
Oxymatrine Modulation of TLR3 Signaling: A Dual-Action Mechanism for H9N2 Avian Influenza Virus Defense and Immune Regulation


Abstract
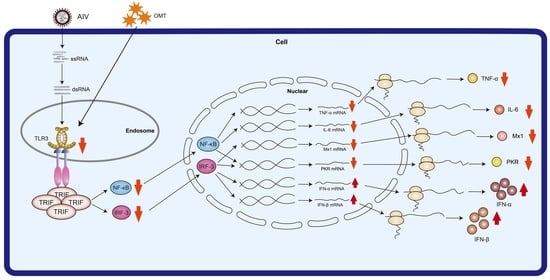
:
1. Introduction
2. Results
2.1. Observation and Immunofluorescent Identification of PMVECs
2.2. H9N2 AIV Infectivity in PMVECs Measured Using TCID50
2.3. Cytotoxic Effects of OMT on PMVECs Assessed with the CCK-8 Assay
2.4. The Influence of OMT on Antiviral Protein Expression in H9N2 AIV-Infected PMVECs
2.5. Regulatory Effects of OMT on the Expression of IFN-α, IFN-β, IL-6, and TNF-α in PMVECs Infected with H9N2 AIV
2.6. Effects of OMT on mRNA and Protein Expression Levels of NF-κB, IRF-3, and TLR3 in H9N2 AIV-Infected PMVECs
2.7. Effects of OMT on mRNA and Protein Expression of NF-κB and IRF-3 Post-TLR3 Silencing in H9N2 AIV-Infected PMVECs
3. Discussion
4. Materials and Methods
4.1. Cell Lines and Viruses
4.2. Immunofluorescence Characterization of PMVECs
4.3. siRNA-Mediated TLR3 Silencing in PMVECs
4.4. OMT Characterization and Cytotoxicity Assessment
4.5. Determination of H9N2 TCID50
4.6. Experimental Grouping and Treatment Methods
4.7. ELISA Determination of Cytokine Levels
4.8. RNA Extraction and Reverse-Transcription Quantitative PCR
4.9. Western Blot Analysis of Protein Expression in PMVECs
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Starick, E.; Römer-Oberdörfer, A.; Werner, O. Type- and subtype-specific RT-PCR assays for avian influenza A viruses (AIV). J. Vet. Med. B Infect. Dis. Vet. Public Health 2000, 47, 295–301. [Google Scholar] [CrossRef] [PubMed]
- Wolff, T.; Veit, M. Influenza B, C and D Viruses (Orthomyxoviridae). Encycl. Virol. 2021, 2021, 561–574. [Google Scholar] [CrossRef]
- Nogales, A.; Aydillo, T.; Ávila-Pérez, G.; Escalera, A.; Chiem, K.; Cadagan, R.; DeDiego, M.L.; Li, F.; García-Sastre, A.; Martínez-Sobrido, L. Functional Characterization and Direct Comparison of Influenza A, B, C, and D NS1 Proteins in vitro and in vivo. Front. Microbiol. 2019, 10, 2862. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.F.; Lai, H.Z.; Li, L.; Liu, Y.P.; Zhang, W.Y.; Gao, R.; Huang, W.K.; Luo, Q.F.; Gao, Y.; Luo, Q.; et al. Endemic Variation of H9N2 Avian Influenza Virus in China. Avian Dis. 2016, 60, 817–825. [Google Scholar] [CrossRef] [PubMed]
- Wei, K.; Li, Y. Global genetic variation and transmission dynamics of H9N2 avian influenza virus. Transbound. Emerg. Dis. 2018, 65, 504–517. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Zhu, W.; Yang, L.; Shu, Y. The Epidemiology, Virology, and Pathogenicity of Human Infections with Avian Influenza Viruses. Cold Spring Harb. Perspect. Med. 2021, 11, a038620. [Google Scholar] [CrossRef]
