
Analysis of Polyphenol Extract from Hazel Leaf and Ameliorative Efficacy and Mechanism against Hyperuricemia Zebrafish Model via Network Pharmacology and Molecular Docking

Abstract
:1. Introduction
2. Results
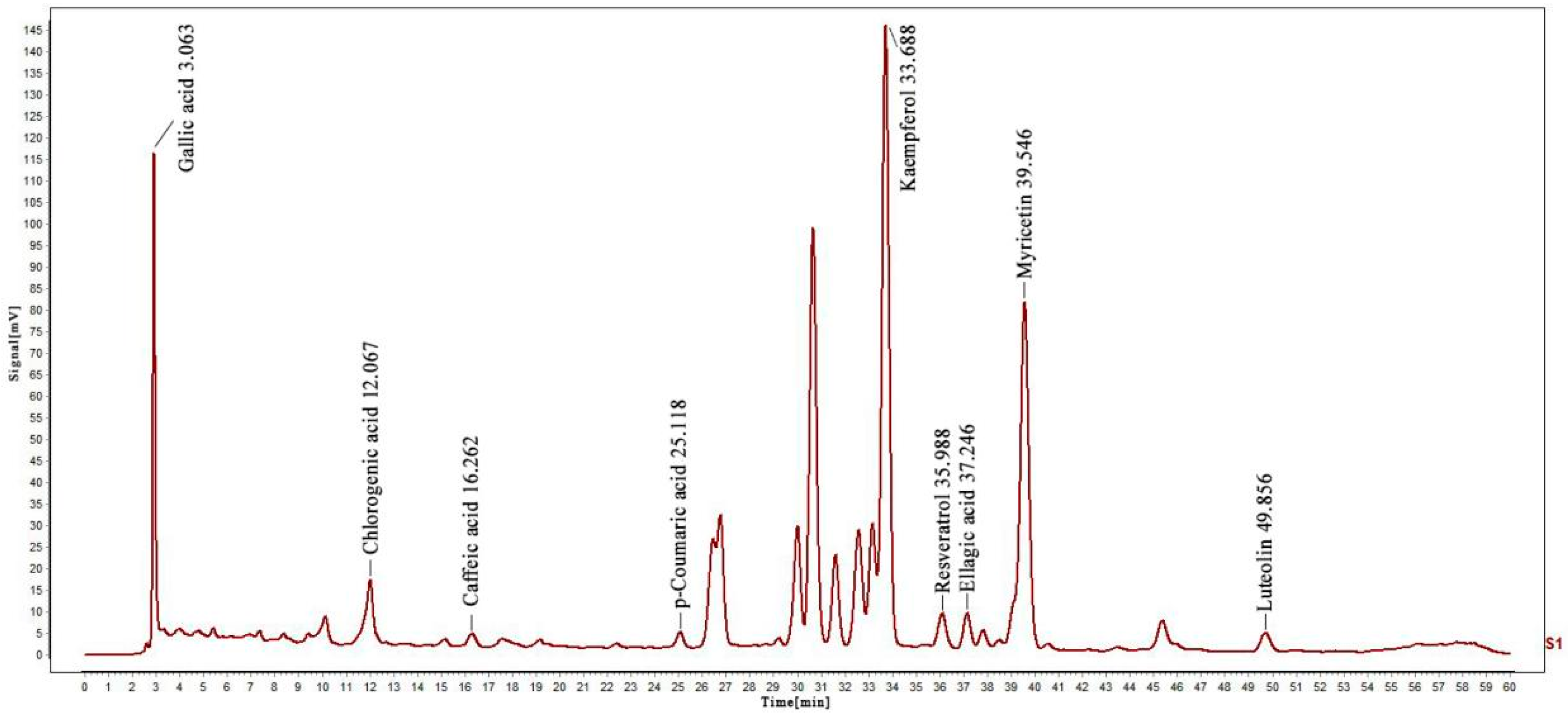
2.1. HPLC Analysis
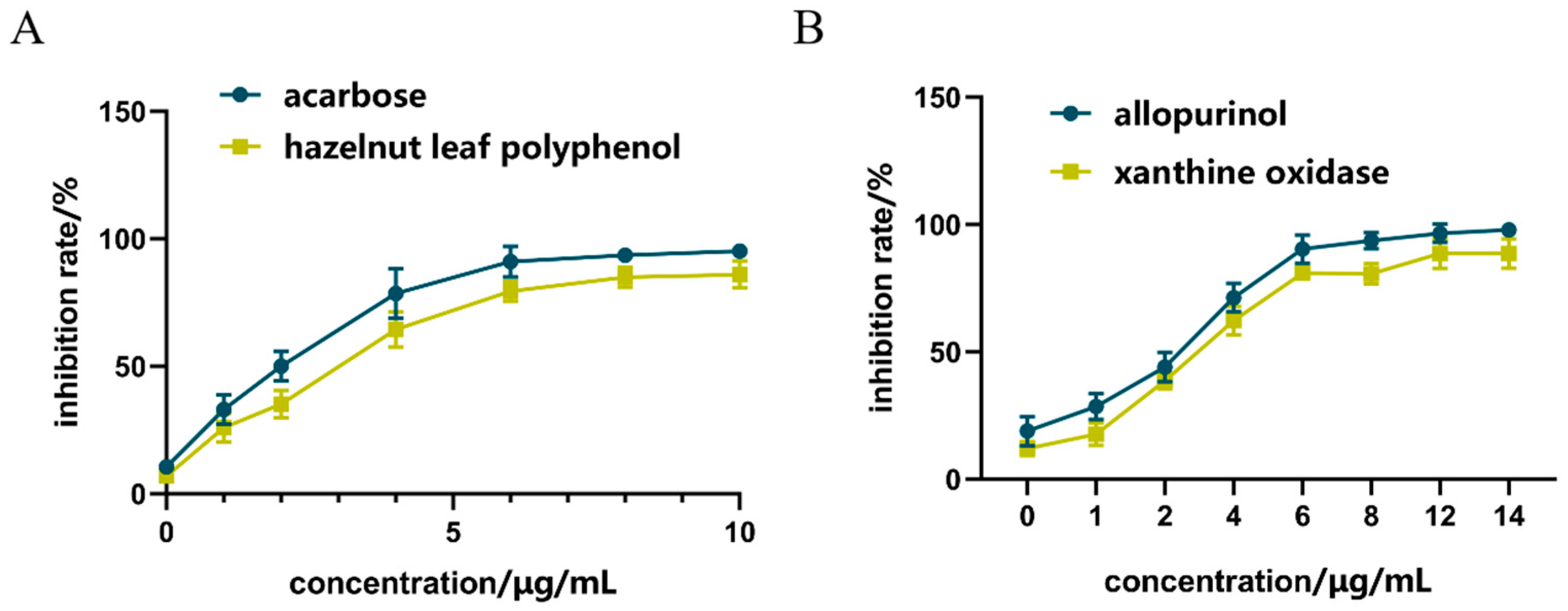
2.2. Inhibition of α-GLU
2.3. Inhibition of XOD
2.4. Effects of ZP on Hyperuricemia Zebrafish
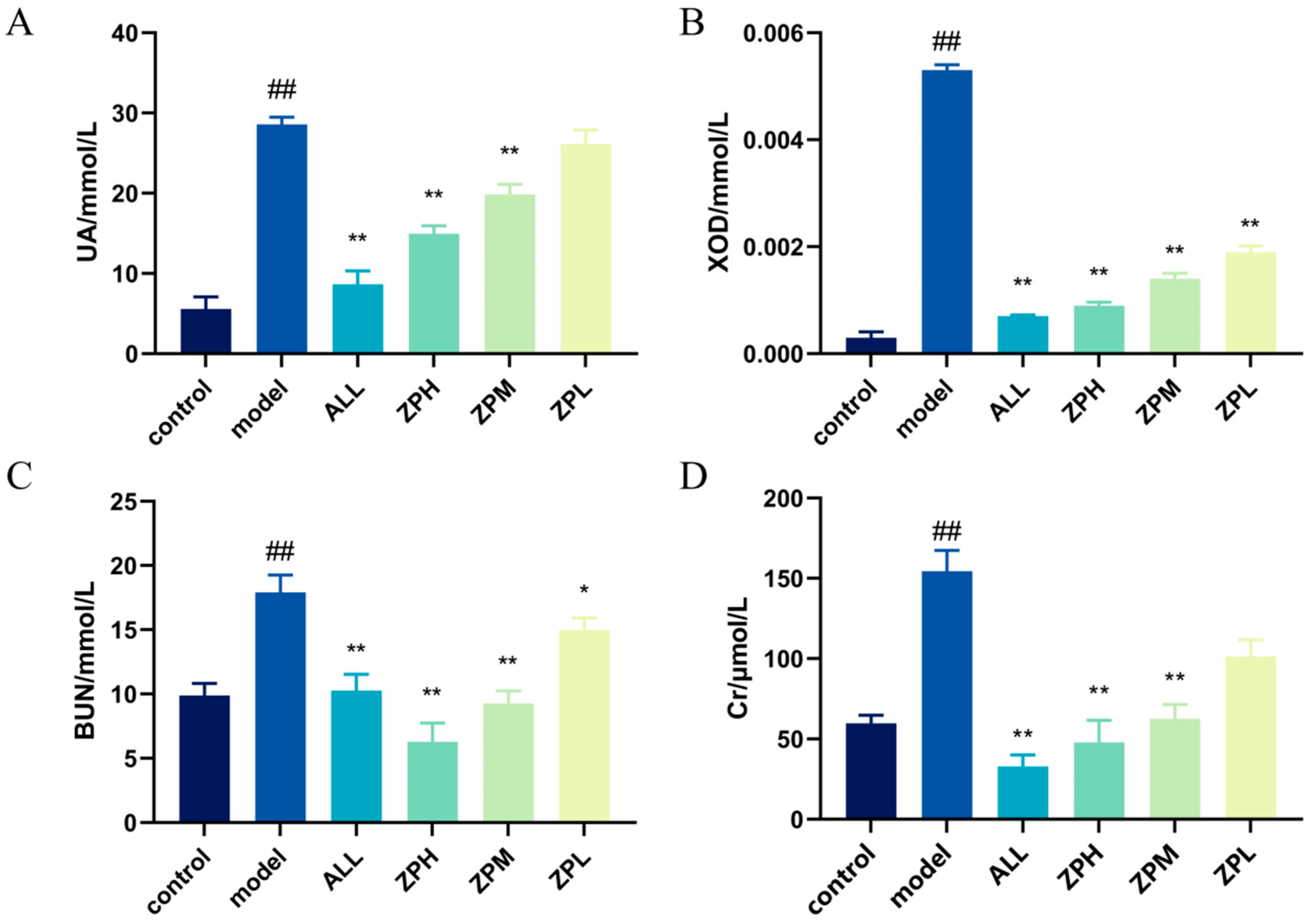
2.4.1. Effects of ZP on UA and XOD in Hyperuricemia Zebrafish
2.4.2. Effects of ZP on BUN and Cr in Hyperuricemia Zebrafish
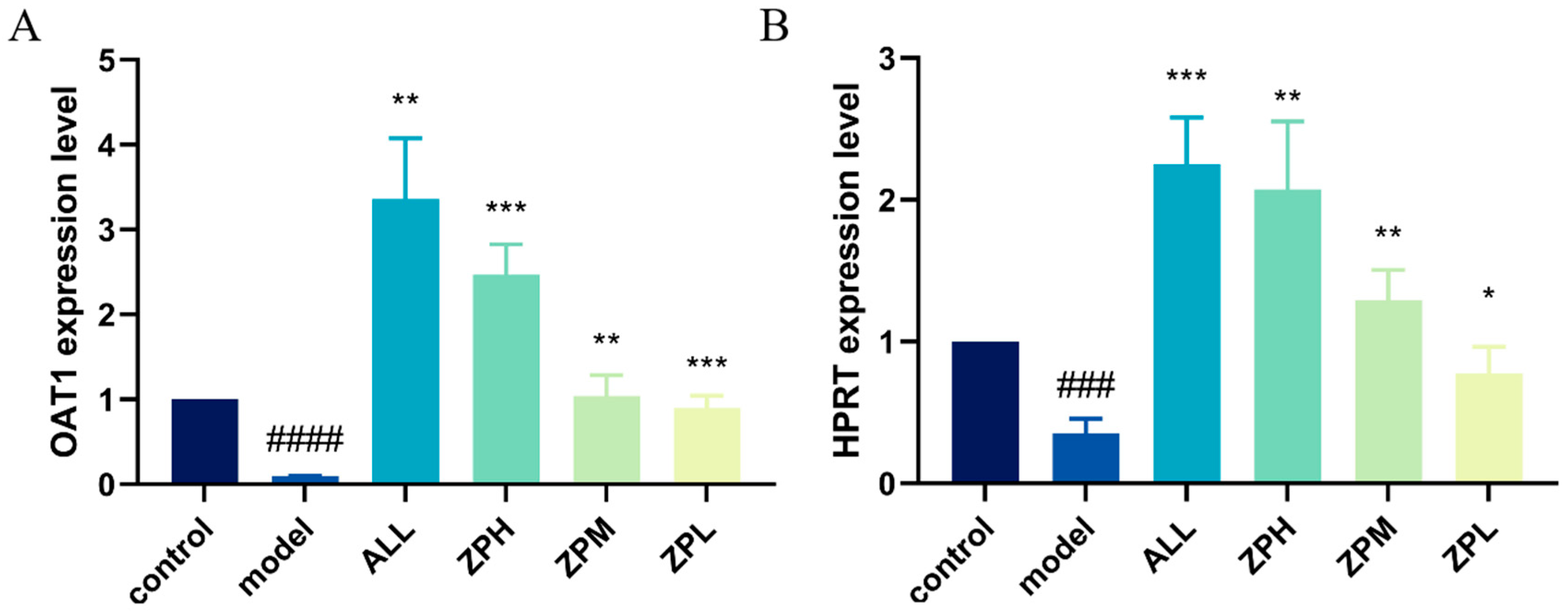
2.4.3. Effects of ZP on the Expression of Genes OAT1 and HPRT1 in Hyperuricemia Zebrafish
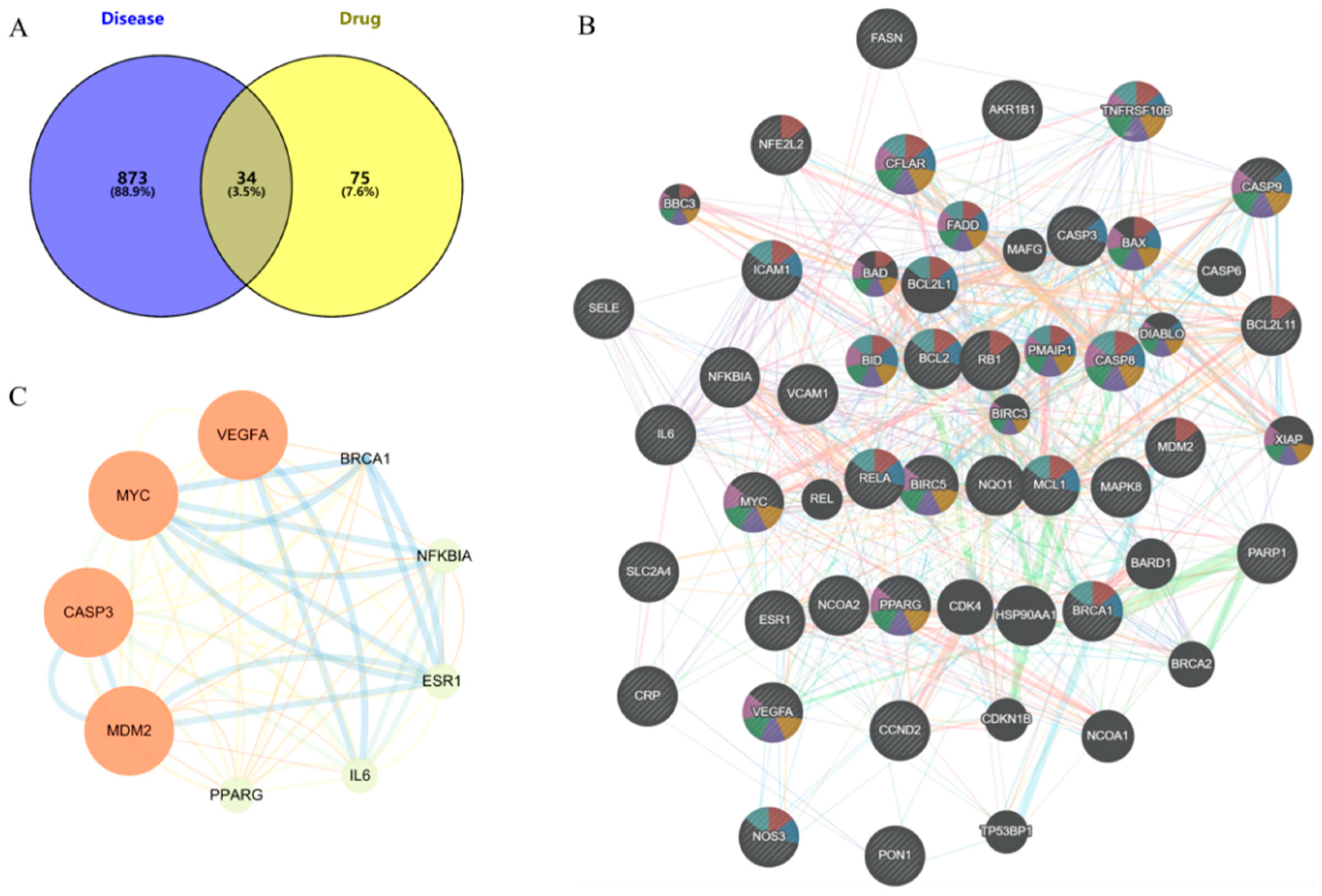
2.5. Network Pharmacology Analysis
2.5.1. Predicted Component and Disease Targets
2.5.2. PPI Network
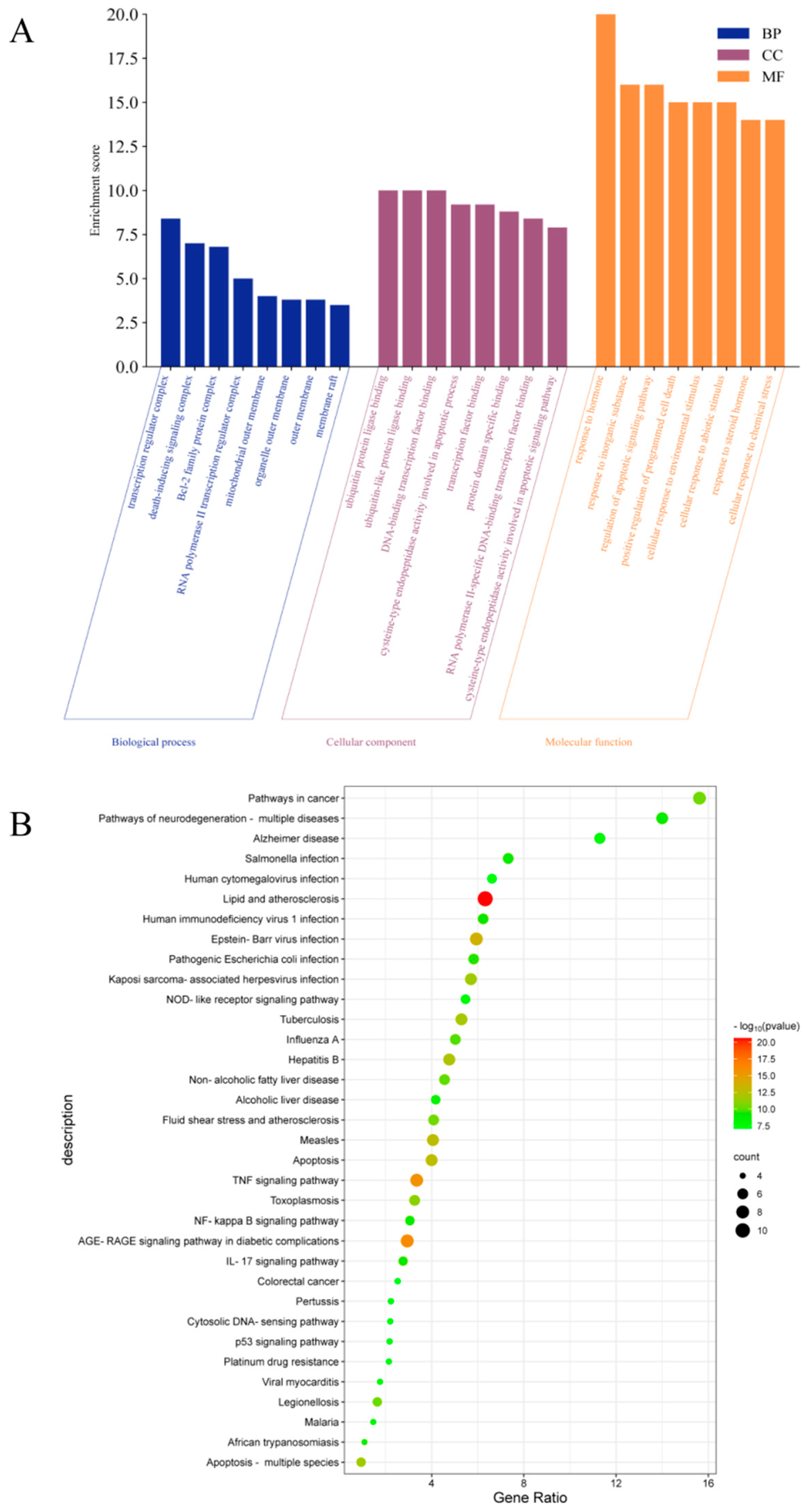
2.5.3. GO and KEGG Enrichment Analysis
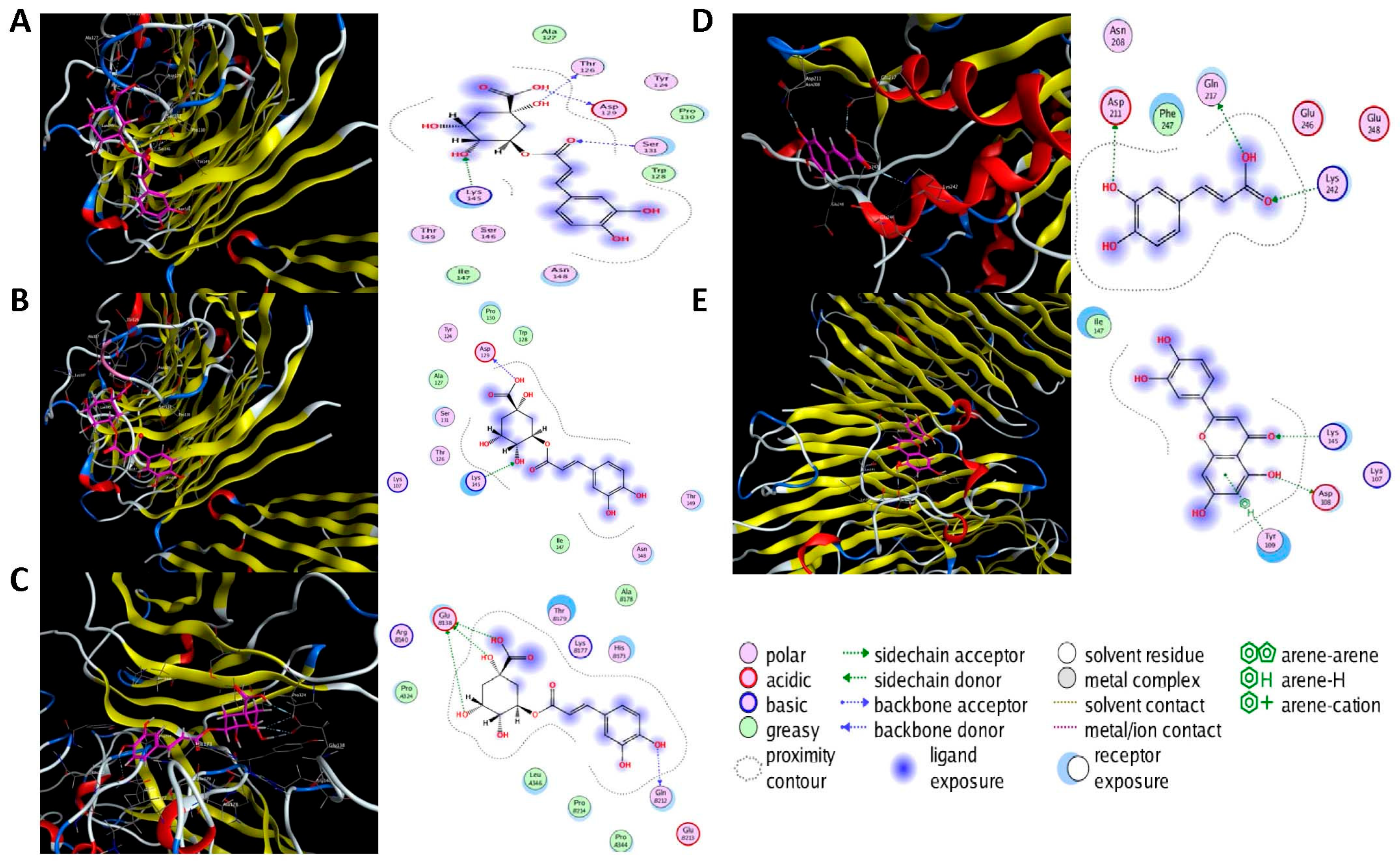
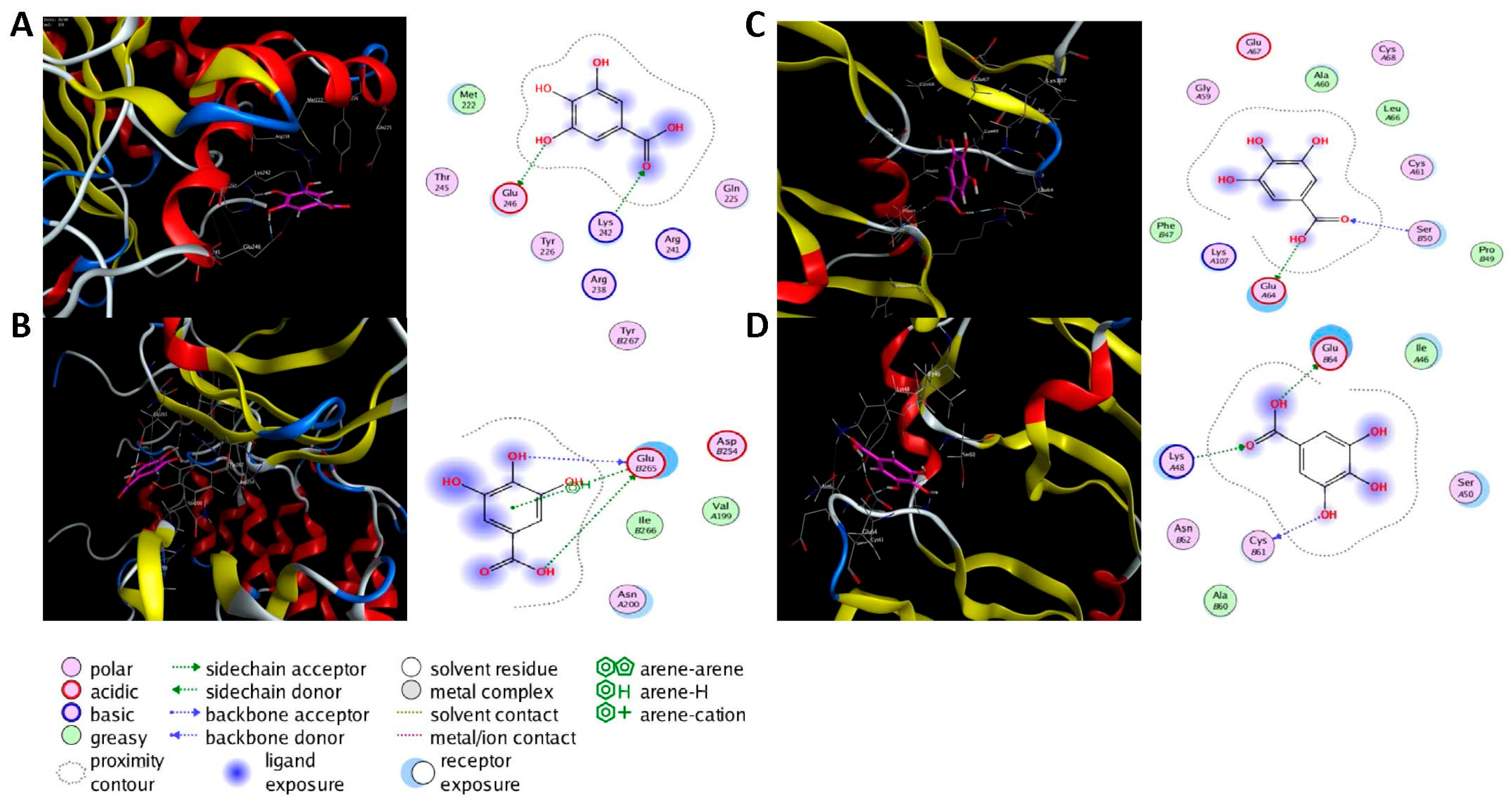
2.6. Molecular Docking Analysis
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Sample Preparation
4.3. HPLC Fingerprint Analysis
4.4. Antioxidant Assays
4.4.1. α-Glucosidase Assay
4.4.2. Xanthine Oxidase Assay
4.5. Effects of ZP on Hyperlipemia Zebrafish
4.5.1. Zebrafish Strains and Maintenance
4.5.2. Construction and Validation of Hyperuricemia Zebrafish
4.5.3. Determination of UA, Cr, BUN and XOD Levels in the Zebrafish
4.5.4. q-PCR Detection
4.6. Statistical Analysis
4.7. Network Pharmacology Analysis
4.7.1. Network Target Collection
4.7.2. Protein–Protein Interaction (PPI) Analysis
4.7.3. Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) Pathway Enrichment Analysis of Targets
4.8. Molecular Docking Method
