
Potential of Thuja occidentalis L. Essential Oil and Water Extracts against Field Crop Pests

 ,
,  , ,
, ,
Abstract
:1. Introduction
- The proposed doses of EOTO and WETOs will increase the mortality of the pests mentioned;
- EOTO and WETOs will reduce the mass of food eaten by CPB and the surface area of leaves eaten by pea leaf weevils;
- Nymphs and wingless females of A. fabae, as well as males and females of CPB and S. lineatus, will differ in response to the EOTO and WETOs used;
- Concentrations effective against aphids will be safe for the non-target organism—H. axyridis larvae, i.e., they will not increase the mortality of lady beetles or affect their voracity.
2. Results
2.1. Black Bean Aphid
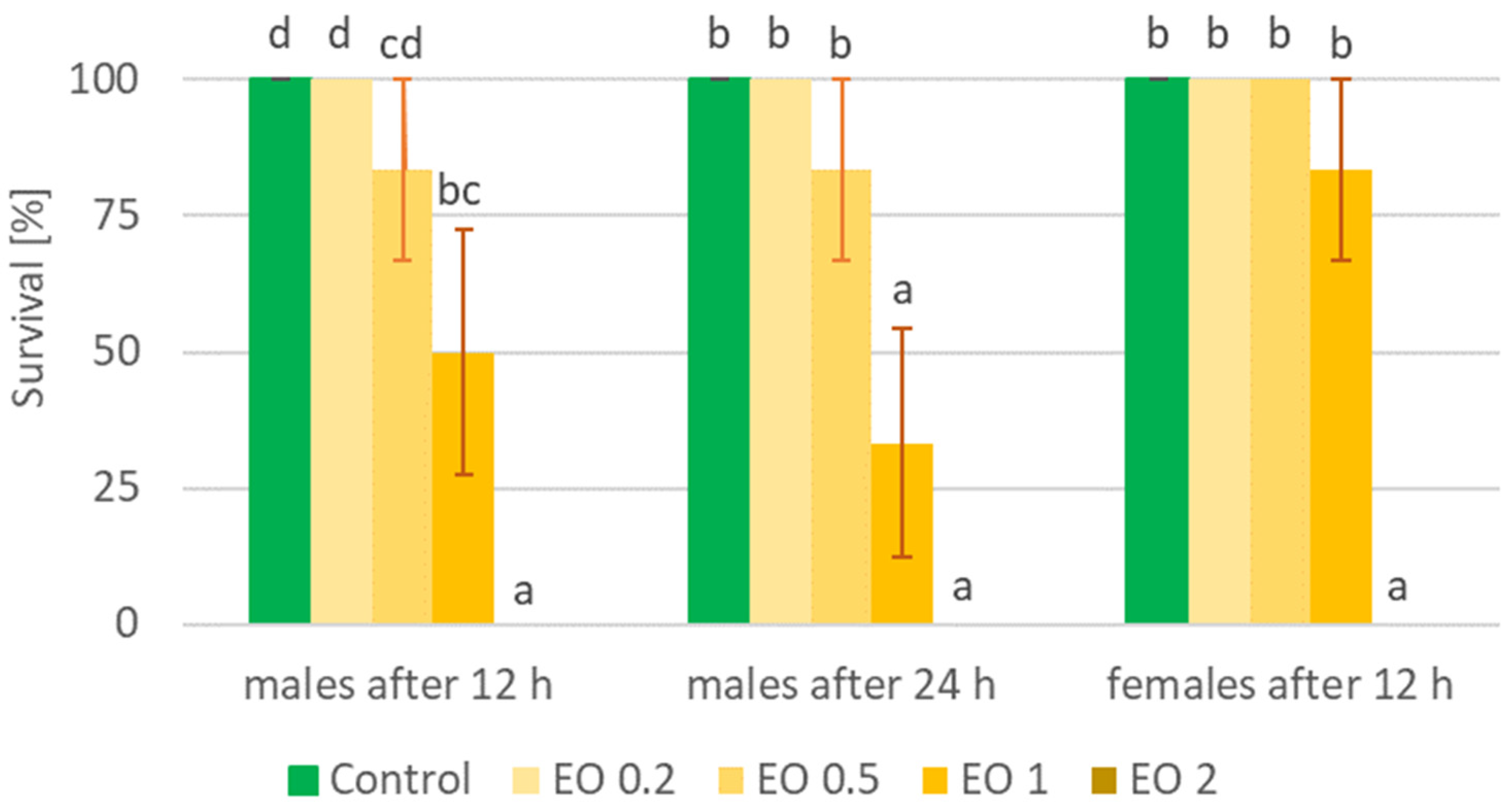
2.2. Colorado Potato Beetle
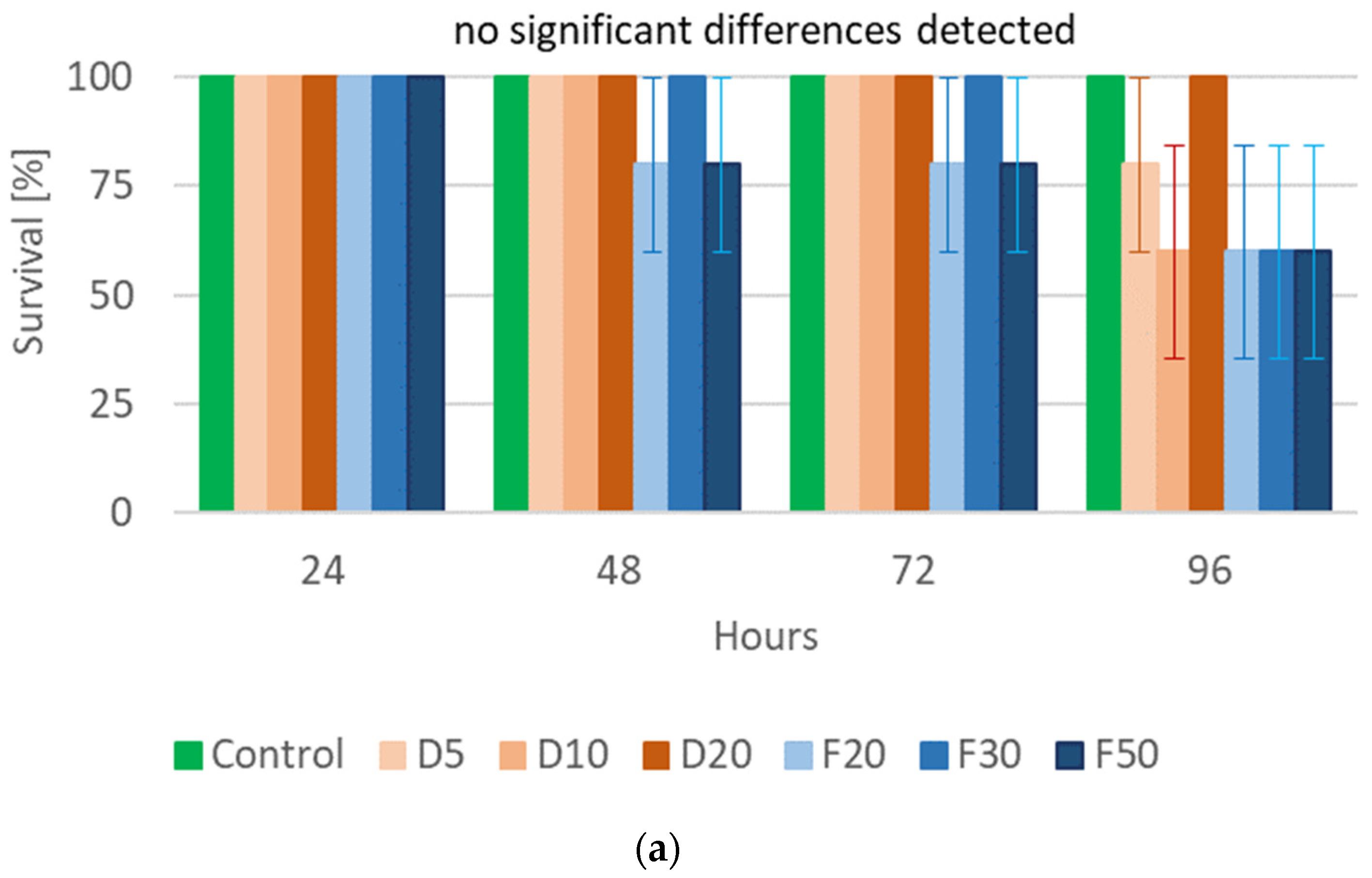
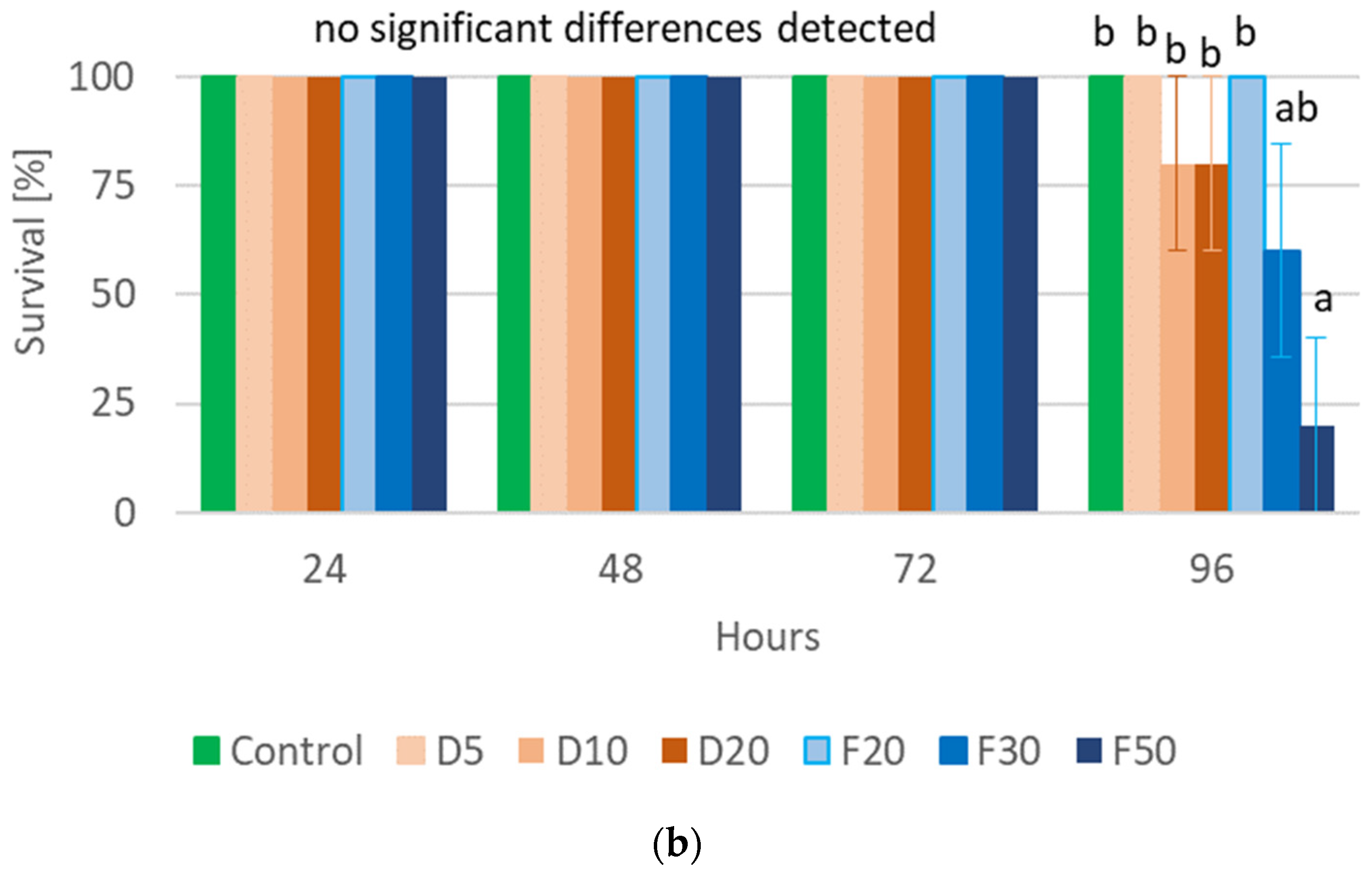
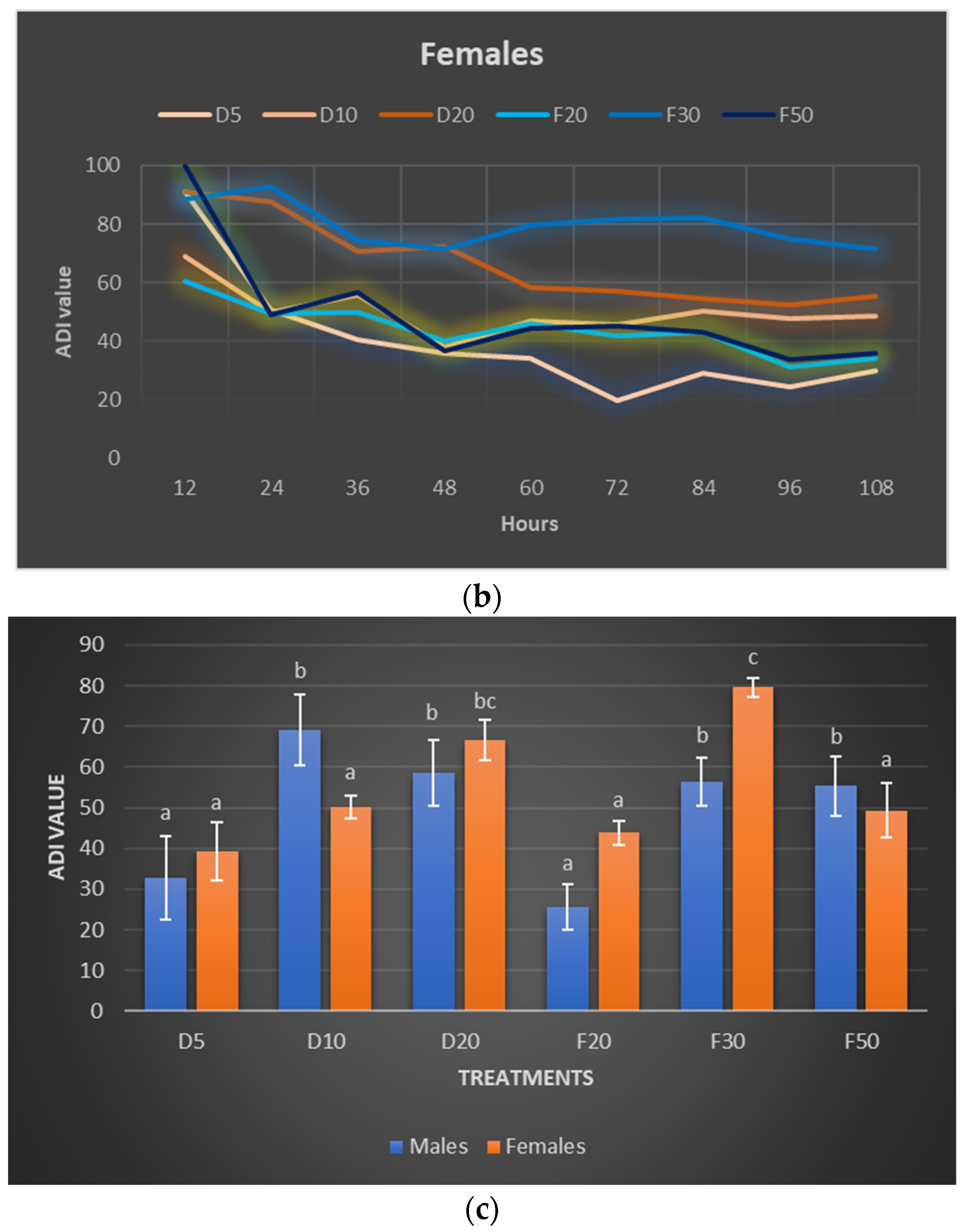
2.3. Pea Leaf Weevil
2.4. Asian Lady Beetle
2.5. Chemical Composition of EOTO
3. Discussion
3.1. Chemical Composition of EOTO
3.2. Black Bean Aphid
3.3. Colorado Potato Beetle
3.4. Pea Leaf Weevil
3.5. Asian Lady Beetle
4. Materials and Methods
4.1. EOTO and WETO Preparation, Chemical Composition Analysis of EOTO, and Experimental Design
4.2. Insect Treatment
4.2.1. Black Bean Aphid
4.2.2. Colorado Potato Beetle
4.2.3. Pea Leaf Weevil
4.2.4. Asian Lady Beetle
4.3. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ikbal, C.; Pavela, R. Essential oils as active ingredients of botanical insecticides against aphids. J. Pest Sci. 2019, 92, 971–986. [Google Scholar] [CrossRef]
- Lupia, C.; Castagna, F.; Bava, R.; Naturale, M.D.; Zicarelli, L.; Marrelli, M.; Statti, G.; Tilocca, B.; Roncada, P.; Britti, D.; et al. Use of Essential Oils to Counteract the Phenomena of Antimicrobial Resistance in Livestock Species. Antibiotics 2024, 13, 163. [Google Scholar] [CrossRef] [PubMed]
- Caruntu, S.; Ciceu, A.; Olah, N.K.; Don, I.; Hermenean, A.; Cotoraci, C. Thuja occidentalis L. (Cupressaceae): Ethnobotany, Phytochemistry and Biological Activity. Molecules 2020, 25, 5416. [Google Scholar] [CrossRef] [PubMed]
- Lebbal, S.; Hedjazi, N.; Tabti, I.; Ouarghi, H.; Zeraib, A. Aphicidal activity of plant extracts against larvae of Aphis fabae (Scopoli, 1763) (Hemiptera: Aphididae). Acta Entomol. Serbica 2018, 23, 1–7. [Google Scholar] [CrossRef]
- Alloui-Griza, R.; Cherif, A.; Attia, S.; Francis, F.; Lognay, G.C.; Grissa-Lebdi, K. Lethal Toxicity of Thymus capitatus Essential Oil Against Planococcus citri (Hemiptera: Pseudococcidae) and its Coccinellid Predator Cryptolaemus montrouzieri (Coleoptera: Coccinellidae). J. Entomol. Sci. 2022, 57, 425–435. [Google Scholar] [CrossRef]
- Edison, A.; Michelbach, A.; Sowade, D.; Kertzel, H.; Schmidt, L.; Schäfer, M.; Lysander, M.; Nauen, R.; Duchen, P.; Xu, S.Q. Evidence of active oviposition avoidance to systemically applied imidacloprid in the Colorado potato beetle. Insect Sci. 2024, 0, 1–12. [Google Scholar] [CrossRef]
