

Therapeutic Potential of Fungal Terpenes and Terpenoids: Application in Skin Diseases


Abstract
:1. Introduction
2. General Characteristics and Occurrence of Terpenes and Terpenoids
3. Biogenesis
4. Structure and Chemical Classification of Terpenes and Terpenoids
4.1. Haemiterpens
4.2. Monoterpenes and Monoterpenoids
4.3. Sesquiterpenes and Sesquiterpenoids
4.4. Diterpenes and Diterpenoids
4.5. Sesterpenes and Sesterpenoids
4.6. Triterpenes and Triterpenoids
4.7. Tetraterpenes and Tetraterpenoids
4.8. Polyterpenoids
5. Pharmacological Activities in Skin Disorders
5.1. Anti-Inflammatory Activity
5.2. Antimicrobial Activity
5.3. Anti-Cancer Effects
5.4. Inhibition of Tyrosinase
5.5. Photoprotective Properties
6. Methods
7. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Chen, H.P.; Liu, J.K. Secondary metabolites from higher fungi. In Progress in the Chemistry of Organic Natural Products; Springer: Cham, Switzerland, 2017; Volume 106, pp. 1–201. [Google Scholar]
- Wang, S.; Bao, L.; Zhao, F.; Wang, Q.; Li, S.; Ren, J.; Li, L.; Wen, H.; Guo, L.; Liu, H. Isolation, identification, and bioactivity of monoterpenoids and sesquiterpenoids from the mycelia of edible mushroom Pleurotus cornucopiae. J. Agric. Food Chem. 2013, 61, 5122–5129. [Google Scholar] [CrossRef]
- Ivanowa, T.S.; Krupodorova, T.A.; Barshteyn, V.Y.; Artamonova, A.B.; Shlyakhovenko, V.A. Anticancer substances of mushroom origin. Exp. Oncol. 2014, 36, 58–66. [Google Scholar]
- Gallo, R.L. Human skin is the largest epithelial surface for interaction with microbes. J. Investig. Dermatol. 2017, 137, 1213–1214. [Google Scholar] [CrossRef]
- Yuan, H.; Ma, Q.; Ye, L.; Piao, G. The traditional medicine and modern medicine from natural products. Molecules 2016, 21, 559. [Google Scholar] [CrossRef] [PubMed]
- Fraga, B.M. Natural sesquiterpenoids. Nat. Prod. Rep. 2011, 28, 1580–1610. [Google Scholar] [CrossRef]
- Hu, J.Y.; Yang, T.; Liu, J.; Xiao, L.; Lin, L.B.; Li, Y.C.; Ge, M.Y.; Ji, P.; Xiao, J.; Wang, X.L. Two new sesquiterpenoids from plant endophytic fungus Flammulina velutipes. J. Asian Nat. Prod. Res. 2023, 25, 156–162. [Google Scholar] [CrossRef] [PubMed]
- Su, H.G.; Peng, X.R.; Shi, Q.Q.; Huang, Y.J.; Zhou, L.; Qiu, M.H. Lanostane triterpenoids with anti-inflammatory activities from Ganoderma lucidum. Phytochemistry 2020, 173, 112256. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Gao, Q.; Rong, C.; Wang, S.; Zhao, Z.; Liu, Y.; Xu, J. Immunomodulatory effects of edible and medicinal mushrooms and their bioactive immunoregulatory products. J. Fungi 2020, 6, 269. [Google Scholar] [CrossRef] [PubMed]
- Erbiai, E.H.; Pinto da Silva, L.; Saidi, R.; Lamrani, Z.; Esteves da Silva, J.C.G.; Maouni, A. Chemical composition, bioactive compounds, and antioxidant activity of two wild edible mushrooms Armillaria mellea and Macrolepiota procera from two countries (Morocco and Portugal). Biomolecules 2021, 11, 575. [Google Scholar] [CrossRef]
- Park, J.; Nguyen, T.M.N.; Park, H.A.; Nguyen, M.T.T.; Lee, N.Y.; Ban, S.Y.; Park, K.B.; Lee, C.K.; Kim, J.; Park, J.T. Protective effects of lanostane triterpenoids from Chaga mushroom in human keratinocytes, HaCaT cells, against inflammatory and oxidative stresses. Int. J. Mol. Sci. 2023, 24, 12803. [Google Scholar] [CrossRef]