- Zhang, J.; Ye, H.; Liu, Y.; Liao, M.; Qi, W. Resurgence of H5N6 avian influenza virus in 2021 poses new threat to public health. Lancet Microbe 2022, 3, e558. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Quan, K.; Chen, Z.; Hu, Q.; Nie, M.; Xu, N.; Gao, R.; Wang, X.; Qin, T.; Chen, S.; et al. The emergence of new antigen branches of H9N2 avian influenza virus in China due to antigenic drift on hemagglutinin through antibody escape at immunodominant sites. Emerg. Microbes Infect. 2023, 12, 2246582. [Google Scholar] [CrossRef] [PubMed]
- de Vries, R.D.; Herfst, S.; Richard, M. Avian Influenza A Virus Pandemic Preparedness and Vaccine Development. Vaccines 2018, 6, 46. [Google Scholar] [CrossRef]
- Huo, C.; Jin, Y.; Zou, S.; Qi, P.; Xiao, J.; Tian, H.; Wang, M.; Hu, Y. Lethal influenza A virus preferentially activates TLR3 and triggers a severe inflammatory response. Virus Res. 2018, 257, 102–112. [Google Scholar] [CrossRef]
- Nang, N.T.; Lee, J.S.; Song, B.M.; Kang, Y.M.; Kim, H.S.; Seo, S.H. Induction of inflammatory cytokines and Toll-like receptors in chickens infected with avian H9N2 influenza virus. Vet. Res. 2011, 42, 64. [Google Scholar] [CrossRef]
- Teijaro, J.R.; Walsh, K.B.; Cahalan, S.; Fremgen, D.M.; Roberts, E.; Scott, F.; Martinborough, E.; Peach, R.; Oldstone, M.B.; Rosen, H. Endothelial cells are central orchestrators of cytokine amplification during influenza virus infection. Cell 2011, 146, 980–991. [Google Scholar] [CrossRef]
- Sun, W.; Wu, W.; Jiang, N.; Ge, X.; Zhang, Y.; Han, J.; Guo, X.; Zhou, L.; Yang, H. Highly Pathogenic PRRSV-Infected Alveolar Macrophages Impair the Function of Pulmonary Microvascular Endothelial Cells. Viruses 2022, 14, 452. [Google Scholar] [CrossRef]
- Edelmann, K.H.; Richardson-Burns, S.; Alexopoulou, L.; Tyler, K.L.; Flavell, R.A.; Oldstone, M.B. Does Toll-like receptor 3 play a biological role in virus infections? Virology 2004, 322, 231–238. [Google Scholar] [CrossRef]
- Nasirudeen, A.M.; Wong, H.H.; Thien, P.; Xu, S.; Lam, K.P.; Liu, D.X. RIG-I, MDA5 and TLR3 synergistically play an important role in restriction of dengue virus infection. PLoS Negl. Trop. Dis. 2011, 5, e926. [Google Scholar] [CrossRef]