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Calamelli, E.; Trozzo, A.; Di, B.E.; Serra, L.; Bottau, P. Hazelnut allergy. Medicina 2021, 57, 67. [Google Scholar] [CrossRef] [PubMed]
- Bottone, A.; Cerulli, A.; D’Urso, G.; Masullo, M.; Montoro, P.; Napolitano, A.; Piacente, S. Plant specialized metabolites in hazelnut (Corylus avellana) kernel and byproducts: An update on chemistry, biological activity, and analytical aspects. Planta Medica 2019, 85, 840–855. [Google Scholar] [CrossRef] [PubMed]
- Uchikura, T.; Kitano, T.; Yoshimura, M.; Amakura, Y. Characterization of phenolic constituents in hazelnut kernels. Biosci. Biotechnol. Biochem. 2023, 87, 688–695. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.; Li, J.; Sun, M.; Wang, X.; Zhao, J.; Zhang, W.; Lv, G.; Wang, Y.; Lin, Z. Effects of hazelnut soluble dietary fiber on lipid-lowering and gut microbiota in high-fat-diet-fed rats. Int. J. Biol. Macromol. 2023, 256 Pt 2, 128538. [Google Scholar] [CrossRef] [PubMed]
- Ramezan, M.; Hosseini, H.M.; Salimi, A.; Ramezan, Y. Study of the Apoptotic Impacts of Hazelnut Oil on the Colorectal Cancer Cell Line. Adv. Biomed. Res. 2023, 12, 76. [Google Scholar]
- Zhao, J.; Wang, X.; Lin, H.; Lin, Z. Hazelnut and its by-products: A comprehensive review of nutrition, phytochemical profile, extraction, bioactivities and applications. Food Chem. 2023, 413, 135576. [Google Scholar] [CrossRef]
- Allegrini, A.; Salvaneschi, P.; Schirone, B.; Cianfaglione, K.; Di, M.A. Multipurpose plant species and circular economy: Corylus avellana L. as a study case. Front. Biosci. 2022, 27, 11. [Google Scholar] [CrossRef]
- Zhao, J.; Wang, X.; Wang, Y.; Lv, G.; Lin, H.; Lin, Z. UPLC-MS/MS profiling, antioxidant andanti-inflammatory activities, and potentialhealth benefits prediction of phenoliccompounds in hazel leaf. Front. Nutr. 2023, 10, 1092071. [Google Scholar] [CrossRef]