- Baser, K.H.C.; Buchbauer, G. Handbook of Essential Oils: Science, Technology, and Applications, 3rd ed.; Baser, K.H.C., Ed.; CRC Press: Boca Raton, FL, USA, 2024. [Google Scholar]
- Khammassi, M.; Ben Ayed, R.; Khedhiri, S.; Souihi, M.; Hanana, M.; Amri, I.; Hamrouni, L. Crude extracts and essential oil of Platycladus orientalis (L.) Franco: A source of phenolics with antioxidant and antibacterial potential as assessed through a chemometric approach. Turk. J. Agric. For. 2022, 46, 477–487. [Google Scholar] [CrossRef]
- Naser, B.; Bodinet, C.; Tegtmeier, M.; Lindequist, U. Thuja occidentalis (Arbor vitae): A review of its pharmaceutical, pharmacological and clinical properties. Evid. Based Complement. Alternat. Med. 2005, 2, 69–78. [Google Scholar] [CrossRef]
- Pavela, R. Essential oils from Foeniculum vulgare Miller as a safe environmental insecticide against the aphid Myzus persicae Sulzer. Environ. Sci. Pollut. Res. 2018, 25, 10904–10910. [Google Scholar] [CrossRef]
- Benelli, G.; Pavela, R.; Zorzetto, C.; Sanchez-Mateo, C.C.; Santini, G.; Canale, A.; Maggi, F. Insecticidal activity of the essential oil from Schizogyne sericea (Asteraceae) on four insect pests and two non-target species. Entomol. Gen. 2019, 39, 9–18. [Google Scholar] [CrossRef]
- Parreira, D.S.; Alcantara-de la Cruz, R.; Zanuncio, J.C.; Lemes, P.G.; Rolim, G.D.; Barbosa, L.R.; Leite, G.L.D.; Serrao, J.E. Essential oils cause detrimental effects on biological parameters of Trichogramma galloi immatures. J. Pest Sci. 2018, 91, 887–895. [Google Scholar] [CrossRef]
- Kéïta, S.M.; Vincent, C.; Schmidt, J.-P.; Arnason, J.T. Insecticidal effects of Thuja occidentalis (Cupressaceae) essential oil on Callosobruchus maculatus [Coleoptera: Bruchidae]. Can. J. Plant Sci. 2001, 81, 173–177. [Google Scholar] [CrossRef]
- Akkol, E.K.; Ilhan, M.; Demirel, M.A.; Keles, H.; Tumen, I.; Suntar, I. Thuja occidentalis L. and its active compound,α-thujone: Promising effect in the treatment of polycystic ovary syndrome without inducingosteroporosis. J. Ethnopharmacol. 2015, 168, 25–30. [Google Scholar] [CrossRef]
- Thakur, M.; Sobti, R.; Kaur, T. Medicinal and biological potential of Thuja occidentalis: A comprehensive review. Asian Pac. J. Trop. Med. 2023, 16, 148–161. [Google Scholar] [CrossRef]
- Lis, A.; Liśkiewicz, R.; Krajewska, A. Comparison of chemical composition of essentials oils from different parts of Thuja occidentalis L. ‘Brabant’ and T. occidentalis L. ‘Smaragd’. Herba Polonica 2016, 62, 20–27. [Google Scholar] [CrossRef]
- Tsiri, D.; Graikou, K.; Poblocka-Olech, L.; Krauze-Baranowska, M.; Spyropoulos, C.; Chinou, I. Chemosystematic value of the essential oil composition of Thuja species cultivated in Poland—Antimicrobial activity. Molecules 2009, 14, 4707–4715. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Pathak, A.; Dikshit, A.; Mishra, R.K. Antibacterial evaluation of essential oils of Juniperus communis, Pinus roxburghii and Thuja occidentalis against Escherichia coli. Biochem. Cell. Arch. 2018, 18, 241–244. [Google Scholar]