- Bi, W.; Tian, M.; Row, K.H. Selective extraction and separation of Oxymatrine from Sophora flavescens Ait. extract by silica-confined ionic liquid. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2012, 880, 108–113. [Google Scholar] [CrossRef]
- Liang, L.; Sun, W.; Wei, X.; Wang, L.; Ruan, H.; Zhang, J.; Li, S.; Zhao, B.; Li, M.; Cai, Z.; et al. Oxymatrine suppresses colorectal cancer progression by inhibiting NLRP3 inflammasome activation through mitophagy induction in vitro and in vivo. Phytother. Res. 2023, 37, 3342–3362. [Google Scholar] [CrossRef]
- Lu, L.G.; Zeng, M.D.; Mao, Y.M.; Fang, J.Y.; Song, Y.L.; Shen, Z.H.; Cao, A.P. Inhibitory effect of Oxymatrine on serum hepatitis B virus DNA in HBV transgenic mice. World J. Gastroenterol. 2004, 10, 1176–1179. [Google Scholar] [CrossRef]
- Li, S.; Zhang, H.; Jiao, Y.; Song, X.; Wei, L.; Liu, X. Oxymatrine induces anti-tumor response in cervical cancer by modulating circ_0008460/miR-197-3p/ribonucleotide reductase subunit M2 (RRM2). Bioengineered 2022, 13, 12912–12926. [Google Scholar] [CrossRef]
- Cao, G.; Mao, Z.; Niu, T.; Wang, P.; Yue, X.; Wang, X.; Ma, A.; Zhang, Y. Oxymatrine ameliorates rheumatoid arthritis by regulation of Tfr/Tfh cell balance via the TLR9-MyD88-STAT3 signaling pathway. J. Sci. Food Agric. 2023, 103, 6017–6024. [Google Scholar] [CrossRef]
- Zhang, F.L.; Yang, L.; He, W.H.; Xie, L.J.; Yang, F.; Wang, Y.H.; Huang, A.G. In vivo antibacterial activity of medicinal plant Sophora flavescens against Streptococcus agalactiae infection. J. Fish Dis. 2023, 46, 977–986. [Google Scholar] [CrossRef]
- Chen, X.S.; Wang, G.J.; Cai, X.; Yu, H.Y.; Hu, Y.P. Inhibition of hepatitis B virus by Oxymatrine in vivo. World J. Gastroenterol. 2001, 7, 49–52. [Google Scholar] [CrossRef]
- Dai, J.P.; Wang, Q.W.; Su, Y.; Gu, L.M.; Deng, H.X.; Chen, X.X.; Li, W.Z.; Li, K.S. Oxymatrine Inhibits Influenza A Virus Replication and Inflammation via TLR4, p38 MAPK and NF-κB Pathways. Int. J. Mol. Sci. 2018, 19, 965. [Google Scholar] [CrossRef] [PubMed]
- Dong, J.; Zhou, Y.; Pu, J.; Liu, L. Status and Challenges for Vaccination against Avian H9N2 Influenza Virus in China. Life 2022, 12, 1326. [Google Scholar] [CrossRef] [PubMed]
- Zuo, W.; Wakimoto, M.; Kozaiwa, N.; Shirasaka, Y.; Oh, S.W.; Fujiwara, S.; Miyachi, H.; Kogure, A.; Kato, H.; Fujita, T. PKR and TLR3 trigger distinct signals that coordinate the induction of antiviral apoptosis. Cell Death Dis. 2022, 13, 707. [Google Scholar] [CrossRef] [PubMed]