- Tsao, R. Chemistry and biochemistry of dietary polyphenols. Nutrients 2010, 2, 1231–1246. [Google Scholar] [CrossRef]
- Mehmood, A.; Ishaq, M.; Zhao, L.; Safdar, B.; Rehman, A.U.; Munir, M.; Raza, A.; Nadeem, M.; Iqbal, W.; Wang, C. Natural compounds with xanthine oxidase inhibitory activity: A review. Chem. Biol. Drug Des. 2019, 93, 387–418. [Google Scholar] [CrossRef]
- Isaka, Y.; Takabatake, Y.; Takahashi, A.; Saitoh, T.; Yoshimori, T. Hyperuricemia-induced inflammasome and kidney diseases. Nephrol. Dial. Transplant. 2016, 31, 890–896. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Wen, X.; Kong, J. Recent progress on uric acid detection: A review. Crit. Rev. Anal. Chem. 2020, 50, 359–375. [Google Scholar] [CrossRef]
- Schmidt, H.M.; Kelley, E.E.; Straub, A.C. The impact of xanthine oxidase (XO) on hemolytic diseases. Redox Biol. 2019, 21, 101072. [Google Scholar] [CrossRef] [PubMed]
- Lyman, J.L. Blood urea nitrogen and creatinine. Emerg. Med. Clin. N. Am. 1986, 4, 223–233. [Google Scholar]
- Granados, J.C.; Ermakov, V.; Maity, K.; Vera, D.R.; Chang, G.; Nigam, S.K. The kidney drug transporter OAT1 regulates gut microbiome–dependent host metabolism. JCI Insight 2023, 8, e160437. [Google Scholar] [CrossRef] [PubMed]
- Sculley, D.G.; Dawson, P.A.; Emmerson, B.T.; Gordon, R.B. A review of the molecular basis of hypoxanthine-guanine phosphoribosyltransferase (HPRT) deficiency. Hum. Genet. 1992, 90, 195–207. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Yang, W.; Cao, W.; Yu, Z.; Zhang, G.; Long, L.; Guo, H.; Qu, H.; Fu, C.; Chen, K. Effects and mechanism of Kedaling tablets for atherosclerosis treatment based on network pharmacology, molecular docking and experimental study. J. Ethnopharmacol. 2024, 319, 117108. [Google Scholar] [CrossRef] [PubMed]