- Song, H.-J.; Yong, S.-H.; Kim, H.-G.; Kim, D.-H.; Park, K.-B.; Shin, K.-C.; Choi, M.-S. Insecticidal Activity against Myzus persicae of Terpinyl Acetate and Bornyl Acetate in Thuja occidentalis Essential Oil. Horticulturae 2022, 8, 969. [Google Scholar] [CrossRef]
- Hosseinzadeh, J.; Farazmand, H.; Karimpour, Y. Insecticidal effect of Thuja occidentalis L. essential oilon adults of Lasioderma serricorne F. (Anobiidae) under laboratory conditions. Iran J. Med. Aromatic Pl. 2014, 30, 123–133. [Google Scholar]
- Abdelgaleil, S.A.M.; Badawy, M.E.I.; Shawir, M.S.; Mohamed, M.I.E. Chemical Composition, Fumigant and Contact Toxicities of Essential Oils Isolated from Egyptian Plants against the Stored Grain Insects Sitophilus oryzae L. and Tribolium castaneum (Herbst). Egypt. J. Biol. Pest Control 2015, 25, 639–647. [Google Scholar]
- Brari, J.; Thakur, D.R. Larvicidal effects of eight essential oils against Plodia interpunctella and Tribolium castaneum, serious pests of stored products worldwide. J. Entomol. Zool. Stud. 2018, 6, 738–742. [Google Scholar]
- Martynov, V.O.; Titov, O.G.; Kolombar, T.M.; Brygadyrenko, V.V. Influence of essential oils of plants on the migration activity of Tribolium confusum (Coleoptera, Tenebrionidae). Biosyst. Divers. 2019, 27, 177–185. [Google Scholar] [CrossRef]
- Pavela, R. Insecticidal properties of several essential oils on the house fly (Musca domestica L.). Phytother. Res. 2008, 22, 274–278. [Google Scholar] [CrossRef] [PubMed]
- Benelli, G.; Flamini, G.; Canale, A.; Cioni, P.L.; Conti, B. Toxicity of some essential oil formulations against the Mediterranean fruit fly Ceratitis capitata (Wiedemann) (Diptera Tephritidae). Crop Protect. 2012, 42, 223–229. [Google Scholar] [CrossRef]
- Pavela, R. Insecticidal activity of some essential oils against larvae of Spodoptera littoralis. Fitoterapia 2005, 76, 691–696. [Google Scholar] [CrossRef] [PubMed]
- Bai, L.; Wang, W.; Hua, J.; Guo, Z.; Luo, S. Defensive functions of volatile organic compounds and essential oils from northern white-cedar in China. BMC Plant Biol. 2020, 20, 500. [Google Scholar] [CrossRef] [PubMed]
- Fitsev, I.M.; Nikitin, E.N.; Rakhmaeva, A.M.; Terenzhev, D.A.; Sakhno, T.M.; Nasybullina, Z.R. Chemical Composition of Cupressus sempervirens L. and Thuja occidentalis L. Essential Oils and Their Activity against Phytopathogenic Fungi. Uchenye Zap. Kazan. Univ.-Seriya Estestv. Nauk. 2022, 164, 392–407. [Google Scholar] [CrossRef]
- Wróblewska-Kurdyk, A.; Gniłka, R.; Dancewicz, K.; Grudniewska, A.; Wawrzenczyk, C.; Gabrys, B. Β-Thujone and its derivatives modify the probing behavior of the peach potato aphid. Molecules 2019, 24, 1847. [Google Scholar] [CrossRef]
- Deepika, T.; Reena, A.; Sourabh, J. Antimicrobial, antioxidant and phytochemical investigation of Thuja occidentalis (Arbor vitae) leave extract. GSC Biol. Pharm. Sci. 2020, 12, 108–116. [Google Scholar] [CrossRef]