- Harhaji, L.; Mijatović, S.; Maksimović-Ivanić, D.; Stojanović, I.; Momčilović, M.; Maksimović, V.; Tufegdžić, S.; Marjanović, Ž.; Mostarica-Stojković, M.; Vučinić, Ž.; et al. Anti-tumor effect of Coriolus versicolor methanol extract against mouse B16 melanoma cells: In vitro and in vivo study. Food Chem. Toxicol. 2008, 46, 1825–1833. [Google Scholar] [CrossRef]
- Paduch, R.; Kandefer-Szerszeń, M.; Trytek, M.; Fiedurek, J. Terpenes: Substances useful in human healthcare. Arch. Immunol. Ther. Exp. 2007, 55, 315–327. [Google Scholar] [CrossRef]
- Jaeger, R.; Cuny, E. Terpenoids with special pharmacological significance: A review. Nat. Prod. Commun. 2016, 11, 1373–1390. [Google Scholar] [CrossRef] [PubMed]
- Breitmaier, E. Terpenes: Flavors, Fragrances, Pharmaca, Pheromones; Wiley-VCH Verlag GmbH & Co KGaA: Weinheim, Germany, 2006; p. 214. ISBN 9783527317868. [Google Scholar]
- Cox-Georgian, D.; Ramadoss, N.; Dona, C.; Basu, C. Therapeutic and medicinal uses of terpenes. In Medicinal Plants; Springer: Cham, Switzerland, 2019; pp. 333–359. [Google Scholar]
- Tetali, S.D. Terpenes and isoprenoids: A wealth of compounds for global use. Planta 2019, 249, 1–8. [Google Scholar] [CrossRef]
- Zhang, L.; Yin, M.; Feng, X.; Ibrahim, S.A.; Liu, Y.; Huang, W. Anti-inflammatory activity of four triterpenoids isolated from Poriae cutis. Foods 2021, 10, 3155. [Google Scholar] [CrossRef] [PubMed]
- Bunbamrung, N.; Intaraudom, C.; Dramae, A.; Boonyuen, N.; Veeranondha, S.; Rachtawee, P.; Pittayakhajonwut, P. Antimicrobial activity of illudalane and alliacane sesquiterpenes from the mushroom Gloeostereum incarnatum BCC41461. Phytochem. Lett. 2017, 20, 274–281. [Google Scholar] [CrossRef]
- Duan, Y.C.; Feng, J.; Bai, N.; Li, G.H.; Zhang, K.Q.; Zhao, P.J. Four novel antibacterial sesquiterpene-α-amino acid quaternary ammonium hybrids from the mycelium of mushroom Stereum hirsutum. Fitoterapia 2018, 128, 213–217. [Google Scholar] [CrossRef]
- Kohlmünzer, S. Farmakognozja; PZWL: Warsaw, Poland, 1998; pp. 277–380. ISBN 83-200-2230-4. [Google Scholar]
- Goodwin, T.W. The biogenesis of terpenes and steroids. In Rodd’s Chemistry of Carbon Compounds, 2nd ed.; Elsevier: Amsterdam, The Netherlands, 1964; Volume II, pp. 54–137. [Google Scholar]
- Wang, Q.; Cao, R.; Zhang, Y.; Qi, P.; Wang, L.; Fang, S. Biosynthesis and regulation of terpenoids from Basidiomycetes: Exploration of new research. AMB Express 2021, 11, 150. [Google Scholar] [CrossRef] [PubMed]
- Ruzicka, L. The isoprene rule and the biogenesis of terpenic compounds. Experientia 1953, 9, 357–367. [Google Scholar] [CrossRef]
- Peng, H.; Shahidi, F. Bioactive compounds and bioactive properties of Chaga (Inonotus obliquus) mushroom: A review. J. Food Bioact. 2020, 12, 9–75. [Google Scholar] [CrossRef]
- Perveen, S. Terpenes and Terpenoids; IntechOpen: London, UK, 2018; pp. 1–152. ISBN 978-1-78984-777-2. [Google Scholar]
- Mabou, D.F.; Belinda, I.; Yossa, N. Terpenes: Structural classification and biological activities. IOSR–J. Pharm. Biol. Sci. 2021, 16, 2319–7676. [Google Scholar]