- Alberti, Á.; Riethmüller, E.; Béni, S.; Kéry, Á. Evaluation of radical scavenging activity of Sempervivum tectorum and Corylus avellana extracts with different phenolic composition. Nat. Prod. Commun. 2016, 11, 469–474. [Google Scholar]
- Alasalvar, C.; Karamać, M.; Amarowicz, R.; Shahidi, F. Antioxidant and antiradical activities in extracts of hazelnut kernel (Corylus avellana L.) and hazelnut green leafy cover. J. Agric. Food Chem. 2006, 54, 4826–4832. [Google Scholar] [CrossRef]
- Yuan, Y.; Zhai, Y.; Chen, J.; Xu, X.; Wang, H. Kaempferol ameliorates oxygen-glucose deprivation/reoxygenation-induced neuronal ferroptosis by activating Nrf2/SLC7A11/GPX4 axis. Biomolecules 2021, 11, 923. [Google Scholar] [CrossRef]
- Feng, Z.; Wang, C.; Yue, J.; Meng, Q.; Wu, J.; Sun, H. Kaempferol-induced GPER upregulation attenuates atherosclerosis via the PI3K/AKT/Nrf2 pathway. Pharm. Biol. 2021, 59, 1104–1114. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Zhao, S.; Song, H.; Xu, T.; Fang, Q.; Hu, G.; Sun, L. Kaempferol alleviates LD-mitochondrial damage by promoting autophagy: Implications in Parkinson’s disease. Redox Biol. 2021, 41, 101911. [Google Scholar] [CrossRef] [PubMed]
- Gao, W.; Wang, C.; Yu, L.; Sheng, T.; Wu, Z.; Wang, X.; Zhang, D.; Lin, Y.; Gong, Y. Chlorogenic acid attenuates dextran sodium sulfate-induced ulcerative colitis in mice through MAPK/ERK/JNK pathway. BioMed Res. Int. 2019, 2019, 6769789. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.L.; Li, X.Y.; Dou, H.Y.; Wang, X.D.; Li, J.D.; Shen, L.; Ji, H.F. Myricetin supplementation decreases hepatic lipid synthesis and inflammation by modulating gut microbiota. Cell Rep. 2021, 36, 109641. [Google Scholar] [CrossRef] [PubMed]
