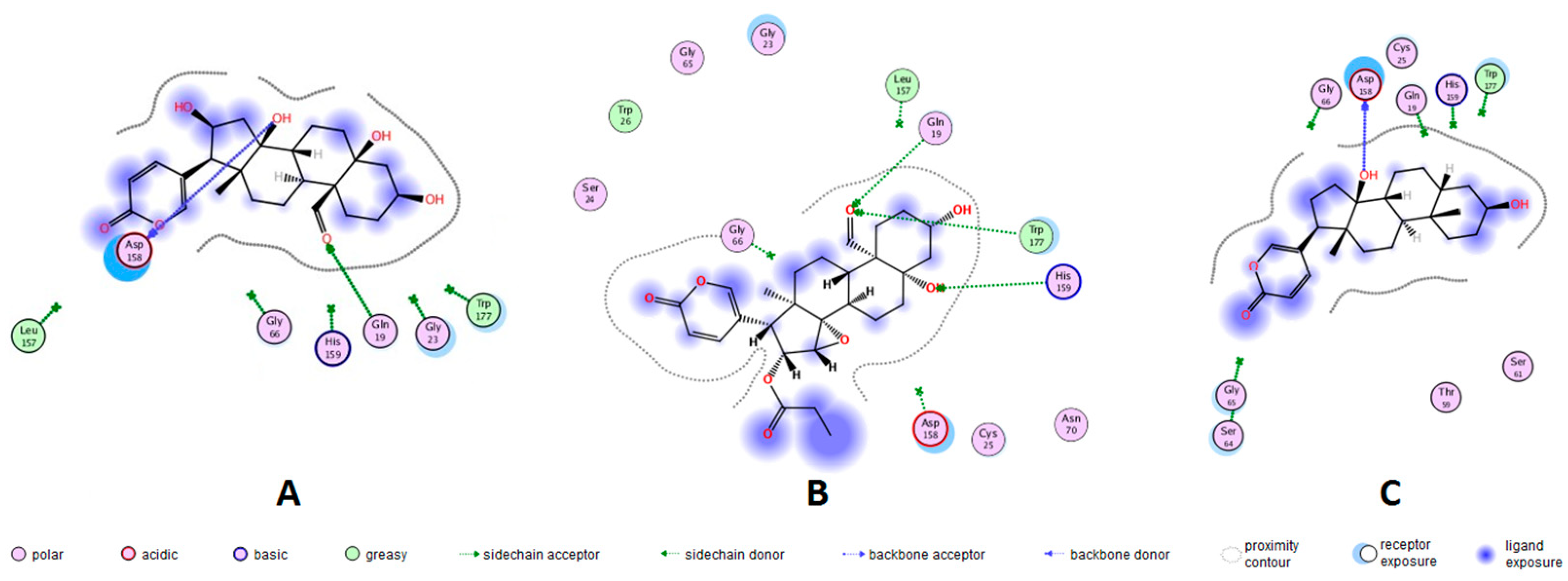
2.3. Molecular Docking of Bufadienolides 1, 3 and 7
3. Materials and Methods
3.1. General Experimental Procedures
3.2. Sample Collection
3.3. Extraction and Isolation of Bufadienolides
3.4. Compounds Characterization Data
3.5. Anti-Trypanosomal Activity
3.6. Cytotoxicity Assay
3.7. Statistical Analysis of Bioassays
3.8. Molecular Docking
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Steverding, D. The history of Chagas disease. Parasites Vectors 2014, 7, 317. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control (CDC)—Chagas Disease. Available online: https://www.cdc.gov/parasites/chagas/index.html (accessed on 21 December 2020).