- Pillai, P.S.; Molony, R.D.; Martinod, K.; Dong, H.; Pang, I.K.; Tal, M.C.; Solis, A.G.; Bielecki, P.; Mohanty, S.; Trentalange, M.; et al. Mx1 reveals innate pathways to antiviral resistance and lethal influenza disease. Science 2016, 352, 463–466. [Google Scholar] [CrossRef] [PubMed]
- Sang, X.; Wang, R.; Han, Y.; Zhang, C.; Shen, H.; Yang, Z.; Xiong, Y.; Liu, H.; Liu, S.; Li, R.; et al. T cell—Associated immunoregulation and antiviral effect of Oxymatrine in hydrodynamic injection HBV mouse model. Acta Pharm. Sin. B 2017, 7, 311–318. [Google Scholar] [CrossRef]
- Lee, B.W.; Ha, T.K.Q.; Cho, H.M.; An, J.P.; Kim, S.K.; Kim, C.S.; Kim, E.; Oh, W.K. Antiviral activity of furanocoumarins isolated from Angelica dahurica against influenza a viruses H1N1 and H9N2. J. Ethnopharmacol. 2020, 259, 112945. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, R.; Shi, W.; Zheng, Z.; Wang, X.; Li, C.; Zhang, S.; Zhang, P. Antiviral effect of fufang yinhua jiedu (FFYH) granules against influenza A virus through regulating the inflammatory responses by TLR7/MyD88 signaling pathway. J. Ethnopharmacol. 2021, 275, 114063. [Google Scholar] [CrossRef]
- van de Sand, L.; Bormann, M.; Schmitz, Y.; Heilingloh, C.S.; Witzke, O.; Krawczyk, A. Antiviral Active Compounds Derived from Natural Sources against Herpes Simplex Viruses. Viruses 2021, 13, 1386. [Google Scholar] [CrossRef]
- Audry, M.; Ciancanelli, M.; Yang, K.; Cobat, A.; Chang, H.H.; Sancho-Shimizu, V.; Lorenzo, L.; Niehues, T.; Reichenbach, J.; Li, X.X.; et al. NEMO is a key component of NF-κB- and IRF-3-dependent TLR3-mediated immunity to herpes simplex virus. J. Allergy Clin. Immunol. 2011, 128, 610–617. [Google Scholar] [CrossRef] [PubMed]
- de Carvalho, R.V.H.; Lima-Junior, D.S.; da Silva, M.V.G.; Dilucca, M.; Rodrigues, T.S.; Horta, C.V.; Silva, A.L.N.; da Silva, P.F.; Frantz, F.G.; Lorenzon, L.B.; et al. Leishmania RNA virus exacerbates Leishmaniasis by subverting innate immunity via TLR3-mediated NLRP3 inflammasome inhibition. Nat. Commun. 2019, 10, 5273. [Google Scholar] [CrossRef] [PubMed]
- Xu, R.H.; Wong, E.B.; Rubio, D.; Roscoe, F.; Ma, X.; Nair, S.; Remakus, S.; Schwendener, R.; John, S.; Shlomchik, M.; et al. Sequential Activation of Two Pathogen-Sensing Pathways Required for Type I Interferon Expression and Resistance to an Acute DNA Virus Infection. Immunity 2015, 43, 1148–1159. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Zhao, F.; Xu, J.; Zhu, X.; Zhao, Y.; Wen, R.; Anirudhan, V.; Rong, L.; Tian, J.; Cui, Q. Qingjin Huatan decoction protects mice against influenza a virus pneumonia via the chemokine signaling pathways. J. Ethnopharmacol. 2023, 317, 116745. [Google Scholar] [CrossRef] [PubMed]