- Kabir, A.; Cacciagrano, F.; Tartaglia, A.; Lipsi, M.; Ulusoy, H.I.; Locatelli, M. Analysis of monoterpenes and monoterpenoids. In Recent Advances in Natural Products Analysis; Elsevier: Amsterdam, The Netherlands, 2020; pp. 274–286. [Google Scholar]
- Pennerman, K.K.; Yin, G.; Bennett, J.W. Health effects of small volatile compounds from east asian medicinal mushrooms. Mycobiology 2015, 43, 9–13. [Google Scholar] [CrossRef]
- Guo, H.; Diao, Q.P.; Zhang, B.; Feng, T. Two new illudane sesquiterpenoids and one new menthane monoterpene from cultures of Craterellus cornucopioides. J. Asian Nat. Prod. Res. 2019, 21, 123–128. [Google Scholar] [CrossRef]
- Xu, Y.C.; Xie, X.X.; Zhou, Z.Y.; Feng, T.; Liu, J.K. A new monoterpene from the poisonous mushroom Trogia venenata, which has caused sudden unexpected death in Yunnan province, China. Nat. Prod. Res. 2018, 32, 2547–2552. [Google Scholar] [CrossRef]
- Negi, K.; Singh, S.; Gahlot, M.S.; Tyagi, S.; Gupta, A. Terpenoids from medicinal plants beneficial for human health care: Review. Int. J. Botany Stud. 2020, 4, 135–138. [Google Scholar]
- Duru, M.E.; Tel-Çayan, G. Biologically active terpenoids from mushroom origin: A review. Rec. Nat. Prod. 2015, 9, 456–483. [Google Scholar]
- Tao, Q.; Ma, K.; Yang, Y.; Wang, K.; Chen, B.; Huang, Y.; Han, J.; Bao, L.; Liu, X.B.; Yang, Z.; et al. Bioactive sesquiterpenes from the edible mushroom Flammulina velutipes and their biosynthetic pathway confirmed by genome analysis and chemical evidence. J. Org. Chem. 2016, 81, 9867–9877. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Ryoo, R.; Choi, J.H.; Kim, J.H.; Kim, S.H.; Kim, K.H. Trichothecene and tremulane sesquiterpenes from a hallucinogenic mushroom Gymnopilus junonius and their cytotoxicity. Arch. Pharm. Res. 2020, 43, 214–223. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.R.; Lee, D.; Lee, B.S.; Ryoo, R.; Pang, C.; Kang, K.S.; Kim, K.H. Phallac acids A and B, new sesquiterpenes from the fruiting bodies of Phallus luteus. J. Antibiot. 2020, 73, 729–732. [Google Scholar] [CrossRef] [PubMed]
- Eksi, G.; Kurbanoglu, S.; Erdem, S.A. Analysis of diterpenes and diterpenoids. In Recent Advances in Natural Products Analysis; Elsevier: Amsterdam, The Netherlands, 2020; pp. 313–345. [Google Scholar]
- Farhat, F.; Tariq, A.; Zikrea, A.; Nasim Fatima, R. Diterpenes from different fungal sources and their 13C-NMR data. In Terpenes and Terpenoids; IntechOpen: London, UK, 2018; Volume 6, pp. 111–141. [Google Scholar]
- Evidente, A.; Kornienko, A.; Lefranc, F.; Cimmino, A.; Dasari, R.; Evidente, M.; Mathieu, V.; Kiss, R. Sesterterpenoids with anticancer activity. Curr. Med. Chem. 2015, 22, 3502–3522. [Google Scholar] [CrossRef] [PubMed]
- Annang, F.; Pérez-Victoria, I.; Appiah, T.; Pérez-Moreno, G.; Domingo, E.; Martín, J.; Mackenzie, T.; Ruiz-Pérez, L.; González-Pacanowska, D.; Genilloud, O.; et al. Antiprotozoan sesterterpenes and triterpenes isolated from two ghanaian mushrooms. Fitoterapia 2018, 127, 341–348. [Google Scholar] [CrossRef]
- Yazdani, M.; Barta, A.; Berkecz, R.; Agbadua, O.G.; Ványolós, A.; Hohmann, J. Pholiols E-K, Lanostane-type triterpenes from Pholiota populnea with anti-inflammatory properties. Phytochemistry 2023, 205, 113480. [Google Scholar] [CrossRef]