- Pérez-Molina, J.P.; Molina, I. Chagas disease. Lancet 2017, 391, 82–94. [Google Scholar] [CrossRef] [PubMed]
- Sueth-Santiago, V.; Decote-Ricardo, D.; Morrot, A.; Freire-de-Lima, C.G.; Freire-Lima, M.E. Challenges in the chemotherapy of Chagas disease: Looking for possibilities related to the differences and similarities between the parasite and host. World J. Biol. Chem. 2017, 8, 57. [Google Scholar] [CrossRef] [PubMed]
- Verpoorte, R.; Phax-quôc-Kinh; Svendsen, A.B. Chemical constituents of Vietnamese toad venom, collected from Bufo Melanostictus Schneider. Part II. The bufadienolides. J. Nat. Prod. 1980, 43, 347–352. [Google Scholar] [CrossRef]
- Sousa, L.Q.; Machado, K.C.; Carvalho, O.S.F.; Silva, A.L.; Moncao-Filho, E.S.; Melo-Cavalcante, A.A.; Vieira-Júnior, G.M.; Pinheiro, F.P.M. Bufadienolides from amphibians: A promising source of anticancer prototypes for radical innovation, apoptosis triggering and Na+/K+-ATPase inhibition. Toxicon 2017, 127, 63–76. [Google Scholar] [CrossRef] [PubMed]
- Steyn, P.S.; Van Heerden, F.R. Bufadienolides of plant and animal origin. Nat. Prod. Rep. 1998, 15, 397–413. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, C.; Rollins-smith, L.; Ibáñez, R.; Durant-Archibold, A.A.; Gutierrez, M. Toxins and pharmacologically active compounds from species of the family Bufonidae (Amphibia, Anura). J. Ethnopharmacol. 2017, 198, 235–254. [Google Scholar] [CrossRef] [PubMed]
- Santos, S.P.; Ibáñez, R.; Ron, S.R. Systematics of the Rhinella margaritifera complex (Anura, Bufonidae) from western Ecuador and Panama with insights in the biogeography of Rhinella alata. Zookeys 2015, 145, 109–145. [Google Scholar] [CrossRef]
- Xiao, J.; Zhao, X.; Zhong, W.T.; Jiao, F.R.; Wang, X.L.; Ma, L.; Duan, D.Z.; Yang, D.S.; Tang, S.Q. Bufadienolides from the venom of Bufo gargarizans and their enzyme inhibition activities and brine shrimp lethality. Nat. Prod. Commun. 2018, 13, 827–830. [Google Scholar] [CrossRef]
- Krenn, L.; Stapf, V.; Kopp, B. Bufadienolides from Drimia robusta BAK. Sci. Pharm. 2000, 68, 421–427. [Google Scholar] [CrossRef]
- Kamano, Y.; Nogawa, T.; Yamashita, A.; Pettit, G.R. The 1H and 13C NMR chemical shift assignments for thirteen bufadienolides isolated from the traditional Chinese drug Ch’an Su. Collect. Czechoslov. Chem. Commun. 2001, 66, 1841–1848. [Google Scholar] [CrossRef]
- Ondetti, M.A.; Williams, N.J.; Sabo, E.; Josip, P.F.; Weaver, E.R.; Kocy, O. Angiotensin-converting enzyme inhibitors from the venom of Bothrops jararaca. Isolation, elucidation of structure, and synthesis. Biochemistry 1971, 10, 4033–4039. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, T.; Katsuyama, S.; Orito, T.; Suzuki, T.; Sakurada, S. Antinociceptive effect of tebanicline for various noxious stimuli-induced behaviours in mice. Neurosci. Lett. 2017, 638, 46–50. [Google Scholar] [CrossRef]