- Maiuolo, J.; Oppedisano, F.; Gratteri, S.; Muscoli, C.; Mollace, V. Regulation of uric acid metabolism and excretion. Int. J. Cardiol. 2016, 213, 8–14. [Google Scholar] [CrossRef]
- Torres, R.J.; Puente, S.; Menendez, A.; Fernandez-Garcia, N. Unapparent hypoxanthine-guanine phosphoribosyltransferase deficiency. Clin. Chim. Acta 2017, 472, 136–138. [Google Scholar] [CrossRef]
- Xiong, X.Y.; Liang, J.; Guo, S.Y.; Dai, M.Z.; Zhou, J.L.; Zhang, Y.; Liu, Y. A natural complex product Yaocha reduces uric acid level in a live zebrafish model. J. Pharmacol. Toxicol. Methods 2020, 102, 106681. [Google Scholar] [CrossRef]
- Su, H.Y.; Yang, C.; Liang, D.; Liu, H.F. Research advances in the mechanisms of hyperuricemia-induced renal injury. BioMed Res. Int. 2020, 2020, 5817348. [Google Scholar] [CrossRef]
- Godfrey, M.S.; Friedman, L.N. Tuberculosis and biologic therapies: Anti-tumor necrosis factor-α and beyond. Clin. Chest Med. 2019, 40, 721–739. [Google Scholar] [CrossRef]
- Zhou, Y.; Fang, L.; Jiang, L.; Wen, P.; Cao, H.; He, W.; Dai, C.; Yang, J. Uric acid induces renal inflammation via activating tubular NF-κB signaling pathway. PLoS ONE 2012, 7, e39738. [Google Scholar] [CrossRef]
- Liu, H.B.; Yang, M.; Li, W.; Luo, T.; Wu, Y.; Huang, X.Y.; Zhang, Y.L.; Liu, T.; Luo, Y. Dispelling Dampness, Relieving Turbidity and Dredging Collaterals Decoction, Attenuates Potassium Oxonate-Induced Hyperuricemia in Rat Models. Drug Des. Dev. Ther. 2023, 17, 2287–2301. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Cao, P.; Xiao, Z.; Ruan, Z. m(6)Amethyltransferase METTL3 relieves cognitive impairment of hyperuricemia mice via inactivating MyD88/NF-κB pathway mediated NLRP3-ASC-Caspase1 inflammasome. Int. Immunopharmacol. 