- Chen, H.P.; Zhao, Z.Z.; Li, Z.H.; Huang, Y.; Zhang, S.B.; Tang, Y.; Yao, J.N.; Chen, L.; Isaka, M.; Feng, T.; et al. Anti-proliferative and anti-inflammatory lanostane triterpenoids from the polish edible mushroom Macrolepiota procera. J. Agric. Food Chem. 2018, 66, 3146–3154. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Chen, B.; Liang, J.; Han, J.; Zhou, L.; Zhao, R.; Liu, H.; Dai, H. Lanostane triterpenoids with PTP1B inhibitory and glucose-uptake stimulatory activities from mushroom Fomitopsis pinicola collected in North America. J. Agric. Food Chem. 2020, 68, 10036–10049. [Google Scholar] [CrossRef] [PubMed]
- He, P.; Zhang, Y.; Li, N. The phytochemistry and pharmacology of medicinal fungi of the genus Phellinus: A review. Food Funct. 2021, 12, 1856–1881. [Google Scholar] [CrossRef] [PubMed]
- Feng, T.; Gan, X.Q.; Zhao, Y.L.; Zhang, S.B.; Chen, H.P.; He, J.; Zheng, Y.S.; Sun, H.; Huang, R.; Li, Z.H.; et al. Tricholopardins A and B, anti-inflammatory terpenoids from the fruiting bodies of Tricholoma pardinum. J. Nat. Prod. 2019, 82, 45–50. [Google Scholar] [CrossRef]
- Jin, M.; Zhou, W.; Jin, C.; Jiang, Z.; Diao, S.; Jin, Z.; Li, G. Anti-inflammatory activities of the chemical constituents isolated from Trametes versicolor. Nat. Prod. Res. 2019, 33, 2422–2425. [Google Scholar] [CrossRef]
- Hu, Z.; Du, R.; Xiu, L.; Bian, Z.; Ma, C.; Sato, N.; Hattori, M.; Zhang, H.; Liang, Y.; Yu, S.; et al. Protective effect of triterpenes of Ganoderma lucidum on lipopolysaccharide-induced inflammatory responses and acute liver injury. Cytokine 2020, 127, 154917. [Google Scholar] [CrossRef]
- Bao, T.R.G.; Long, G.Q.; Wang, Y.; Wang, Q.; Liu, X.L.; Hu, G.S.; Gao, X.X.; Wang, A.H.; Jia, J.M. New lanostane-type triterpenes with anti-inflammatory activity from the epidermis of Wolfiporia cocos. J. Agric. Food Chem. 2022, 70, 4418–4433. [Google Scholar] [CrossRef]
- Kou, R.W.; Xia, B.; Wang, Z.J.; Li, J.N.; Yang, J.R.; Gao, Y.Q.; Yin, X.; Gao, J.M. Triterpenoids and meroterpenoids from the edible Ganoderma resinaceum and their potential anti-inflammatory, antioxidant and anti-apoptosis activities. Bioorg. Chem. 2022, 121, 105689. [Google Scholar] [CrossRef]
- Rohr, M.; Oleinikov, K.; Jung, M.; Sandjo, L.P.; Opatz, T.; Erkel, G. Anti-inflammatory tetraquinane diterpenoids from a Crinipellis species. Bioorg. Med. Chem. 2017, 25, 514–522. [Google Scholar] [CrossRef]
- Lagunes-Castro, M.S.; Aguila, S.; Herrera-Covarrubias, D.; Hernández-Aguilar, M.E.; Trigos, Á.; Vidal-Limon, A.; Suárez-Medellín, J. Structure-based virtual screening of sterols and triterpenoids isolated from Ganoderma (Agaricomycetes) medicinal mushrooms shows differences in their affinity for human glucocorticoid and mineralocorticoid receptors. Int. J. Med. Mushrooms 2021, 23, 1–13. [Google Scholar] [CrossRef]
- Taofiq, O.; Rodrigues, F.; Barros, L.; Barreiro, M.F.; Ferreira, I.C.F.R.; Oliveira, M.B.P.P. Mushroom ethanolic extracts as cosmeceuticals ingredients: Safety and ex vivo skin permeation studies. Food Chem. Toxicol. 2019, 127, 228–236. [Google Scholar] [CrossRef]