- Spande, T.F.; Garrafo, H.M.; Edwards, M.W.; Yeh, H.J.C.; Pannell, L.; Daly, J.W. Epibatidine: A novel (Chloropyridy1)azabicycloheptane with potent analgesic activity from an Ecuadorian poison frog. J. Am. Chem. Soc. 1992, 114, 3475–3478. [Google Scholar] [CrossRef]
- Gao, H.; Zehl, M.; Leitner, A.; Wu, X.; Wang, Z.; Kopp, B. Comparison of toad venoms from different Bufo species by HPLC and LC-DAD-MS/MS. J. Ethnopharmacol. 2010, 131, 368–376. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.W.; Tian, H.Y.; Nie, Q.L.; Wang, L.; Zhou, S.W.; Ye, W.C.; Zhang, D.M.; Jiang, R.W. Structures and inhibitory activity against breast cancer cells of new bufadienolides from the eggs of toad: Bufo bufo gargarizans. RSC Adv. 2016, 6, 93832–93841. [Google Scholar] [CrossRef]
- Servillon, R.J.T.; Dingal, M.C.; Lusica, M.C.; Yamson, M.K.A.; Balonebro, M.J.B.; Alzate, F.B. Phytochemical screening and in vitro antibacterial evaluation of Persea americana (Avocado) crude peel extract. Optima 2013, 1, 96. [Google Scholar]
- Tempone, A.G.; Pimenta, D.C.; Lebrun, I.; Sartorelli, P.; Taniwaki, N.N.; De Andrade, H.F., Jr.; Antoniazzi, M.M.; Jared, C. Antileishmanial and anti-trypanosomal activity of bufadienolides isolated from the toad Rhinella jimi parotoid macrogland secretion. Toxicon 2008, 52, 13–21. [Google Scholar] [CrossRef]
- Liu, J.; Zhang, D.; Li, Y.; Chen, W.; Ruan, Z.; Deng, L.; Wang, L.; Tian, H.; Yiu, A.; Fan, C.; et al. Discovery of bufadienolides as a novel class of ClC-3 chloride channel activators with antitumor activities. J. Med. Chem. 2013, 56, 5734–5743. [Google Scholar] [CrossRef]
- Rodriguez, C.; Ibáñez, R.; NG, M.; Spadafora, C.; Durant-Archibold, A.A.; Gutiérrez, M. 19-Hydroxy-bufalin, a major bufadienolide isolated from the parotoid gland secretions of the Panamanian endemic toad Rhinella centralis (Bufonidae), inhibits the growth of Trypanosoma cruzi. Toxicon 2020, 177, 89–92. [Google Scholar] [CrossRef]
- Córdova, W.H.P.; Leitao, S.G.; Cunha-Filho, G.; Bosch, R.A.; Pascual, I.A.; Pereda-Miranda, R.; Gervou, R.; Arújo, T.N.; Quintas, M.L.E.; Noel, F. Bufadienolides from the parotoid gland secretions of Cuban toad Peltophryne fustiger (Bufonidae): Inhibition of human (Na+/K+)-ATPase activity. Toxicon 2016, 110, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Van Der Heyden, N.; Docampo, R. Proton and sodium pumps regulate the plasma membrane potential of different stages of Trypanosoma cruzi. Mol. Biochem. Parasitol. 2002, 120, 127–139. [Google Scholar] [CrossRef] [PubMed]
- Kyoichi, I.; Mikami, Y.; Hashimoto, M.; Nara, T.; Hara, Y.; Aoki, T. Molecular cloning and characterization of ouabain-insensitive Na+-ATPase in the parasitic protist, Trypanosoma cruzi. Biochim. Biophys. Acta 2006, 1758, 738–746. [Google Scholar] [CrossRef]
- Kamano, Y.; Kotake, A.; Hashima, H.; Inoue, M.; Morita, H.; Takeya, K.; Itokawa, H.; Nandachi, N.; Segawa, T.; Yukita, A.; et al. Structure-cytotoxic activity relationship for the toad poison bufadienolides. Bioorg. Med. Chem. 1998, 6, 1103–1115. [Google Scholar] [CrossRef] [PubMed]