- Govorkova, E.A.; Fang, H.B.; Tan, M.; Webster, R.G. Neuraminidase inhibitor-rimantadine combinations exert additive and synergistic anti-influenza virus effects in MDCK cells. Antimicrob. Agents Chemother. 2004, 48, 4855–4863. [Google Scholar] [CrossRef] [PubMed]
- Ocaña-Macchi, M.; Bel, M.; Guzylack-Piriou, L.; Ruggli, N.; Liniger, M.; McCullough, K.C.; Sakoda, Y.; Isoda, N.; Matrosovich, M.; Summerfield, A. Hemagglutinin-dependent tropism of H5N1 avian influenza virus for human endothelial cells. J. Virol. 2009, 83, 12947–12955. [Google Scholar] [CrossRef]
- Mostafa, A.; Mahmoud, S.H.; Shehata, M.; Müller, C.; Kandeil, A.; El-Shesheny, R.; Nooh, H.Z.; Kayali, G.; Ali, M.A.; Pleschka, S. PA from a Recent H9N2 (G1-Like) Avian Influenza a Virus (AIV) Strain Carrying Lysine 367 Confers Altered Replication Efficiency and Pathogenicity to Contemporaneous H5N1 in Mammalian Systems. Viruses 2020, 12, 1046. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Liang, T.; Luo, Q.; Li, P.; Zhang, R.; Xu, M.; Su, J.; Xu, T.; Wu, Q. H9N2 swine influenza virus infection-induced damage is mediated by TRPM2 channels in mouse pulmonary microvascular endothelial cells. Microb. Pathog. 2020, 148, 104408. [Google Scholar] [CrossRef] [PubMed]
- Kovacs-Kàsa, A.; Varn, M.N.; Verin, A.D.; Gonzales, J.N. Method for the Culture of Mouse Pulmonary Microvascular Endothelial Cells. Sci. Pages Pulmonol. 2017, 1, 7–18. [Google Scholar]
- Kleinman, M.E.; Yamada, K.; Takeda, A.; Chandrasekaran, V.; Nozaki, M.; Baffi, J.Z.; Albuquerque, R.J.; Yamasaki, S.; Itaya, M.; Pan, Y.; et al. Sequence- and target-independent angiogenesis suppression by siRNA via TLR3. Nature 2008, 452, 591–597. [Google Scholar] [CrossRef]
- Ramakrishnan, M.A. Determination of 50% endpoint titer using a simple formula. World J. Virol. 2016, 5, 85–86. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.; Chen, T.; Hao, Z.; Yang, X.; Wang, M.; Zhang, Z.; Hao, S.; Lang, F.; Hao, H. Oxymatrine Alleviates Gentamicin-Induced Renal Injury in Rats. Molecules 2022, 27, 6209. [Google Scholar] [CrossRef] [PubMed]
- Meng, S.; Xia, F.; Xu, J.; Zhang, X.; Xue, M.; Gu, M.; Guo, F.; Huang, Y.; Qiu, H.; Yang, Y. Hepatocyte growth factor protects pulmonary endothelial barrier against oxidative stress and mitochondria-dependent apoptosis. Chin. Med. J. 2022, 135, 837–848. [Google Scholar] [CrossRef]