2022, 113, 109375. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Gong, S.; Li, K.; Wu, G.; Zheng, X.; Zheng, J.; Lu, X.; Zhang, L.; Li, J.; Su, Z.; et al. Coptisine protects against hyperuricemic nephropathy through alleviating inflammation, oxidative stress and mitochondrial apoptosis via PI3K/Akt signaling pathway. Biomed. Pharmacother. 2022, 156, 113941. [Google Scholar] [CrossRef] [PubMed]
- Amatjan, M.; Li, N.; He, P.; Zhang, B.; Mai, X.; Jiang, Q.; Xie, H.; Shao, X. A novel approach based on gut microbiota analysis and network pharmacology to explain the mechanisms of action of Cichorium intybus L. formula in the improvement of hyperuricemic nephropathy in rats. Drug Des. Dev. Ther. 2023, 17, 107–128. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, F.J.; Shah, Y.M. PPARalpha: Mechanism of species differences and hepatocarcinogenesis of peroxisome proliferators. Toxicology 2008, 246, 2–8. [Google Scholar] [CrossRef] [PubMed]
- Poynter, M.E.; Daynes, R.A. Peroxisome proliferator-activated receptor α activation modulates cellular redox status, represses nuclear factor-κB signaling, and reduces inflammatory cytokine production in aging. J. Biol. Chem. 1998, 273, 32833–32841. [Google Scholar] [CrossRef]
- Liu, L.; Zhang, C.; Qu, S.; Liu, R.; Chen, H.; Liang, Z.; Tian, Z.; Li, L.; Ma, S.; Liu, X. ESR1 inhibits ionizing radiation-induced ferroptosis in breast cancer cells via the NEDD4L/CD71 pathway. Arch. Biochem. Biophys. 2022, 725, 109299. [Google Scholar] [CrossRef]