- Imokawa, G.; Ishida, K. Inhibitors of intracellular signaling pathways that lead to stimulated epidermal pigmentation: Perspective of anti-pigmenting agents. Int. J. Mol. Sci. 2014, 15, 8293–8315. [Google Scholar] [CrossRef]
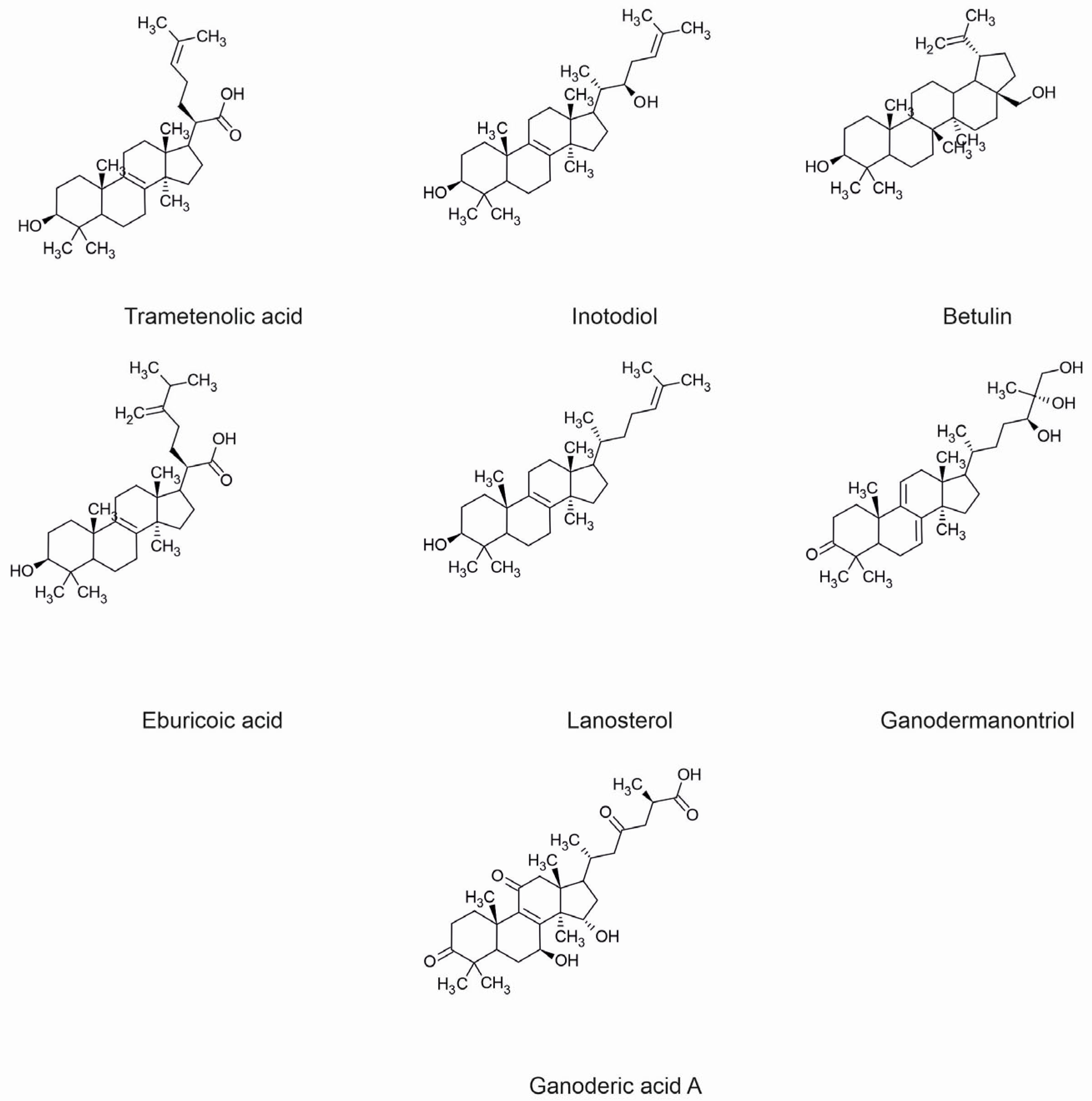
- Kim, J.W.; Kim, H.I.; Kim, J.H.; Kwon, O.C.; Son, E.S.; Lee, C.S.; Park, Y.J. Effects of ganodermanondiol, a new melanogenesis inhibitor from the medicinal mushroom Ganoderma lucidum. Int. J. Mol. Sci. 2016, 17, 1798. [Google Scholar] [CrossRef] [PubMed]
- Taofiq, O.; Martins, A.; Barreiro, M.F.; Ferreira, I.C.F.R. Anti-inflammatory potential of mushroom extracts and isolated metabolites. Trends Food Sci. Technol. 2016, 50, 193–210. [Google Scholar] [CrossRef]
- Deng, J.S.; Huang, S.S.; Lin, T.H.; Lee, M.M.; Kuo, C.C.; Sung, P.J.; Hou, W.C.; Huang, G.J.; Kuo, Y.H. Analgesic and anti-inflammatory bioactivities of eburicoic acid and dehydroeburicoic acid isolated from Antrodia camphorata on the inflammatory mediator expression in mice. J. Agric. Food Chem. 2013, 61, 5064–5071. [Google Scholar] [CrossRef]
- Kuo, Y.C.; Lai, C.S.; Tsai, C.Y.; Nagabhushanam, K.; Ho, C.T.; Pan, M.H. Inotilone suppresses phorbol ester-induced inflammation and tumor promotion in mouse skin. Mol. Nutr. Food Res. 2012, 56, 1324–1332. [Google Scholar] [CrossRef]
- Wisetsai, A.; Lekphrom, R.; Bua-Art, S.; Suebrasri, T.; Boonlue, S.; Tontapha, S.; Amornkitbamrung, V.; Senawong, T.; Schevenels, F.T. Scalarane sesterterpenoids with antibacterial and anti-proliferative activities from the mushroom Neonothopanus Nambi. Molecules 2021, 26, 7667. [Google Scholar] [CrossRef] [PubMed]
- Rahi, D.K.; Malik, D. Diversity of mushrooms and their metabolites of nutraceutical and therapeutic significance. J. Mycol. 2016, 2016, 7654123. [Google Scholar] [CrossRef]
- Liu, Y.P.; Dai, Q.; Wang, W.X.; He, J.; Li, Z.H.; Feng, T.; Liu, J.K. Psathyrins: Antibacterial diterpenoids from Psathyrella candolleana. J. Nat. Prod. 2020, 83, 1725–1729. [Google Scholar] [CrossRef]
- Stöckli, M.; Morinaka, B.I.; Lackner, G.; Kombrink, A.; Sieber, R.; Margot, C.; Stanley, C.E.; de Mello, A.J.; Piel, J.; Künzler, M. Bacteria-induced production of the antibacterial sesquiterpene lagopodin B in Coprinopsis cinerea. Mol. Microbiol. 2019, 112, 605–619. [Google Scholar] [CrossRef]
- Dasgupta, A.; Acharya, K. Mushrooms: An emerging resource for therapeutic terpenoids. 3 Biotech 2019, 9, 369. [Google Scholar] [CrossRef]
- Alves, M.J.; Froufe, H.J.C.; Costa, A.F.T.; Santos, A.F.; Oliveira, L.G.; Osório, S.R.M.; Abreu, R.M.V.; Pintado, M.; Ferreira, I.C.F.R. Docking studies in target proteins involved in antibacterial action mechanisms: Extending the knowledge on standard antibiotics to antimicrobial mushroom compounds. Molecules 2014, 19, 1672. [Google Scholar] [CrossRef] [PubMed]
- Kreuzenbeck, N.B.; Dhiman, S.; Roman, D.; Burkhardt, I.; Conlon, B.H.; Fricke, J.; Guo, H.; Blume, J.; Görls, H.; Poulsen, M.; et al. Isolation, (bio)synthetic studies and evaluation of antimicrobial properties of drimenol-type sesquiterpenes of Termitomyces fungi. Commun. Chem. 2023, 6, 79. [Google Scholar] [CrossRef] [PubMed]