- Xue, X.; Guo, Y.; Zhao, Q.; Li, Y.; Rao, M.; Qi, W.; Shi, H. Weighted Gene Co-Expression Network Analysis of Oxymatrine in Psoriasis Treatment. J. Inflamm. Res. 2023, 16, 845–859. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
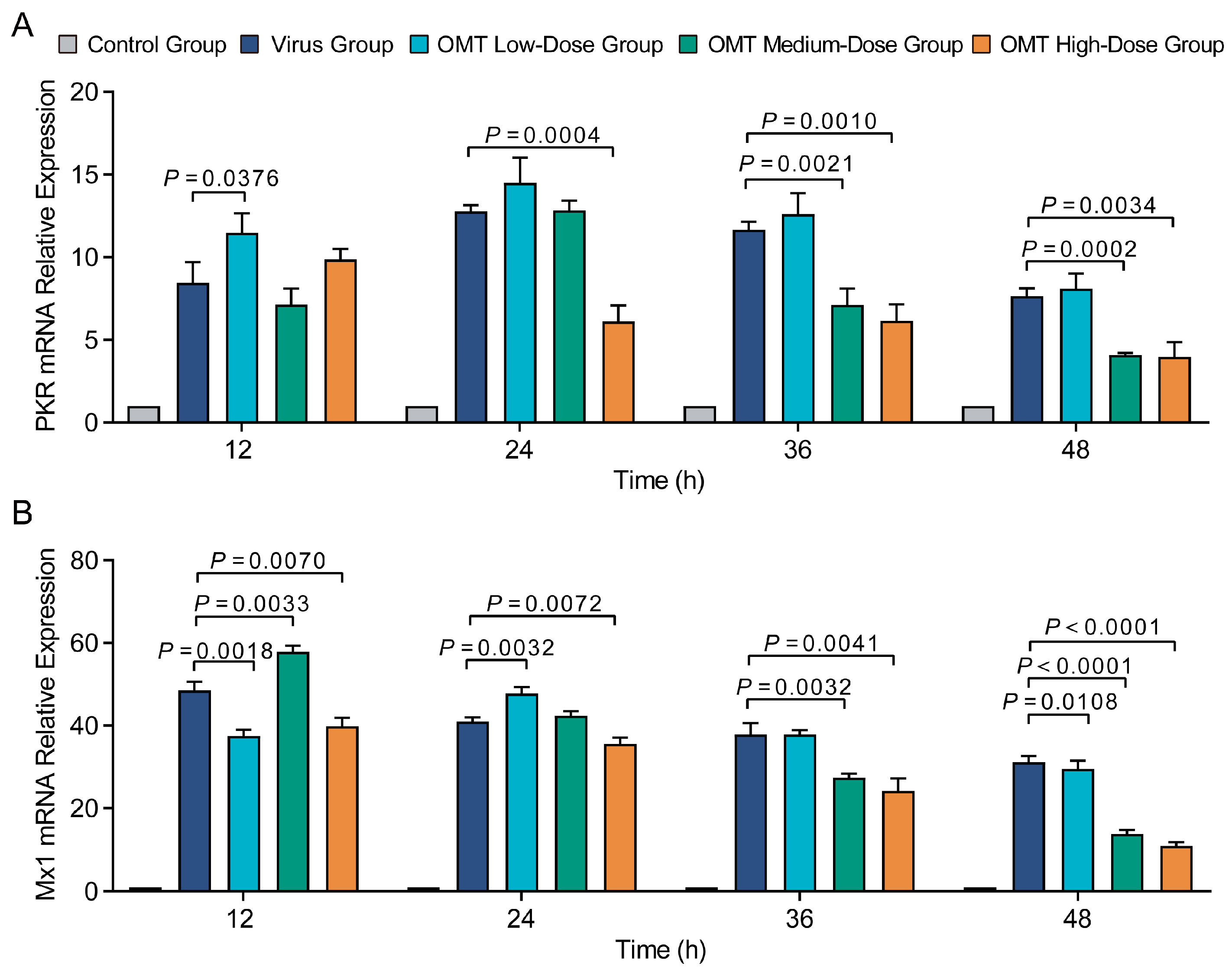{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Virus Dilution | Observed CPE | Cumulative CPE | Infection Rate (%) | ||||
|---|---|---|---|---|---|---|---|
| Well Count | CPE Positive Wells | CPE Negative Wells | Cumulative CPE Positive Wells | Cumulative CPE Negative Wells | Proportion of CPE Positive Wells | CPE Positive Rate (%) | |
| 10−1 | 8 | 8 | 0 | 33 | 0 | 33/33 | 100 |
| 10−2 | 8 | 8 | 0 | 25 | 0 | 25/25 | 100 |
| 10−3 | 8 | 8 | 0 | 17 | 0 | 17/17 | 100 |
| 10−4 | 8 | 6 | 2 | 11 | 2 | 11/13 | 84.61 |
| 10−5 | 8 | 3 | 5 | 5 | 7 | 5/12 | 41.67 |
| 10−6 | 8 | 2 | 6 | 2 | 13 | 2/15 | 13.33 |
| 10−7 | 8 | 0 | 8 | 0 | 21 | 0/21 | 0 |
| OMT Concentration (μg/mL) | 0 | 3.125 | 6.25 | 12.5 | 25 | 50 | 100 | 200 | ||
|---|---|---|---|---|---|---|---|---|---|---|
| Survival Rate (%) | ||||||||||
| Time (h) | ||||||||||
| 12 | 100.00 | 95.96 | 95.03 | 92.72 | 92.52 | 88.77 | 68.06 | 70.39 | ||
| 24 | 100.00 | 95.36 | 96.23 | 90.67 | 79.12 | 72.94 | 57.66 | 36.79 | ||
| 36 | 100.00 | 95.39 | 93.52 | 84.81 | 74.71 | 62.07 | 47.47 | 29.69 | ||
| 48 | 100.00 | 95.98 | 95.06 | 84.34 | 73.61 | 63.61 | 27.38 | 27.04 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhi, Y.; Zhao, X.; Liu, Z.; Shen, G.; Zhang, T.; Zhang, T.; Hu, G. Oxymatrine Modulation of TLR3 Signaling: A Dual-Action Mechanism for H9N2 Avian Influenza Virus Defense and Immune Regulation. Molecules 2024, 29, 1945. https://doi.org/10.3390/molecules29091945
Zhi Y, Zhao X, Liu Z, Shen G, Zhang T, Zhang T, Hu G. Oxymatrine Modulation of TLR3 Signaling: A Dual-Action Mechanism for H9N2 Avian Influenza Virus Defense and Immune Regulation. Molecules. 2024; 29(9):1945. https://doi.org/10.3390/molecules29091945
Chicago/Turabian StyleZhi, Yan, Xinping Zhao, Zhenyi Liu, Guoyu Shen, Taiming Zhang, Tao Zhang, and Ge Hu. 2024. "Oxymatrine Modulation of TLR3 Signaling: A Dual-Action Mechanism for H9N2 Avian Influenza Virus Defense and Immune Regulation" Molecules 29, no. 9: 1945. https://doi.org/10.3390/molecules29091945
APA StyleZhi, Y., Zhao, X., Liu, Z., Shen, G., Zhang, T., Zhang, T., & Hu, G. (2024). Oxymatrine Modulation of TLR3 Signaling: A Dual-Action Mechanism for H9N2 Avian Influenza Virus Defense and Immune Regulation. Molecules, 29(9), 1945. https://doi.org/10.3390/molecules29091945