- Silva-Junior, E.F.; Barcellos, F.P.H.; Ribeiro, F.F.; Bezerra, M.J.; Scotti, L.; Tullius, S.M.; De Aquino, T.M.; De Araujo-Junior, J.X. Molecular docking studies applied to a dataset of cruzain inhibitors. Curr. Comput. Aided. Drug Des. 2017, 14, 68–78. [Google Scholar] [CrossRef] [PubMed]
- Sajid, M.; Robertson, S.A.; Brinen, L.S.; McKerrow, J.H. Cruzain: The path from target validation to the clinic. Adv. Exp. Med. Biol. 2011, 712, 100–115. [Google Scholar] [CrossRef]
- Nogueira, R.C.; Oliveira-Costa, J.F.; De Sá, M.S.; Dos Santos, R.R.; Soares, M.B.P. Early toxicity screening and selection of lead compounds for parasitic diseases. Curr. Drug Targets 2009, 10, 291–298. [Google Scholar] [CrossRef]
- Calderón-Montaño, J.; Burgos-Morón, E.; López-Lázaro, M. The in vivo antitumor activity of cardiac glycosides in mice xenografted with human cancer cells is probably an experimental artifact. Oncogene 2014, 33, 2947–2948. [Google Scholar] [CrossRef]
- Lu, J.; Deng, S.; Chen, H.; Hou, J.; Zhang, B.; Tian, Y.; Wang, C.; Ma, X. Microbial transformation of cinobufotalin by Alternaria alternate AS 3.4578 and Aspergillus niger AS 3.739. J. Mol. Catal. B Enzym. 2013, 89, 102–107. [Google Scholar] [CrossRef]
- Xiu-Lan, X.; Li-Bin, Z.; Feng-Yun, L.; Xiao-Chi, M.; Ke-Xin, L.; Jian, H.; De-An, G. Microbial transformation of bufotalin by Alternaria alternata AS 3.4578. J. Asian Nat. Prod. Res. 2009, 11, 7–11. [Google Scholar] [CrossRef]
- Ye, M.; Han, J.; Tu, G.; An, D.; Guo, D. Microbial hydroxylation of bufalin by Cunninghamella blakesleana and Mucor spinosus. J. Nat. Prod. 2005, 68, 626–628. [Google Scholar] [CrossRef] [PubMed]
- Yin, P.; Wang, Y.; Qiu, Y.Y.; Hou, L.; Liu, X.; Qin, J.; Duan, Y.; Liu, P.; Qiu, M.; Li, Q. Bufalin-loaded mPEG-PLGA-PLL-cRGD nanoparticles: Preparation, cellular uptake, tissue distribution, and anticancer activity. Int. J. Nanomed. 2012, 7, 3961–3969. [Google Scholar] [CrossRef]
- Barnhart, K.; Forman, M.E.; Umile, T.P.; Kuenemn, J.; McKenzie, V.; Salinas, I.; Minbiole, K.P.C.; Woodhams, D.C. Identification of bufadienolides from the boreal toad, Anaxyrus boreas, active against a fungal pathogen. Microb. Ecol. 2017, 74, 990–1000. [Google Scholar] [CrossRef] [PubMed]
- Densmore, C.L.; Green, D.E. Diseases of Amphibians. Inst. fo Lab. Anim. Res. J. 2007, 48, 235–254. [Google Scholar] [CrossRef]
- Gupta, D.K.; Gupta, N.; Gangwar, R. Infectivity of Bufo melanostictus (Amphibia: Bufonidae) to two new species of haematozoan parasites from Rohilkhand. India. Proc. Zool. Soc. 2012, 65, 22–32. [Google Scholar] [CrossRef]
- Ferreira, J.I.G.S.; Da Costa, A.P.; Ramirez, D.; Roldan, J.A.M.; Saraiva, D.; Founier, S.G.F.R.; Sue, A.; Zambelli, E.R.; Minervino, H.A.H.; Verdade, K.V.; et al. Anuran trypanosomes: Phylogenetic evidence for new clades in Brazil. Syst. Parasitol. 2015, 91, 63–70. [Google Scholar] [CrossRef]