- Benahmed Djilali, A.; Mehraz, R.; Bouacem, K.; Benseddik, A.; Moualek, I.; Nabiev, M.; Benzara, A. Bioactive Substances of Cydonia oblonga Fruit: Insecticidal Effect of Tannins on Tribuliumm confusum. Int. J. Fruit Sci. 2021, 21, 721–731. [Google Scholar] [CrossRef]
- Oulebsir-Mohandkaci, H.; Aissa, A.B.; Badaoui, S.; Bouyahiaoui, H.; Kaki, S.A.; Mohammedi, A. Comparative study of the toxicity of phenolic compounds of coriander (Coriandrum sativum) and false fennel (Aneth graveolens) on Galleria mellonella (Lepidoptera, Pyralidae). Euro-Mediterr. J. Environ. Integr. 2018, 3, 30. [Google Scholar] [CrossRef]
- Petchidurai, G.; Sahayaraj, K.; Al-Shuraym, L.A.; Albogami, B.Z.; Sayed, S.M. Insecticidal Activity of Tannins from Selected Brown Macroalgae against the Cotton Leafhopper Amrasca devastans. Plants 2023, 12, 3188. [Google Scholar] [CrossRef]
- Mouna, M.; Khadra, B.; Assia, B.; Salim, L. Insecticidal effect of two aqueous extracts from the leaves of Salvia officinalis and Eucalyptus camaldulensis against Aphis fabae. J. Entomol. Res. 2021, 45, 268–279. [Google Scholar] [CrossRef]
- de Melo Burger, M.C.; Matos, A.P.; Sabbag Cunha, G.O.; Bernardo, A.R.; Severo Menezes, A.C.; Vieira, P.C.; Fernandes, J.B.; das Gracas Fernandes da Silva, M.F. Flavonoids and Insecticidal Activity Against Spodoptera frugiperda of Myrsine coriacea (Primulaceae). Revista Virtual Quimica 2021, 13, 953–958. [Google Scholar] [CrossRef]
- Zhang, T.; Tian, C.-Y.; Zhang, J.; An, Q.; Yi, P.; Yuan, C.-M.; Zhang, Z.-K.; Zhao, L.-H.; Hao, X.-J.; Hu, Z.-X. Quinolizidine Alkaloids and Isoflavones from the Herb of Thermopsis lupinoides and Their Antiviral, Antifungal, and Insecticidal Activities. J. Agr. Food Chem. 2024, 72, 5047–5061. [Google Scholar] [CrossRef] [PubMed]
- Souto, A.L.; Sylvestre, M.; Tolke, E.D.; Tavares, J.F.; Barbosa-Filho, J.M.; Cebrian-Torrejon, G. Plant-Derived Pesticides as an Alternative to Pest Management and Sustainable Agricultural Production: Prospects, Applications and Challenges. Molecules 2021, 26, 4835. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Stahl, A.; Rostas, M.; Will, T. Temporal and species-specific resistance of sugar beet to green peach aphid and black bean aphid: Mechanisms and implications for breeding. Pest Manag. Sci. 2023, 80, 404–413. [Google Scholar] [CrossRef] [PubMed]
- Shannag, H.K.; Ababneh, J.A. Influence of black bean aphid, Aphis fabae Scopoli on growth rates of faba bean. World J. Agric. Sci. 2007, 3, 344–349. [Google Scholar]
- Hanavan, R.P.; Bosque-Pérez, N.A. Effects of tillage practices on pea leaf weevil (Sitona lineatus L., Coleoptera: Curculionidae) biology and crop damage: A farm-scale study in the US Pacific Northwest. Bull. Entomol. Res. 2012, 102, 682–691. [Google Scholar] [CrossRef]
- Lohaus, K.; Vidal, S. Abundance of Sitona lineatus L. (Col., Curculionidae) in peas (Pisum sutivum L.): Effects on yield parameters and nitrogen balance. Crop Protect. 2010, 29, 283–289. [Google Scholar] [CrossRef]
- EPPO. Leptinotarsa decemlineata. EPPO Datasheets on Pests Recommended for Regulation. 2024. Available online: https://gd.eppo.int (accessed on 8 March 2024).