- Cheng, T.; Chepkirui, C.; Decock, C.; Matasyoh, J.C.; Stadler, M. Sesquiterpenes from an Eastern African medicinal mushroom belonging to the genus Sanghuangporus. J. Nat. Prod. 2019, 82, 1283–1291. [Google Scholar] [CrossRef] [PubMed]
- Hassan, K.; Kemkuignou, B.M.; Stadler, M. Two new triterpenes from Basidiomata of the medicinal and edible mushroom, Laetiporus sulphureus. Molecules 2021, 26, 7090. [Google Scholar] [CrossRef]
- Richter, C.; Helaly, S.E.; Thongbai, B.; Hyde, K.D.; Stadler, M. Pyristriatins A and B: Pyridino-cyathane antibiotics from the Basidiomycete Cyathus cf. striatus. J. Nat. Prod. 2016, 79, 1684–1688. [Google Scholar] [CrossRef]
- Sandargo, B.; Michehl, M.; Stadler, M.; Surup, F. Antifungal sesquiterpenoids, rhodocoranes, from submerged cultures of the wrinkled peach mushroom, Rhodotus palmatus. J. Nat. Prod. 2020, 83, 720–724. [Google Scholar] [CrossRef]
- Bryant, J.M.; Bouchard, M.; Haque, A. Anticancer activity of ganoderic acid DM: Current status and future perspective. J. Clin. Cell. Immunol. 2017, 8, 535. [Google Scholar] [CrossRef]
- Ahmad, M.F. Ganoderma lucidum: A rational pharmacological approach to surmount cancer. J. Ethnopharmacol. 2020, 260, 113047. [Google Scholar] [CrossRef]
- Rodolfo, C.; Rocco, M.; Cattaneo, L.; Tartaglia, M.; Sassi, M.; Aducci, P.; Scaloni, A.; Camoni, L.; Marra, M. Ophiobolin a induces autophagy and activates the mitochondrial pathway of apoptosis in human melanoma cells. PLoS ONE 2016, 11, e0167672. [Google Scholar] [CrossRef]
- Liu, G.; Wang, K.; Kuang, S.; Cao, R.; Bao, L.; Liu, R.; Liu, H.; Sun, C. The natural compound GL22, isolated from Ganoderma mushrooms, suppresses tumor growth by altering lipid metabolism and triggering cell death. Cell Death Dis. 2018, 9, 689. [Google Scholar] [CrossRef]
- Wu, Z.; Li, Y. Grifolin exhibits anti-cancer activity by inhibiting the development and invasion of gastric tumor cells. Oncotarget 2017, 8, 21454. [Google Scholar] [CrossRef]
- Bouyahya, A.; El Allam, A.; Zeouk, I.; Taha, D.; Zengin, G.; Goh, B.H.; Catauro, M.; Montesano, D.; El Omari, N. Pharmacological Effects of Grifolin: Focusing on Anticancer Mechanisms. Molecules 2022, 27, 284. [Google Scholar] [CrossRef]
- Akihisa, T.; Nakamura, Y.; Tagata, M.; Tokuda, H.; Yasukawa, K.; Uchiyama, E.; Suzuki, T.; Kimura, Y. Anti-inflammatory and anti-tumor-promoting effects of triterpene acids and sterols from the fungus Ganoderma lucidum. Chem. Biodivers. 2007, 4, 224–231. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.M.; Zhang, X.J.; Liang, G.Y.; Yang, Y.F.; Zhong, J.J.; Xiao, J.H. Antitumor and antimetastatic activities of chloroform extract of medicinal mushroom Cordyceps taii in mouse models. BMC Complement. Altern. Med. 2015, 15, 216. [Google Scholar]
- Parvez, S.; Kang, M.; Chung, H.S.; Bae, H. Naturally occurring tyrosinase inhibitors: Mechanism and applications in skin health, cosmetics and agriculture industries. Phytother. Res. 2007, 21, 805–816. [Google Scholar] [CrossRef] [PubMed]
- Ren, Q.; Lu, X.Y.; Han, J.X.; Aisa, H.A.; Yuan, T. Triterpenoids and phenolics from the fruiting bodies of Inonotus hispidus and their activations of melanogenesis and tyrosinase. Chin. Chem. Lett. 2017, 28, 1052–1056. [Google Scholar] [CrossRef]