- Bernal, X.E.; Pinto, C.M. Parasites and wildlife sexual differences in prevalence of a new species of trypanosome infecting túngara frogs. Int. J. Parasitol. Parasites Wildl. 2016, 5, 40–47. [Google Scholar] [CrossRef]
- Barta, J.R.; Desser, S.S. Blood parasites of amphibians from Algonquin Park, Ontario. J. Wildl. Dis. 1984, 20, 180–189. [Google Scholar] [CrossRef]
- Garraffo, H.M.; Gras, E.G. Biosynthesis of bufadienolides in toads VI. Experiments with [1,2-3H] cholesterol, [21-14C] coprostanol, and 5-beta-[21-14C] pregnonolone in the toad Bufo arenarum. Steroids 1986, 48, 251–257. [Google Scholar] [CrossRef]
- Akizawa, T.; Mukai, T.; Matsukawa, M.; Yoshioka, M.; Morris, J.F.; Butler, V.P. Structures of novel bufadienolides in the eggs of a toad, Bufo marinus. Chem. Pharm. Bull. 1994, 42, 754–756. [Google Scholar] [CrossRef]
- Lichtstein, D.; Gati, I.; Haver, E.; Katz, U. Digitalis-like compounds in the toad Bufo viridis: Tissue and plasma levels and significance in osmotic stress. Life Sci. 1992, 51, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.S.; Derguini, F.; Bruening, R.C.; Nakanishi, K.; Wallick, E.T.; Akizawa, T.; Rosenbaum, C.S.; Butler, V.P. Digitalis-like compounds of toad bile: Sulfation and reduction of bufadienolides decrease potency of Na+, K+-ATPase inhibition. Heterocycles 1994, 39, 669–686. [Google Scholar]
- Flier, J.; Edwards, M.; Daly, J.W.; Myers, C. Widespread occurrence in frogs and toads of skin compounds interacting with the ouabain site of Na+, K+-ATPase. Science 1980, 208, 503–505. [Google Scholar] [CrossRef] [PubMed]
- Buckner, F.S.; Verlinde, C.L.M.J.; Flamme, A.C.L.A.; Van Voorhis, W.C. Efficient technique for screening drugs for activity against Trypanosoma cruzi using parasites expressing beta-galactosidase. Antimicrob. Agents Chemother. 1996, 40, 2592–2597. [Google Scholar] [CrossRef] [PubMed]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Brinen, L.S.; Gillmor, S.A.; Fletterick, R.J. RCSB PDB—1EWO: The Cysteine Protease Cruzain Bound to WRR-204. Available online: https://www.rcsb.org/structure/1EWO (accessed on 7 January 2021).
- Gillmor, S.A.; Craik, C.S.; Fletterick, R.J. Structural determinants of specificity in the cysteine protease cruzain. Protein Sci. 1997, 6, 1603–1611. [Google Scholar] [CrossRef]
- Choe, Y.; Brinen, L.S.; Price, M.S.; Engel, J.C.; Lange, M.; Grisostomi, C.; Weston, S.G.; Pallai, P.V.; Cheng, H.; Hardy, L.W.; et al. Development of alpha-keto-based inhibitors of cruzain, a cysteine protease implicated in Chagas disease. Bioorganic Med. Chem. 2005, 13, 2141–2156. [Google Scholar] [CrossRef]
- Jones, B.D.; Tochowicz, A.; Tang, Y.; Cameron, M.D.; McCall, L.-I.; Hirata, K.; Siqueira-Neto, J.L.; Reed, S.L.; McKerrow, J.H.; Roush, W.R. Synthesis and evaluation of oxyguanidine analogues of the cysteine protease inhibitor WRR-483 against cruzain. Med. Chem. Lett. 2016, 7, 77–82. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| No. | 13C, Type | 1H, Mult. (J in Hz) |
|---|---|---|
| 1a | 18.6, CH2 | 2.26, m |
| 1b | 1.77, m | |