- Skryabin, K. Do Russia and Eastern Europe need GM plants? New Biotechnol. 2010, 27, 593–595. [Google Scholar] [CrossRef]
- Vandereycken, A.; Durieux, D.; Joie, E.; Francis, F.; Haubruge, E.; Verheggen, F.J. Aphid species and associated natural enemies in field crops: What about the invasive ladybird Harmonia axyridis (Coleoptera: Coccinellidae)? Entomol. Faun. 2015, 68, 3–15. [Google Scholar]
- Lawrence, B.M. Progress in essential oil. Perfum. Flavorist 1995, 20, 103–104. [Google Scholar]
- Lawrence, B.M. Progress in essential oil. Perfum. Flavorist 1993, 18, 54–55. [Google Scholar]
- Nukenine, E.N.; Adler, C.; Reichmuth, C. Bioactivity of fenchone and Plectranthus glandulosus oil against Prostephanus truncatus and two strains of Sitophilus zeamais. J. Appl. Entom. 2010, 134, 132–141. [Google Scholar] [CrossRef]
- Abdel-Baki, A.-A.S.; Aboelhadid, S.M.; Sokmen, A.; Al-Quraishy, S.; Hassan, A.O.; Kamel, A.A. Larvicidal and pupicidal activities of Foeniculum vulgare essential oil, trans-anethole and fenchone against house fly Musca domestica and their inhibitory effect on acetylcholinestrase. Entom. Res. 2021, 51, 568–577. [Google Scholar] [CrossRef]
- Gospodarek, J.; Endalamew, A.; Worsdale, M.; Pasmionka, I.B. Effects of Artemisia dracunculus L. Water Extracts on Selected Pests and Aphid Predator Coccinella septempunctata L. Agronomy 2022, 12, 788. [Google Scholar] [CrossRef]
- Gospodarek, J.; Krajewska, A.; Pasmionka, I.B. Contact and Gastric Effect of Peppermint Oil on Selected Pests and Aphid Predator Harmonia axyridis Pallas. Molecules 2023, 28, 4647. [Google Scholar] [CrossRef]
- Sayed, S.; Soliman, M.M.; Al-Otaibi, S.; Hassan, M.M.; Elarrnaouty, S.A.; Abozeid, S.M.; El-Shehawi, A.M. Toxicity, Deterrent and Repellent Activities of Four Essential Oils on Aphis punicae (Hemiptera: Aphididae). Plants 2022, 11, 463. [Google Scholar] [CrossRef] [PubMed]
- Heydari, M.; Amirjani, A.; Bagheri, M.; Sharifian, I.; Sabahi, Q. Eco-friendly pesticide based on peppermint oil nanoemulsion: Preparation, physicochemical properties, and its aphicidal activity against cotton aphid. Environ. Sci. Pollut. Res. 2020, 27, 6667–6679. [Google Scholar] [CrossRef]
- Albouchi, F.; Ghazouani, N.; Souissi, R.; Abderrabba, M.; Boukhris-Bouhachem, S. Aphidicidal activities of Melaleuca styphelioides Sm. essential oils on three citrus aphids: Aphis gossypii Glover; Aphis spiraecola Patch and Myzus persicae (Sulzer). S. Afr. J. Bot. 2018, 117, 149–154. [Google Scholar] [CrossRef]
- Kwiecień, N.; Gospodarek, J.; Boligłowa, E. The Effects of Water Extracts from Tansy on Pea Leaf Weevil and Black Bean Aphid. J. Ecol. Eng. 2020, 21, 220–227. [Google Scholar] [CrossRef] [PubMed]
- Binias, B.; Gospodarek, J.; Rusin, M. Effect of water extract from mint on selected crop pests feeding and survival. J. Ecol. Eng. 2017, 18, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Rusin, M.; Gospodarek, J. The Effects of Water Extracts from Lemon Balm on Pea Leaf Weevil and Black Bean Aphid Behaviour. J. Ecol. Eng. 2018, 19, 139–145. [Google Scholar] [CrossRef] [PubMed]
- Nation, J.L. Insect Physiology and Biochemistry, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2008; 560p. [Google Scholar] [CrossRef]
- Feng, Y.-X.; Wang, Y.; Chen, Z.-Y.; Guo, S.-S.; You, C.-X.; Du, S.-S. Efficacy of bornyl acetate and camphene from Valeriana officinalis essential oil against two storage insects. Environ. Sci. Poll. Res. 2019, 26, 16157–16165. [Google Scholar] [CrossRef] [PubMed]
- Rafiee-Dastjerdi, H.; Khorrami, F.; Ganbalani, G.N.; Fathi, A.A.; Esmaielpour, B. Efficacy of some medicinal plant extracts and essential oils against Colorado potato beetle, Leptinotarsa decemlineata Say (Coleoptera: Chrysomelidae). Arch. Phytopathol. Plant Prot. 2014, 47, 1175–1178. [Google Scholar] [CrossRef]
- Sajfrtova, M.; Sovova, H.; Karban, J.; Rochova, K.; Pavela, R.; Barnet, M. Effect of separation method on chemical composition and insecticidal activity of Lamiaceae isolates. Ind. Crops Prod. 2013, 47, 69–77. [Google Scholar] [CrossRef]
- Kostic, M.; Drazic, S.; Popovic, Z.; Stankovic, S.; Sivcev, I.; Zivanovic, T. Developmental and feeding alternations in Leptinotarsa decemlineata Say. (Coleoptera: Chrysomelidae) caused by Salvia officinalis L. (Lamiaceae) essential oil. Biotechnol. Biotechnol. Equip. 2007, 21, 426–430. [Google Scholar] [CrossRef]
- Biniaś, B.; Gospodarek, J.; Rusin, M. Effect of fennel water extracts on reduction of feeding of pea leaf weevil. J. Ecol. Eng. 2016, 17, 192–197. [Google Scholar] [CrossRef]
- Kimbaris, A.C.; Papachristos, D.P.; Michaelakis, A.; Martinou, A.F.; Polissiou, M.G. Toxicity of plant essential oil vapours to aphid pests and their coccinellid predators. Biocontrol Sci. Technol. 2010, 20, 411–422. [Google Scholar] [CrossRef]
- Abdelatti, Z.A.S.; Hartbauer, M. Plant oil mixtures as a novel botanical pesticide to control gregarious locusts. J. Pest Sci. 2020, 93, 341–353. [Google Scholar] [CrossRef]
- Ebadollahi, A.; Setzer, W.N. Evaluation of the Toxicity of Satureja intermedia C. A. Mey Essential Oil to Storage and Greenhouse Insect Pests and a Predator Ladybird. Foods 2020, 9, 712. [Google Scholar] [CrossRef] [PubMed]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography/Mass Spectrometry; Allured Publishing Corporation: Carol Stream, IL, USA, 2007; Volume 456, pp. 544–545. [Google Scholar]