- Yan, Z.F.; Yang, Y.; Tian, F.H.; Mao, X.X.; Li, Y.; Li, C.T. Inhibitory and acceleratory effects of Inonotus obliquus on tyrosinase activity and melanin formation in B16 melanoma cells. Evid. Based. Complement. Alternat. Med. 2014, 2014, 259836. [Google Scholar] [CrossRef] [PubMed]
- Knežević, A.; Stajić, M.; Sofrenić, I.; Stanojković, T.; Milovanović, I.; Tešević, V.; Vukojević, J. Antioxidative, antifungal, cytotoxic and antineurodegenerative activity of selected Trametes species from Serbia. PLoS ONE 2018, 13, e0203064. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.K.; Cha, H.J. Poria cocos Wolf extracts repress pigmentation in vitro and in vivo Cell. Mol. Biol. 2018, 64, 80–84. [Google Scholar]
- Zhang, T.; Huang, S.; Qiu, J.; Wu, X.; Yuan, H.; Park, S. Beneficial effect of Gastrodia elata Blume and Poria cocos Wolf administration on acute uvb irradiation by alleviating inflammation through promoting the gut-skin axis. Int. J. Mol. Sci. 2022, 23, 10833. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Terpenes/Terpenoids | Species | Activity | Activity Description | Reference |
|---|---|---|---|---|
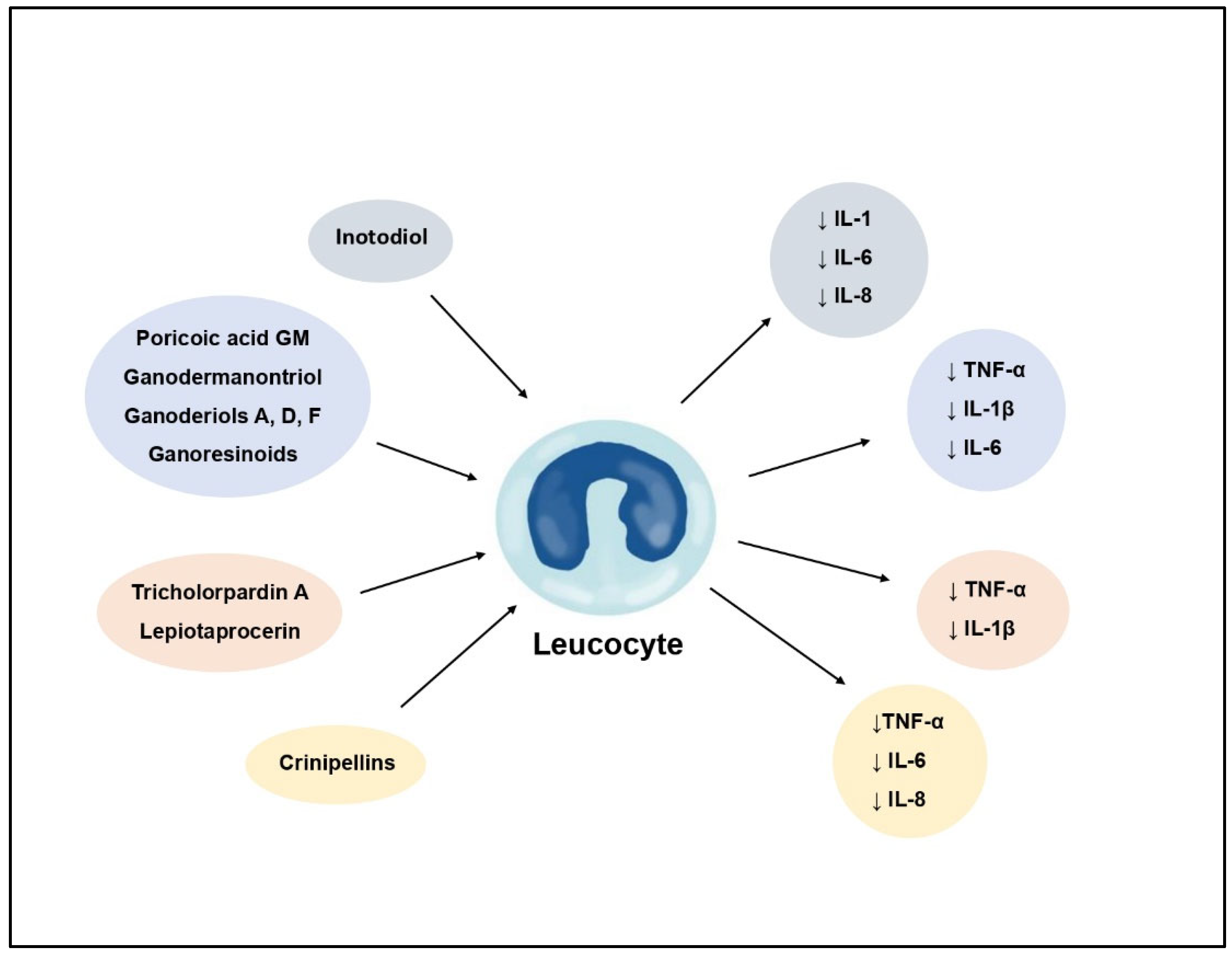
| Inotodiol | Inonotus obliquus | Anti- inflammatory | Decrease of IL-1, IL-6 and IL-8 levels | [11] |
| Tyrosinase activation | Increase of melanin level | [79] | ||
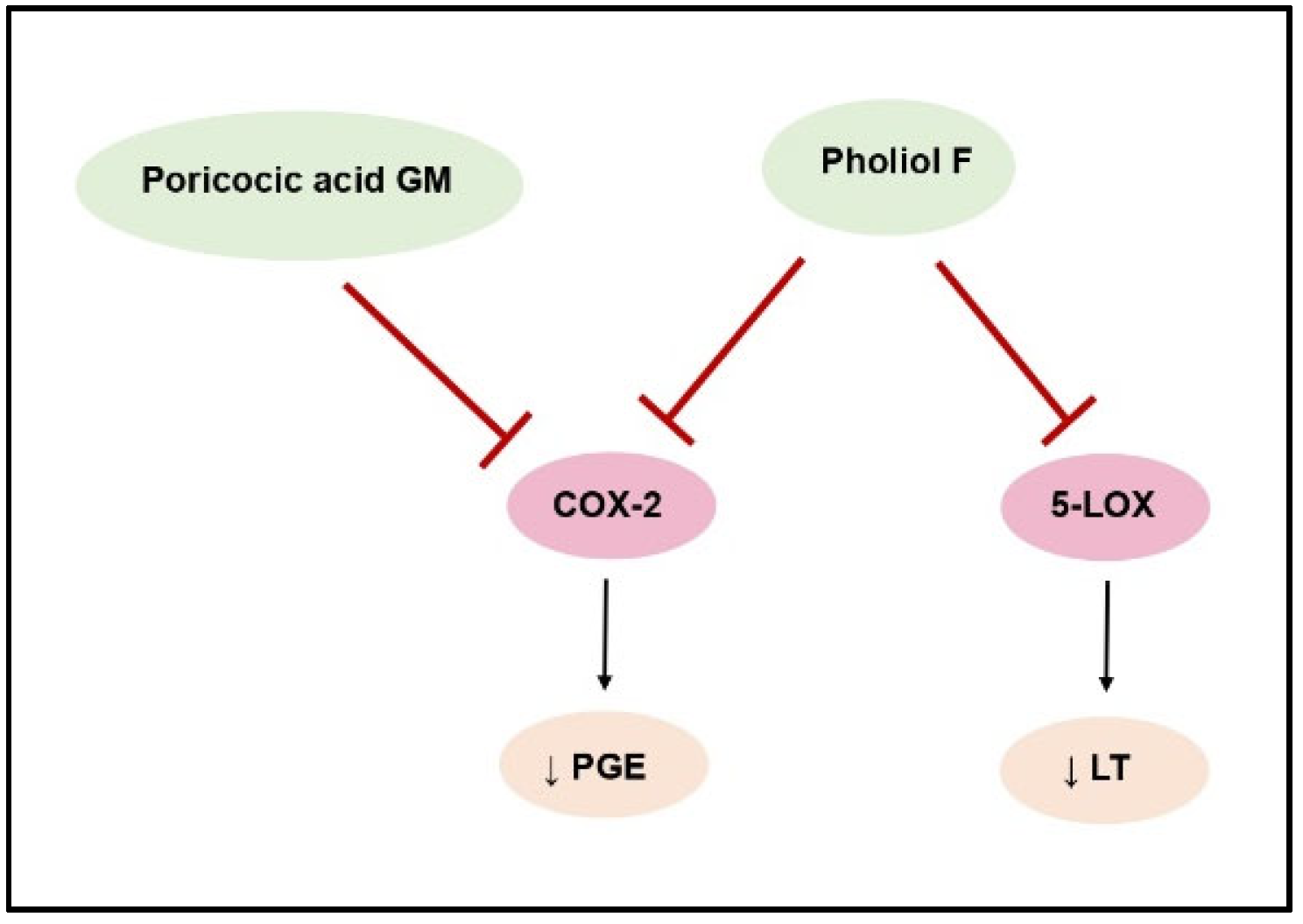
| Pholiol F | Pholiota populnea | Anti- inflammatory | COX-2 and 5-LOX inhibition—decrease of leukotrienes and prostaglandins secretion | [41] |
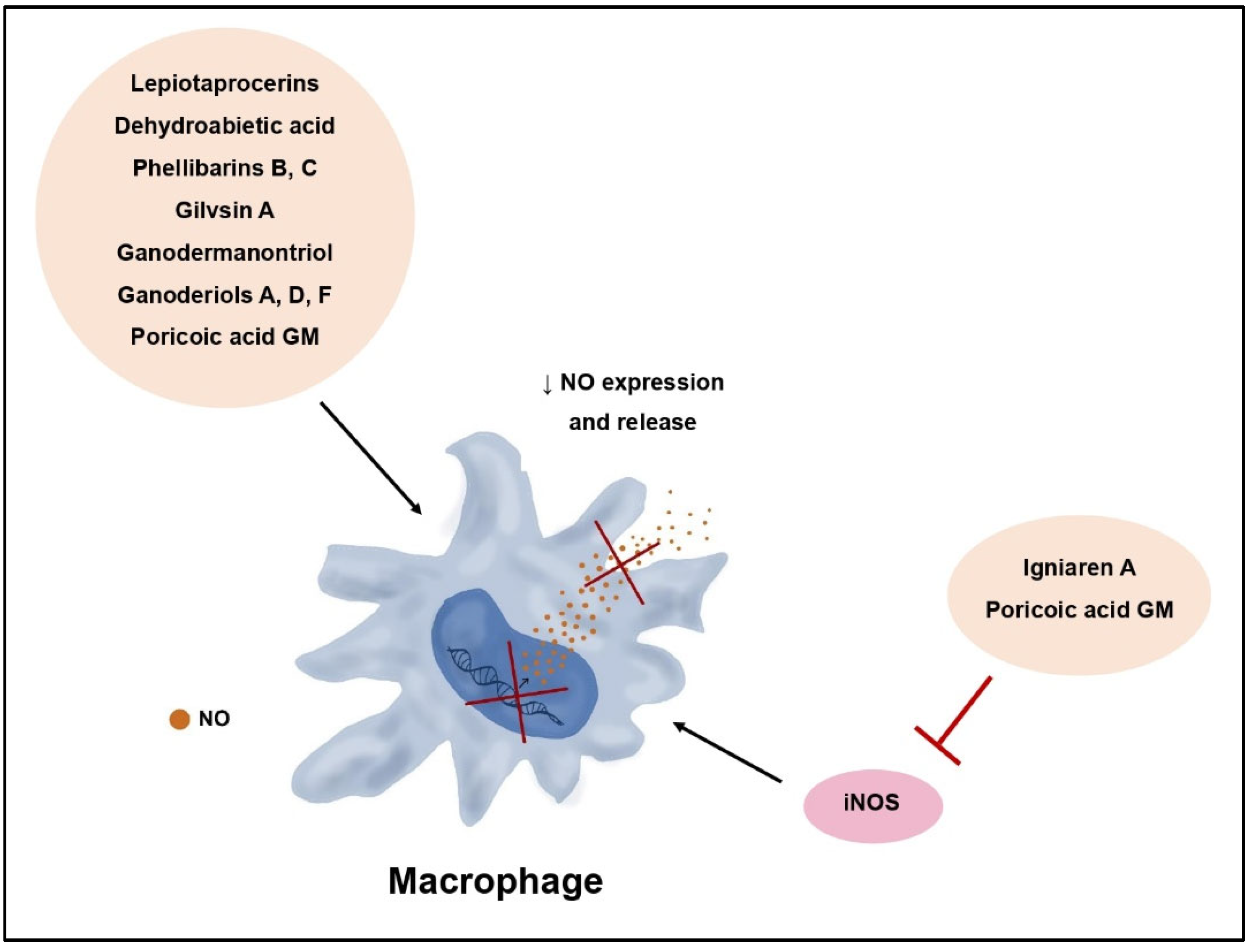
| Lepiotaprocerins | Macrolepiota procera | Anti- inflammatory | Decrease of NO synthesis, decrease of PGE2, TNF-α and IL-1β levels | [42] |
| Dehydroabietanoic acid | Phellinus pini | Anti- inflammatory | Inhibition of NO release | [44] |
| Phellibarins B, C | Phellinus rhabarbarinus | Anti- inflammatory | Inhibition of NO release | [44] |
| Gilvinin A | Phellinus rhabarbarinus | Anti- inflammatory | Inhibition of NO release | [44] |
| Igniarene B | Phellinus igniarius | Anti- inflammatory | iNOS inhibition | [44] |
| Tricholopardin A | Tricholoma pardinum | Anti- inflammatory | Inhibition of NO production, decrease of PGE2 TNF-α and IL-1β levels | [45] |
| Ganodermanontriol | Ganoderma lucidum | Anti- inflammatory | Decrease of NO, TNF-α, IL-1β, and IL-6 levels, inhibition of NF-κB and MAPK pathway, decrease of TLR4 and MyD88 expression | [47] |
| Anti-cancer | Anti-proliferative effect, apoptosis of cancer cells, inhibition of cell adhesion and migration | [70] | ||
| Ganoderiols A, D, F | Ganoderma lucidum | Anti- inflammatory | Decrease of NO, TNF-α, IL-1β, and IL-6 levels | [47] |
| Poricoic acid GM | Wolfiporia cocos | Anti- inflammatory | Decrease of NO, IL-1β, IL-6, TNF-α, and PGE2 levels, inhibition of NF-κB pathway, iNOS inhibition, decrease of COX-2 expression | [48] |
| Ganoresinoids | Ganoderma resinaceum | Anti- inflammatory | Decrease of IL-1β, IL-6, and TNF-α levels, inhibition of NF-κB and MAPK pathway, decrease of TLR4 expression | [49] |
| Crinipellins F, G | Crinipellis sp. | Anti- inflammatory | Decrease of CxCl1, CxCl11, CCL3, IL-6, IL-8, TNF-α, CCL2 and COX-2 synthesis, inhibition of NF-κB pathway | [50] |