- Wang, W.; Liou, T.H.; Lee, W.J.; Hsu, C.T.; Lee, M.F.; Chen, H.H. ESR1 gene and insulin resistance remission are associated with serum uric acid decline for severely obese patients undergoing bariatric surgery. Surg. Obes. Relat. Dis. 2014, 10, 14–22. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | Retention Time/min | Peak Area of Sample | Peak Area of Standard | Concentration of Standard/μg/mL | Quantity Contained/mg/g |
|---|---|---|---|---|---|
| Kaempferol | 33.688 | 174,535 | 13,444 | 380 | 221.995 |
| Chlorogenic acid | 12.067 | 23,233 | 25,414 | 200 | 8.228 |
| Myricetin | 39.546 | 131,239 | 89,580 | 60 | 3.956 |
| Ellagic acid | 37.246 | 10,252 | 32,741 | 40 | 0.564 |
| Luteolin | 49.856 | 7840 | 68,931 | 50 | 0.256 |
| Resveratrol | 35.988 | 1998 | 83,851 | 200 | 0.214 |
| Caffeic acid | 16.262 | 4607 | 37,340 | 20 | 0.111 |
| Gallic acid | 3.063 | 52,577 | 922,439 | 40 | 0.103 |
| p-Coumaric acid | 25.118 | 5200 | 466,775 | 35.8 | 0.018 |
| Target | PDB ID | Target Structure | Compound | Interaction | Affinity (kcal/mol) | Receptor | Distance |
|---|---|---|---|---|---|---|---|
| BRCA1 | 1LOB |  | Chlorogenic acid | H-acceptor | −7.6 | LYS C145 | 2.98 |
| −7.5 | LYS C145 | 2.99 | |||||
| Luteolin | H-acceptor | −8.3 | LYS G145 | 3.10 | |||
| CASP3 | 7RN7 |  | Caffeic acid | H-acceptor | −7.5 | LYS B242 | 2.93 |
| Gallic acid | H-acceptor | −9.0 | LYS B242 | 3.18 | |||
| NFKBIA | 1IKN |  | Chlorogenic acid | H-donor | −7.4 | GLU D138 | −7.4 |
| Gallic acid | H-donor | −9.4 | GLU C265 | 2.80 | |||
| VEGFA | 1MJV |  | Gallic acid | H-donor | −7.3 | GLU A64 | 2.80 |
| H-acceptor | −10.2 | LYS A48 | 2.94 |
| Time (min) | 0.1% Phosphoric Acid Aqueous Solution (A) | Methanol (B) | Flow Rate (mL/min) |
|---|---|---|---|
| 0 | 80% | 20% | 1.0 |
| 8 | 75% | 25% | 1.0 |
| 15 | 70% | 30% | 1.0 |
| 30 | 60% | 40% | 1.0 |
| 50 | 55% | 45% | 1.0 |
| 55 | 40% | 60% | 1.0 |