- Al-Antary, T.M.; Belghasem, I.H.; Alaraj, S.A. Effect of mint oil against the green peach aphid Myzus persicae Sulzer (Homop-tera: Aphididae) using four solvents. Adv. Environ. Biol. 2017, 11, 61–67. [Google Scholar]
- Finney, D. Probit Analysis, 3rd ed.; Cambridge University Press: Cambridge, UK, 1971. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Hours | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| 6 | 18 | 30 | 42 | 54 | 66 | 78 | 90 | 102 | |
| EOTO | |||||||||
| Control | 100.00 (±0.00) a* | 100.00 (±0.00) a | 97.50 (±2.50) a | 97.50 (±2.50) a | 95.00 (±2.89) b | 95.00 (±2.89) b | 87.50 (±6.29) b | 67.50 (±8.54) ab | 40.00 (±17.80) a |
| EO 0.05 | 98.00 (±2.00) a | 94.00 (±4.00) a | 94.00 (±4.00) a | 92.00 (±3.74) a | 92.00 (±3.74) b | 84.36 (±2.32) b | 74.55 (±3.90) b | 72.55 (±3.71) b | 50.55 (±16.90) a |
| EO 0.1 | 96.00 (±2.45) a | 94.00 (±4.00) a | 94.00 (±4.00) a | 90.00 (±6.32) a | 87.78 (±5.82) b | 85.78 (±7.46) b | 81.78 (±6.57) b | 73.78 (±10.21) b | 61.78 (±15.84) a |
| EO 0.2 | 96.00 (±4.00) a | 96.00 (±4.00) a | 96.00 (±4.00) a | 96.00 (±4.00) a | 92.00 (±4.90) b | 88.00 (±5.83) b | 70.00 (±8.37) ab | 48.00 (±12.41) ab | 42.00 (±14.63) a |
| EO 0.5 | 92.00 (±5.83) a | 92.00 (±5.83) a | 84.00 (±6.78) a | 74.00 (±10.30) a | 64.00 (±13.27) a | 56.00 (±12.08) a | 50.00 (±11.40) a | 38.00 (±9.70) a | 32.00 (±9.70) a |
| WETOs | |||||||||
| Control | 100.00 (±0.00) *a | 96.67 (±3.33) a | 95.00 (±3.42) a | 95.00 (±3.42) a | 90.00 (±6.83) a | 83.33 (±9.19) a | 73.33 (±10.85) a | 61.67 (±13.52) a | 51.67 (±14.24) a |
| D2 | 98.33 (±1.67) a | 93.33 (±3.33) a | 91.67 (±4.01) a | 90.00 (±4.47) a | 68.33 (±10.46) a | 65.00 (±10.57) a | 43.15 (±13.94) a | 34.44 (±10.53) a | 27.59 (±8.98) a |
| D5 | 100.00 (±0.00) a | 93.33 (±2.11) a | 93.33 (±2.11) a | 93.33 (±2.11) a | 78.33 (±7.03) a | 73.33 (±8.43) a | 63.33 (±12.56) a | 56.67 (±14.06) a | 40.00 (±15.49) a |
| D10 | 96.67 (±3.33) a | 96.67 (±3.33) a | 96.67 (±3.33) a | 95.00 (±3.42) a | 85.00 (±3.42) a | 71.67 (±8.33) a | 55.00 (±1 4.75) a | 50.00 (±12.91) a | 41.67 (±13.76) a |
| F10 | 95.00 (±3.42) a | 88.33 (±3.07) a | 88.33 (±3.07) a | 85.00 (±4.28) a | 78.33 (±4.77) a | 70.00 (±8.94) a | 60.00 (±9.66) a | 46.67 (±8.82) a | 35.00 (±10.57) a |
| F20 | 98.33 (±1.67) a | 96.67 (±2.11) a | 93.33 (±2.11) a | 91.67 (±3.07) a | 76.67 (±8.03) a | 70.00 (±10.65) a | 65.00 (±11.76) a | 51.67 (±10.14) a | 40.00 (±12.65) a |
| F30 | 96.82 (±2.02) a | 86.97 (±3.22) a | 85.45 (±3.98) a | 82.12 (±3.77) a | 75.76 (±6.16) a | 62.58 (±8.31) a | 54.55 (±9.47) a | 44.70 (±8.65) a | 34.85 (±7.29) a |
| Treatment | Hours | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| 6 | 18 | 30 | 42 | 54 | 66 | 78 | 90 | 102 | |
| EOTO | |||||||||
| Control | 100.00 (±0.00) b* | 100.00 (±0.00) c | 98.33 (±1.67) c | 98.33 (±1.67) c | 96.67 (±3.33) c | 93.33 (±4.22) c | 64.26 (±6.35) b | 49.26 (±8.81) b | 35.93 (±9.56) b |
| EO 0.05 | 100.00 (±0.00) b | 100.00 (±0.00) c | 100.00 (±0.00) c | 100.00 (±0.00) c | 100.00 (±0.00) c | 91.67 (±4.77) c | 80.00 (±6.83) b | 48.33 (±9.10) b | 28.33 (±5.43) b |
| EO 0.1 | 98.33 (±1.67) b | 98.33 (±1.67) c | 95.00 (±2.24) c | 95.00 (±2.24) c | 90.00 (±2.58) c | 83.33 (±5.58) c | 75.00 (±6.71) b | 53.33 (±8.82) b | 35.00 (±5.63) b |
| EO 0.2 | 71.67 (±18.33) b | 65.00 (±19.10) b | 55.00 (±19.62) b | 51.67 (±18.33) b | 39.81 (±14.71) b | 29.81 (±16.19) b | 21.30 (±13.94) a | 8.70 (±5.60) a | 3.52 (±2.23) a |
| EO 0.5 | 13.33 (±13.33) a | 13.33 (±13.33) a | 13.33 (±13.33) a | 6.67 (±6.67) a | 6.67 (±6.67) a | 0.00 (±0.00) a | 0.00 (±0.00) a | 0.00 (±0.00) a | 0.00 (±0.00) a |
| WETOs | |||||||||
| Control | 100.00 (±0.00) a | 100.00 (±0.00) a | 100.00 (±0.00) a | 98.33 (±1.67) a | 98.33 (±1.67) c | 93.33 (±2.11) b | 68.33 (±4.01) c | 43.33 (±3.33) c | 16.67 (±3.33) b |
| D2 | 96.67 (±2.11) a | 96.67 (±2.11) a | 93.33 (±2.11) a | 91.67 (±3.07) a | 80.00 (±2.58) b | 53.33 (±10.85) a | 23.33 (±11.16) ab | 8.33 (±4.77) a | 1.67 (±1.67) a |
| D5 | 98.33 (±1.67) a | 95.00 (±3.42) a | 95.00 (±3.42) a | 88.33 (±3.07) a | 76.67 (±4.22) ab | 58.33 (±8.33) a | 30.00 (±7.30) ab | 13.33 (±5.58) a | 8.33 (±4.77) ab |
| D10 | 100.00 (±0.00) a | 98.33 (±2.67) a | 96.67 (±2.11) a | 90.00 (±2.58) a | 73.33 (±7.15) ab | 51.67 (±8.72) a | 25.00 (±8.47) ab | 13.33 (±4.94) a | 1.67 (±1.67) a |
| F10 | 96.67 (±2.11) a | 95.00 (±2.24) a | 91.67 (±1.67) a | 83.33 (±4.22) a | 78.33 (±4.01) ab | 60.00 (±12.65) a | 46.67 (±12.29) bc | 33.33 (±10.22) bc | 13.33 (±6.15) b |
| F20 | 98.33 (±1.67) a | 96.67 (±2.11) a | 91.67 (±4.77) a | 90.00 (±4.47) a | 65.00 (±6.71) a | 41.67 (±8.33) a | 16.67 (±6.67) a | 3.33 (±2.11) a | 1.67 (±1.67) a |
| F30 | 100.00 (±0.00) a | 98.33 (±1.67) a | 95.00 (±3.42) a | 93.33 (±3.33) a | 85.00 (±6.19) bc | 63.33 (±6.67) a | 31.67 (±9.46) ab | 18.33 (±6.54) ab | 3.33 (±3.33) a |
| Life Stage | Hours | LC50 (%) | 95% Confidence Limits | Slope * | (X2) ** | |
|---|---|---|---|---|---|---|
| Nymphs | Lower | Upper | ||||
| 30 | 1.0989 | 0.7083 | 5.8427 | 1.4417 | 26.8668 1 | |
| 42 | 0.8267 | 0.4358 | 1.8103 | 1.7420 | 35.0288 | |
| 54 | 0.6501 | 0.4485 | 1.0293 | 2.1088 | 34.4596 | |
| 66 | 0.5883 | 0.4661 | 0.8729 | 1.8802 | 29.8071 1 | |
| 78 | 0.5053 | 0.3811 | 0.8452 | 1.4368 | 25.3154 1 | |
| 90 | 0.3132 | 0.1943 | 0.4843 | 1.6663 | 36.3392 | |
| 102 | 0.1050 | 0.0094 | 0.5457 | 0.4781 | 73.1526 | |
| Wingless females | 6 | 0.3237 | 0.2204 | 0.4342 | 6.1580 | 137.0640 |
| 18 | 0.3095 | 0.2043 | 0.4229 | 5.9938 | 143.6515 | |
| 30 | 0.2849 | 0.1794 | 0.3996 | 5.6576 | 148.4458 | |