| 2a | 27.5, CH2 | 1.77, m |
| 2b | 1.77, m | |
| 3 | 68.0, CH | 4.15, m |
| 4a | 36.7, CH2 | 1.99, m |
| 4b | 1.61, m | |
| 5 | 75.8, C | |
| 6a | 38.3, CH2 | 2.23, m |
| 6b | 1.49, m | |
| 7a | 24.1, CH2 | 1.77, m |
| 7b | 1.11, m | |
| 8 | 43.1, CH | 1.77, m |
| 9 | 34.4, CH | 2.47, td (3.8, 12.2) |
| 10 | 56.0, C | |
| 11a | 22.9, CH2 | 1.66, m |
| 11b | 1.39, m | |
| 12a | 40.5, CH2 | 1.77, m |
| 12b | 1.49, m | |
| 13 | 46.0, C | |
| 14 | 73.1, C | |
| 15 | 60.7, CH | 3.75, m |
| 16 | 76.3, CH | 5.47, m |
| 17 | 51.2, CH | 2.93, d (9.2) |
| 18 | 17.3, CH3 | 0.81, s |
| 19 | 210.1, CH | 10.00 s |
| 20 | 118.2, C | |
| 21 | 153.6, CH | 7.36, m |
| 22 | 150.9, CH | 8.02, m |
| 23 | 114.2, CH | 6.24, dd (0.8, 9.7) |
| 24 | 164.0, C=O | |
| 1’ | 174.9, C=O | |
| 2’a | 28.1, CH2 | 2.23, m |
| 2’b | 2.11, m | |
| 3’ | 9.3, CH3 | 0.94, t (7.8) |
| Compound | T. cruzi | VERO CELL | Selectivity (CC50/EC50) |
|---|---|---|---|
| EC50 (µM) | CC50 (µM) | ||
| 1 | 22.80 ± 4.11 | >135 | >5 |
| 2 | Inactive | >28 | ---- |
| 3 | 0.19 ± 0.07 | 0.22 ± 0.09 | 1.16 |
| 4 | 19.60 ± 2 | >46 | >2 |
| 5 | 27.80 ± 3.77 | 1.88 ± 0.20 | 0.07 |
| 6 | 12.30 ± 1.87 | 0.02 ± 0.002 | <0.01 |
| 7 | 3.93 ± 0.57 | 10.61 ± 1.54 | 2.69 |
| Benznidazole | 2.73 ± 0.10 | 51.36 ± 6.0 | 18.81 |
| Doxorubicin | Not tested | 0.23 | ---- |
| Bufadienolide | RMSD (Å) | Docking Score | Ligand Group | Amino Acid Residue | Energy (kcal/mol) | Distance (Å) |
|---|---|---|---|---|---|---|
| 1 | 2.5812 | −4.2777 | CH=O at 19 | GLN-19 | −1.5 | 2.32 |
| OH at 14 | ASP-158 | −1.7 | 2.10 | |||
| 3 | 2.0099 | −5.3984 | CH=O at 19 | GLN-19 | −2.2 | 2.09 |
| CH=O at 19 | TRP-177 | −0.5 | 2.59 | |||
| OH at 5 | HIS-159 | −1.5 | 2.35 | |||
| 7 | 3.0381 | −4.1206 | OH at 14 | ASP-158 | +1.3 | 1.95 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodriguez, C.; Ibáñez, R.; Olmedo, D.A.; Ng, M.; Spadafora, C.; Durant-Archibold, A.A.; Gutiérrez, M. Anti-Trypanosomal Bufadienolides from the Oocytes of the Toad Rhinella alata (Anura, Bufonidae). Molecules 2024, 29, 196. https://doi.org/10.3390/molecules29010196
Rodriguez C, Ibáñez R, Olmedo DA, Ng M, Spadafora C, Durant-Archibold AA, Gutiérrez M. Anti-Trypanosomal Bufadienolides from the Oocytes of the Toad Rhinella alata (Anura, Bufonidae). Molecules. 2024; 29(1):196. https://doi.org/10.3390/molecules29010196
Chicago/Turabian StyleRodriguez, Candelario, Roberto Ibáñez, Dionisio A. Olmedo, Michelle Ng, Carmenza Spadafora, Armando A. Durant-Archibold, and Marcelino Gutiérrez. 2024. "Anti-Trypanosomal Bufadienolides from the Oocytes of the Toad Rhinella alata (Anura, Bufonidae)" Molecules 29, no. 1: 196. https://doi.org/10.3390/molecules29010196
APA StyleRodriguez, C., Ibáñez, R., Olmedo, D. A., Ng, M., Spadafora, C., Durant-Archibold, A. A., & Gutiérrez, M. (2024). Anti-Trypanosomal Bufadienolides from the Oocytes of the Toad Rhinella alata (Anura, Bufonidae). Molecules, 29(1), 196. https://doi.org/10.3390/molecules29010196