| Psatyrins | Psathyrella candolleana | Antibacterial | Stapylococcus aureus | [60] |
| Lagopodin B | Coprinopsis cinerea | Antibacterial | Gram-positive bacteria | [61] |
| Stereumamides A, D | Stereum hirsutum | Antibacterial | Stapylococcus aureus | [20] |
| Stereostrein Q | Stereum hirsutum | Antibacterial | Stapylococcus aureus | [20] |
| (Z)-4-hydroxy-3-(3-hydroxy-3-methylbut-1-en-1-yl)benzoic acid | Montagnula donacina | Antibacterial | Stapylococcus aureus | [62] |
| Drimenol | Termitomyces sp. | Antifungal Antibacterial | Candida albicans, Penicilium notatum Stapylococcus aureus, Pseudomonas aeruginosa | [64] |
| Leptosporin C | Laetiporus sulphureus | Antifungal | Mucor hiemalis | [66] |
| Pyristriatins A, B | Cyathus cf. striatus | Antibacterial | Stapylococcus aureus | [67] |
| Rhodocoranes H, I | Rhodotus palmatus | Antibacterial | Stapylococcus aureus | [68] |
| Ganoderic acid DM | Ganoderma lucidum | Anti-cancer | Anti-proliferative and anti-metastatic effect, apoptosis and autophagy of cancer cells, increased recognition of cancer cells by CD 4+ T lymphocytes | [69] |
| Ganoderic acid A | Ganoderma lucidum | Anti-cancer | Anti-proliferative effect, apoptosis of cancer cells, inhibition of cell adhesion and migration | [70] |
| Ganoderic acid T | Ganoderma lucidum | Anti-cancer | Anti-proliferative effect, apoptosis of cancer cells, cytotoxicity | [70] |
| Lucidenic acids A, B, N | Ganoderma lucidum | Anti-cancer | Inhibition of cell migration | [70] |
| Ganoderiol F | Ganoderma lucidum | Anti-cancer | Cytotoxicity | [70] |
| Ophiobolin A | Bipolaris sp. | Anti-cancer | Mitochondrial dysfunction and apoptosis of cancer cells | [71] |
| GL22 | Ganoderma leucocontextum | Anti-cancer | FABP inhibition, mitochondrial dysfunction and apoptosis of cancer cells | [72] |
| Grifolin | Albatrellus confluens | Anti-cancer | Cytotoxicity and apoptosis of cancer cells | [73,74] |
| Trametenolic acid | Inonotus obliquus | Tyrosinase inhibition | Decrease of melanin level | [79] |
| Hispindic acids A, B | Inonotus hispidus | Tyrosine activation | Melanogenesis activation | [78] |
| Ganodermanondiol | Ganoderma lucidum | Tyrosinase inhibition | Decrease of melanin level | [53,54] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Trepa, M.; Sułkowska-Ziaja, K.; Kała, K.; Muszyńska, B. Therapeutic Potential of Fungal Terpenes and Terpenoids: Application in Skin Diseases. Molecules 2024, 29, 1183. https://doi.org/10.3390/molecules29051183
Trepa M, Sułkowska-Ziaja K, Kała K, Muszyńska B. Therapeutic Potential of Fungal Terpenes and Terpenoids: Application in Skin Diseases. Molecules. 2024; 29(5):1183. https://doi.org/10.3390/molecules29051183
Chicago/Turabian StyleTrepa, Monika, Katarzyna Sułkowska-Ziaja, Katarzyna Kała, and Bożena Muszyńska. 2024. "Therapeutic Potential of Fungal Terpenes and Terpenoids: Application in Skin Diseases" Molecules 29, no. 5: 1183. https://doi.org/10.3390/molecules29051183
APA StyleTrepa, M., Sułkowska-Ziaja, K., Kała, K., & Muszyńska, B. (2024). Therapeutic Potential of Fungal Terpenes and Terpenoids: Application in Skin Diseases. Molecules, 29(5), 1183. https://doi.org/10.3390/molecules29051183