| 42 | 0.2453 | 0.1614 | 0.3413 | 7.4098 | 123.1570 | |
| 54 | 0.2176 | 0.1342 | 0.3121 | 7.5166 | 147.9525 | |
| 66 | 0.1625 | 0.1337 | 0.1948 | 11.6526 | 53.5328 | |
| 78 | 0.1376 | 0.1079 | 0.1703 | 9.8867 | 51.4601 | |
| 90 | 0.0713 | 0.0302 | 0.1030 | 7.4935 | 36.4395 | |
| 102 | 0.0097 | −0.0797 | 0.0451 | 6.7504 | 19.5659 1 | |
| Treatment | Hours | Body Weight Change Compared to T0 (mg) | |||
|---|---|---|---|---|---|
| 24 | 48 | 72 | 96 | ||
| Females | |||||
| Control | 256.27 (±63.59) a* | 305.01 (±60.28) a | 572.75 (±59.93) a | 596.50 (±73.84) a | −60.50 (±5.75) a |
| EO 0.2 | 254.06 (±43.50) a | 268.88 (±51.16) a | 484.26 (±66.09) a | 497.24 (±71.77) a | −50.00 (±13.57) a |
| EO 0.5 | 282.18 (±32.67) a | 282.18 (±32.67) a | 605.85 (±72.69) a | 664.86 (±67.10) a | −74.80 (±4.85) a |
| EO 1 | 216.57 (±52.85) a | 245.68 (±54.03) a | 687.20 (±109.95) a | 779.90 (±121.06) a | −69.60 (±7.35) a |
| EO 2 | 297.56 (±63.17) a | 310.93 (±0.00) a | 474.01 (±0.00) a | 474.01 (±0.00) a | −37.00 (±0.00) a |
| Males | |||||
| Control | 227.17 (±48.93) a | 254.12 (±53.95) a | 557.95 (±43.98) a | 606.25 (±48.29) a | −15.33 (±3.42) a |
| EO 0.2 | 204.49 (±32.61) a | 204.49 (±32.61) a | 491.10 (±72.10) a | 505.83 (±72.10) a | −18.00 (±4.07) a |
| EO 0.5 | 224.89 (±51.48) a | 220.58 (±62.82) a | 459.62 (±84.54) a | 499.30 (±117.30) a | −5.60 (±3.04) a |
| EO 1 | 276.17 (±41.96) a | 276.17 (±41.96) a | 948.69 (±192.21) b | 1020.31 (±166.52) b | −39.00 (±21.18) a |
| EO 2 | 223.14 (±27.36) a | 238.69 (±24.03) a | 514.42 (±58.98) a | 555.06 (±86.41) a | −16.17 (±3.94) a |
| Treatment | Hours | Body Weight Change Compared to T0 (mg) | |||
|---|---|---|---|---|---|
| 24 | 48 | 72 | 96 | ||
| Females | |||||
| Control | 282.79 (±35.04) a* | 315.54 (±42.55) a | 383.25 (±46.81) a | 453.01 (±51.56) a | −56.50 (±7.60) a |
| D5 | 199.22 (±65.02) a | 241.34 (±56.61) a | 281.55 (±43.31) a | 367.36 (±35.14) a | −33.33 (±4.67) a |
| D10 | 224.47 (±66.57) a | 279.83 (±65.54) a | 330.86 (±70.06) a | 383.67 (±84.24) a | −51.00 (±12.29) a |
| D20 | 206.30 (±46.25) a | 296.58 (±49.27) a | 368.88 (±66.91) a | 516.57 (±106.84) a | −39.60 (±16.67) a |
| F20 | 389.55 (±85.10) a | 407.40 (±91.22) a | 565.50 (±98.10) a | 603.91 (±126.79) a | −27.33 (±6.39) a |
| F30 | 340.83 (±63.96) a | 376.81 (±42.49) a | 400.99 (±30.37) a | 444.08 (±20.63) a | −59.67 (±1.45) a |
| F50 | 271.97 (±94.64) a | 283.04 (±93.00) a | 273.47 (±88.29) a | 407.50 (±129.42) a | −29.00 (±6.00) a |
| Males | |||||
| Control | 248.05 (±52.35) a | 254.70 (±49.63) a | 266.01 (±53.76) a | 295.88 (±48.87) a | 2.25 (±5.98) a |
| D5 | 174.16 (±33.25) a | 184.95 (±32.50) a | 188.44 (±33.09) a | 246.32 (±42.31) a | −2.20 (±3.02) a |
| D10 | 179.99 (±40.17) a | 194.16 (±32.40) a | 213.47 (±34.93) a | 231.83 (±38.46) a | −12.33 (±6.57) a |
| D20 | 212.78 (±97.23) a | 216.68 (±99.82) a | 235.76 (±109.32) a | 260.09 (±123.44) a | 2.50 (±5.56) a |
| F20 | 260.82 (±24.00) a | 261.52 (±22.30) a | 270.52 (±27.24) a | 295.15 (±38.13) a | −1.40 (±3.01) a |
| F30 | 226.99 (±44.51) a | 229.99 (±41.51) a | 233.18 (±47.72) a | 294.83 (±78.61) a | −1.00 (±2.00) a |
| F50 | 238.90 (±50.49) a | 239.20 (±52.49) a | 262.18 (±40.29) a | 317.47 (±38.39) a | 2.00 (±0.00) a |
| Treatment | Hours | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| 12 | 24 | 36 | 48 | 60 | 72 | 84 | 96 | 108 | |
| Males | |||||||||
| Control | 7.76 (±2.82) b* | 17.66 (±3.22) c | 43.28 (±3.71) b | 61.61 (±6.95) b | 66.82 (±7.87) c | 74.38 (±6.28) b | 121.14 (±16.61) c | 140.91 (±20.05) c | 173.24 (±31.30) c |
| EO 0.2 | 8.44 (±4.40) b | 14.39 (±5.29) bc | 31.73 (±11.03) b | 39.25 (±13.09) ab | 54.15 (±15.53) bc | 69.84 (±21.39) b | 95.19 (±30.47) bc | 107.21 (±35.00) bc | 112.68 (±33.95) bc |
| EO 0.5 | 1.70 (±1.55) ab | 4.55 (±2.28) ab | 4.79 (±2.19) a | 14.13 (±4.33) a | 16.96 (±3.35) ab | 19.78 (±2.75) a | 39.85 (±8.03) ab | 42.35 (±7.89) ab | 51.77 (±9.20) ab |
| EO 1 | 0.07 (±0.07) a | 1.18 (±0.99) a | 1.20 (±0.90) a | 4.71 (±1.57) a | 4.71 (±1.57) a | 4.71 (±1.57) a | 4.71 (±1.57) a | 4.71 (±1.57) a | 4.71 (±1.57) a |
| EO 2 | 0.00 (±0.00) a | - | - | - | - | - | - | - | - |
| Females | |||||||||
| Control | 12.32 (±4.07) ab | 49.37 (±12.74) ab | 71.25 (±17.54) ab | 95.77 (±24.63) ab | 128.89 (±35.37) b | 145.58 (±36.67) ab | 240.85 (±69.66) a | 256.24 (±72.09) a | 285.58 (±76.95) b |
| EO 0.2 | 34.33 (±9.35) c | 64.62 (±12.16) b | 84.37 (±14.81) b | 131.29 (±24.57) b | 153.25 (±25.85) b | 175.23 (±30.05) b | 217.04 (±36.36) a | 240.37 (±44.14) a | 258.66 (±43.84) b |
| EO 0.5 | 24.91 (±5.72) bc | 78.00 (±13.67) b | 105.42 (±16.70) b | 131.58 (±21.27) b | 170.11 (±31.14) b | 185.76 (±29.89) b | 243.83 (±45.49) a | 314.28 (±52.46) a | 361.30 (±67.13) b |
| EO 1 | 8.44 (±3.80) a | 14.52 (±5.46) a | 25.73 (±11.55) a | 39.33 (±16.70) a | 41.74 (±14.51) a | 55.86 (±20.31) a | 65.27 (±20.42) a | 82.18 (±26.79) a | 81.25 (±21.18) a |
| EO 2 | 0.00 (±0.00) a | - | - | - | - | - | - | - | - |
| Treatment | Hours | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| 12 | 24 | 36 | 48 | 60 | 72 | 84 | 96 | 108 | |
| Males | |||||||||
| Control | 5.02 (±3.08) a* | 29.52 (±18.05) a | 62.14 (±17.54) c | 73.51 (±15.76) b | 119.25 (±42.78) a | 145.92 (±40.32) b | 160.04 (±40.76) a | 168.20 (±37.98) a | 210.85 (±34.21) a |
| D5 | 0.00 (±0.00) a | 6.99 (±4.31) a | 28.96 (±12.83) abc | 44.90 (±13.21) ab | 75.01 (±14.85) a | 85.66 (±15.18) ab | 134.60 (±25.23) a | 151.24 (±30.36) a | 171.59 (±30.97) a |