| 60 | 80% | 20% | 1.0 |
| Gene Name | Sequence (5′-3′) | Size (bp) |
|---|---|---|
| β-actin | TCG AGC AGG AGA TGG GAA CC | 26 |
| β-actin | CTC GTG GAT ACC GCA AGA TTC | 27 |
| OAT1 | GGA CCT GTA AGG CCA GAT CC | 26 |
| OAT1 | TTG CAG TAG CTT GTC GGT GT | 26 |
| HPRT | TTG CAG TAG CTT GTC GGT GT | 26 |
| HPRT | CAG ACG TTC AGT TCG GTC CA | 26 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Zhao, J.; Lin, Z.; Li, J.; Li, X.; Xu, X.; Wang, Y.; Lv, G.; Lin, H.; Lin, Z. Analysis of Polyphenol Extract from Hazel Leaf and Ameliorative Efficacy and Mechanism against Hyperuricemia Zebrafish Model via Network Pharmacology and Molecular Docking. Molecules 2024, 29, 317. https://doi.org/10.3390/molecules29020317
Wang X, Zhao J, Lin Z, Li J, Li X, Xu X, Wang Y, Lv G, Lin H, Lin Z. Analysis of Polyphenol Extract from Hazel Leaf and Ameliorative Efficacy and Mechanism against Hyperuricemia Zebrafish Model via Network Pharmacology and Molecular Docking. Molecules. 2024; 29(2):317. https://doi.org/10.3390/molecules29020317
Chicago/Turabian StyleWang, Xinhe, Jiarui Zhao, Zhi Lin, Jun Li, Xiaowen Li, Xinyi Xu, Yuchen Wang, Guangfu Lv, He Lin, and Zhe Lin. 2024. "Analysis of Polyphenol Extract from Hazel Leaf and Ameliorative Efficacy and Mechanism against Hyperuricemia Zebrafish Model via Network Pharmacology and Molecular Docking" Molecules 29, no. 2: 317. https://doi.org/10.3390/molecules29020317
APA StyleWang, X., Zhao, J., Lin, Z., Li, J., Li, X., Xu, X., Wang, Y., Lv, G., Lin, H., & Lin, Z. (2024). Analysis of Polyphenol Extract from Hazel Leaf and Ameliorative Efficacy and Mechanism against Hyperuricemia Zebrafish Model via Network Pharmacology and Molecular Docking. Molecules, 29(2), 317. https://doi.org/10.3390/molecules29020317