| D10 | 0.00 (±0.00) a | 0.00 (±0.00) a | 0.39 (±0.39) a | 11.48 (±5.86) a | 23.16 (±7.65) a | 31.01 (±5.89) a | 57.65 (±10.74) a | 74.93 (±16.15) a | 104.77 (±27.75) a |
| D20 | 0.00 (±0.00) a | 0.79 (±0.61) a | 12.40 (±5.17) ab | 20.09 (±6.33) a | 42.05 (±6.88) a | 46.46 (±4.98) a | 67.78 (±8.79) a | 82.53 (±9.48) a | 100.40 (±16.19) a |
| F20 | 1.26 (±1.26) a | 16.01 (±13.31) a | 36.02 (±17.58) bc | 39.39 (±19.17) ab | 67.44 (±35.36) a | 75.61 (±40.61) ab | 121.09 (±57.43) a | 159.63 (±74.91) a | 180.94 (±86.94) a |
| F30 | 0.08 (±0.08) a | 8.56 (±3.45) a | 18.76 (±4.88) ab | 29.04 (±5.87) a | 29.27 (±7.79) a | 33.51 (±7.99) a | 45.42 (±14.57) a | 77.68 (±13.13) a | 85.53 (±12.33) a |
| F50 | 0.00 (±0.00) a | 3.93 (±2.46) a | 17.90 (±7.97) ab | 19.47 (±7.95) a | 36.72 (±17.40) a | 45.50 (±19.67) a | 83.15 (±30.12) a | 86.28 (±32.61) a | 91.30 (±30.06) a |
| Females | |||||||||
| Control | 8.48 (±5.65) a | 53.99 (±15.79) b | 103.45 (±33.90) a | 137.75 (±30.51) b | 225.02 (±51.31) b | 253.28 (±62.06) b | 332.99 (±84.62) b | 358.76 (±93.85) a | 434.05 (±105.38) b |
| D5 | 0.39 (±0.39) a | 17.66 (±11.02) a | 43.94 (±25.98) a | 65.52 (±35.80) ab | 111.05 (±48.83) a | 169.93 (±68.82) ab | 184.01 (±68.28) ab | 219.31 (±82.50) a | 234.61 (±80.97) ab |
| D10 | 1.57 (±1.57) a | 18.05 (±6.36) a | 29.53 (±7.57) a | 61.99 (±21.29) ab | 81.56 (±32.63) a | 95.29 (±28.45) a | 110.64 (±38.24) a | 127.51 (±40.48) a | 150.61 (±57.65) a |
| D20 | 0.39 (±0.39) a | 3.53 (±3.03) a | 17.66 (±9.45) a | 21.98 (±9.44) a | 59.65 (±8.37) a | 69.07 (±13.23) a | 98.08 (±15.64) a | 111.80 (±13.22) a | 124.73 (±14.57) a |
| F20 | 2.09 (±2.09) a | 18.32 (±15.28) a | 34.54 (±16.32) a | 59.12 (±12.69) ab | 83.73 (±26.06) a | 104.66 (±29.04) a | 132.88 (±42.80) a | 188.33 (±72.24) a | 213.45 (±83.57) ab |
| F30 | 0.52 (±0.52) a | 2.09 (±1.38) a | 15.18 (±6.04) a | 23.03 (±8.62) a | 25.64 (±10.43) a | 25.64 (±10.43) a | 32.96 (±14.50) a | 51.28 (±23.71) a | 72.73 (±35.91) a |
| F50 | 0.00 (±0.00) a | 18.52 (±7.99) a | 28.64 (±8.97) a | 64.03 (±19.79) ab | 86.76 (±26.10) a | 96.17 (±26.60) a | 132.56 (±37.20) a | 177.80 (±49.76) a | 204.51 (±61.21) a |
| No. | Compound | RIlit 1 | RIlab | EOTO [%] | Lit [%] |
|---|---|---|---|---|---|
| 1 | Methylethyl butyrate | 833 | 0.2 | ||
| 2 | α-Tchujene | 926 | 920 | 1.0 | 0–0.7 |
| 3 | α-Pinene | 934 | 927 | 2.3 | 0.2–5.0 |
| 4 | α-Fenchene | 948 | 938 | 0.7 | 0–2.0 |
| 5 | Camphene | 950 | 939 | 0.8 | 0.1–3.0 |
| 6 | Sabinene | 970 | 964 | 12.9 | 1.4–12.0 |
| 7 | β-Pinene | 974 | 967 | 0.1 | 0–1.1 |
| 8 | Myrcene | 983 | 979 | 2.3 | t–4.1 |
| 9 | α-Phellenadrene | 998 | 993 | 0.1 | 0–0.1 |
| 10 | Car-3-ene | 1006 | 1001 | 0.3 | 0–1.0 |
| 11 | α-Terpinene | 1013 | 1006 | 1.7 | t–1.8 |
| 12 | p-Cymene | 1016 | 1008 | 0.6 | 0.3–2.4 |
| 13 | β-Phellandrene | 1025 | 1008 | 0.3 | 0–1.7 |
| 14 | Limonene | 1025 | 1018 | 1.3 | 0.3–3.2 |
| 15 | γ-Terpinene | 1051 | 1048 | 2.7 | 0.2–2.3 |
| 16 | trans-Sabinene hydrate | 1053 | 1053 | 0.1 | 0–1.1 |
| 17 | Fenchone | 1069 | 1068 | 9.3 | 0.4–15.0 |
| 18 | p-Cymenene | 1075 | 1068 | t | |
| 19 | Terpinolene | 1081 | 1078 | 0.7 | 0.2–0.3 |
| 20 | α-Thujone | 1089 | 1090 | 38.5 | 2.8–69.8 |
| 21 | β-Thujone | 1103 | 1098 | 4.9 | 3.1–10.7 |
| 22 | cis-p-Menth-2-en-1-ol | 1108 | 1107 | 0.5 | 0–0.6 |
| 23 | Camphor | 1123 | 1119 | 0.1 | 0–3.3 |
| 24 | Neoisothujol | 1132 | 1123 | 0.3 | 0.2 |
| 25 | Sabina ketone | 1132 | 1126 | 0.1 | |
| 26 | Camphene hydrate | 1143 | 1131 | 0.2 | 0.2 |
| 27 | Neothujol | 1136 | 1134 | 0.1 | |
| 28 | Borneol | 1150 | 1149 | 0.3 | 0–0.7 |
| 29 | Terpinen-4-ol | 1164 | 1163 | 7.3 | 1.2–3.3 |
| 30 | α-Terpineol | 1176 | 1172 | 0.4 | 0.1–0.9 |
| 31 | Estragol | 1175 | 1176 | 0.1 | |
| 32 | cis-Piperitol | 1181 | 1180 | 0.1 | |
| 33 | trans-Piperitol | 1193 | 1190 | 0.1 | |
| 34 | Fenchyl acetate | 1205 | 1207 | 2.7 | 0–1.1 |
| 35 | Piperitone | 1226 | 1226 | t | |
| 36 | Linalyl acetate | 1239 | 1240 | t | 0–1.2 |
| 37 | Neoisothujan-3-ol acetate | 1256 | 0.1 | ||
| 38 | Bornyl acetate | 1270 | 1270 | 4.3 | 0.1–3.9 |
| 39 | trans-Sabinyl acetate | 1278 | 1272 | 0.1 | 0–16.6 |
| 40 | Terpinen-4-ol acetate | 1289 | 1283 | 0.1 | |
| 41 | α-Terpinyl acetate | 1335 | 1332 | 1.0 | 0–1.8 |
| 42 | δ-Cadinene | 1520 | 1515 | t | 0–1.3 |
| 43 | Beyerene | 1936 | 0.5 | 0–13.2 | |
| Total identified | 99.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gospodarek, J.; Krajewska, A.; Paśmionka, I.B.; Bruździńska, J.; Tamiru, G. Potential of Thuja occidentalis L. Essential Oil and Water Extracts against Field Crop Pests. Molecules 2024, 29, 1457. https://doi.org/10.3390/molecules29071457
Gospodarek J, Krajewska A, Paśmionka IB, Bruździńska J, Tamiru G. Potential of Thuja occidentalis L. Essential Oil and Water Extracts against Field Crop Pests. Molecules. 2024; 29(7):1457. https://doi.org/10.3390/molecules29071457
Chicago/Turabian StyleGospodarek, Janina, Agnieszka Krajewska, Iwona B. Paśmionka, Joanna Bruździńska, and Gedyon Tamiru. 2024. "Potential of Thuja occidentalis L. Essential Oil and Water Extracts against Field Crop Pests" Molecules 29, no. 7: 1457. https://doi.org/10.3390/molecules29071457
APA StyleGospodarek, J., Krajewska, A., Paśmionka, I. B., Bruździńska, J., & Tamiru, G. (2024). Potential of Thuja occidentalis L. Essential Oil and Water Extracts against Field Crop Pests. Molecules, 29(7), 1457. https://doi.org/10.3390/molecules29071457