Glycosylation of Fluorophenols by Plant Cell Cultures

Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results and Discussion
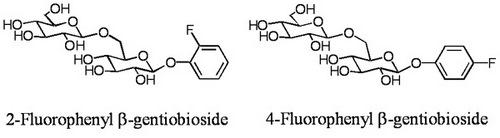
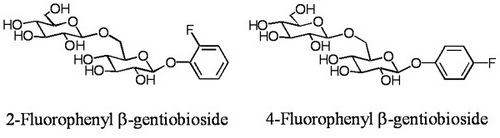
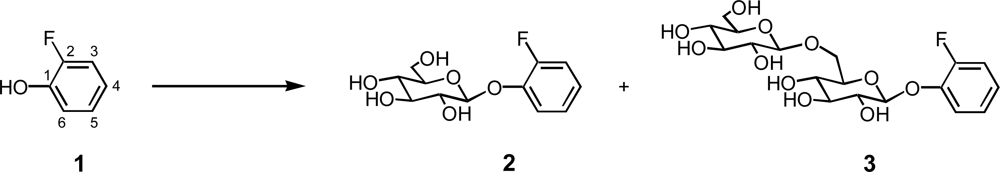
2.1. Glycosylation of 2-fluorophenol (1)
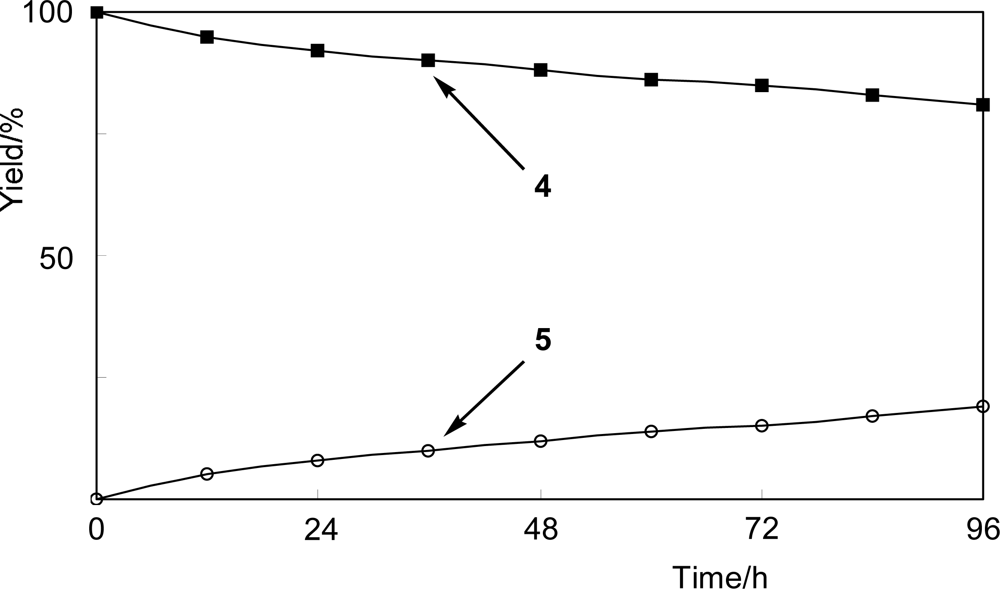
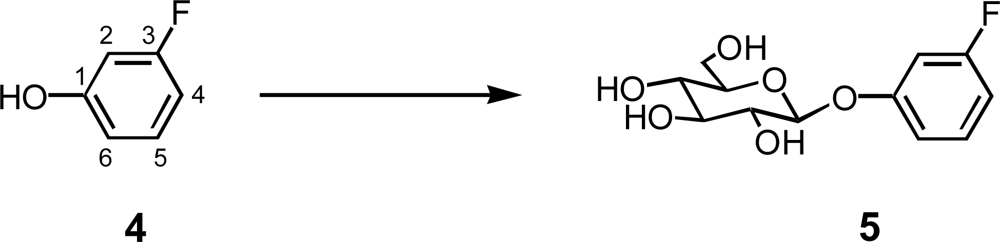
2.2. Glycosylation of 3-fluorophenol (4)
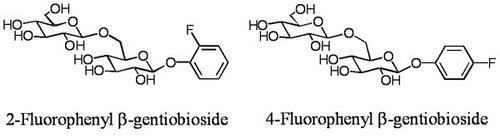
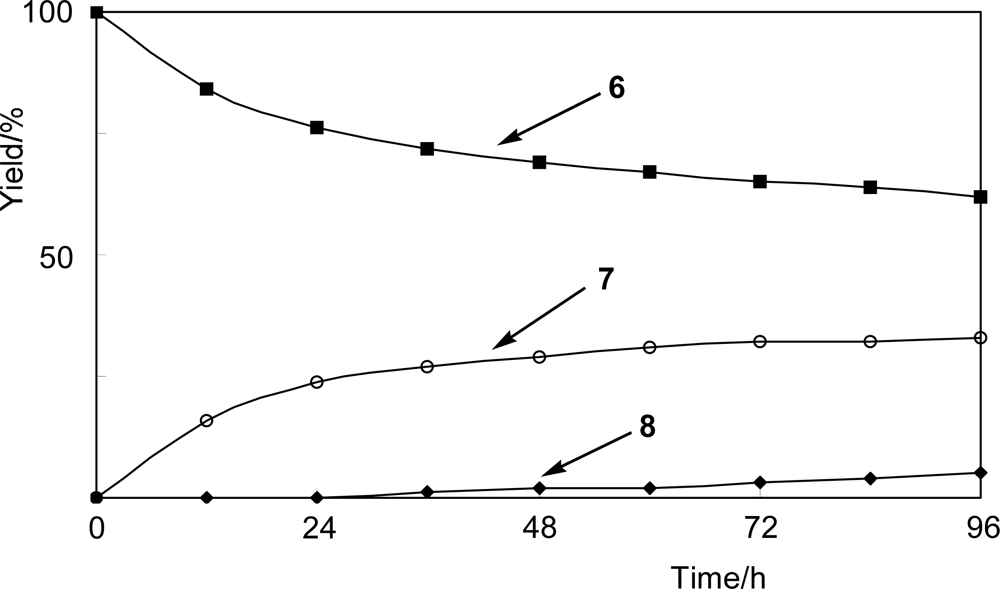
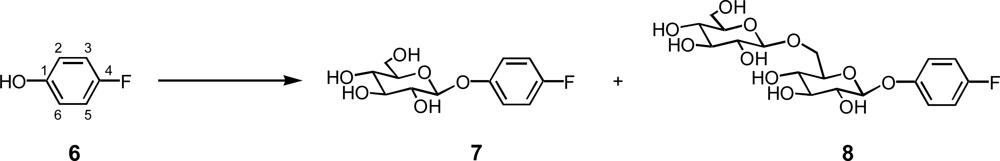
2.3. Glycosylation of 4-fluorophenol (6)
3. Conclusions
4. Experimental Section
4.1. General
4.2. Biotransformation of 1, 4, and 6 with plant cell cultures of N. tabacum
2-Fluorophenyl β-gentiobioside (3):
4-Fluorophenyl β-gentiobioside (8):
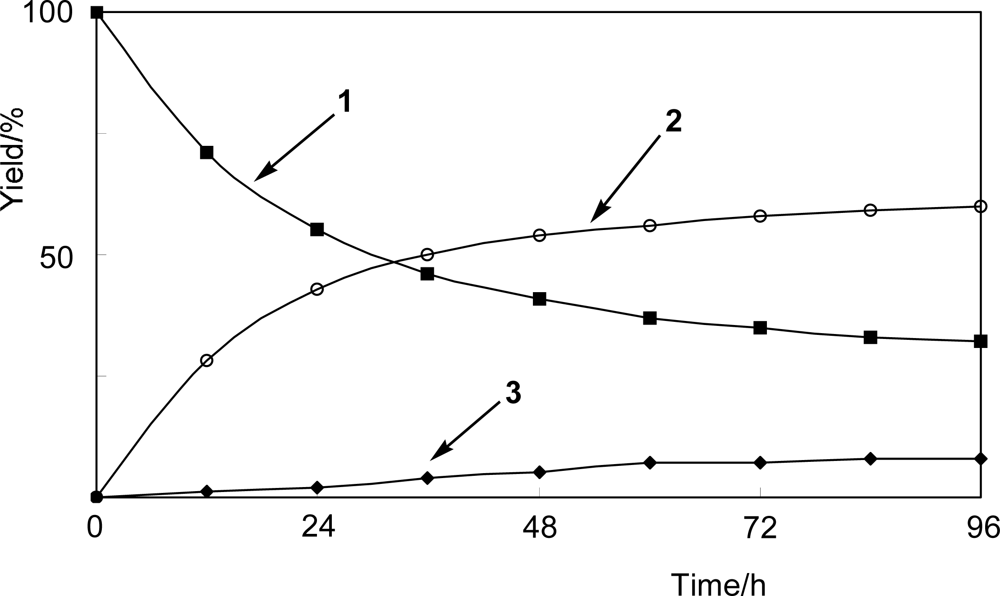
4.3. Time-course experiment
References
- Key, BD; Howell, RD; Criddle, CS. Fluorinated organics in the biosphere. Environ. Sci. Technol 1997, 31, 2445–2454. [Google Scholar]
- Haggblom, MM. Microbial breakdown of halogenated aromatic pesticides and related compounds. FEMS Microbiol. Rev 1992, 9, 29–71. [Google Scholar]
- Kiyohara, H; Hatta, T; Ogawa, Y; Kakuda, T; Yokoyama, H; Takizawa, N. Isolation of Pseudomonas pickettii strains that degrade 2,4,6-trichlorophenol and their dechlorination of chlorophenols. Appl. Environ. Microbiol 1992, 58, 1276–1283. [Google Scholar]
- Hofrichter, M; Bublitz, F; Fritsche, W. Unspecific degradation of halogenated phenols by the soil fungus Penicillium frequentans Bi 7/2. J. Basic. Microbiol 1994, 34, 163–172. [Google Scholar]
- Bae, HS; Lee, JM; Lee, ST. Biodegradation of 4-chlorophenol via a hydroquinone pathway by Arthrobacter ureafaciens CPR706. FEMS Microbiol. Lett 1996, 145, 125–129. [Google Scholar]
- Hollender, J; Hopp, J; Dott, W. Degradation of 4-chlorophenol via the meta cleavage pathway by Comamonas testosteroni JH5. Appl. Environ. Microbiol 1997, 63, 4567–4572. [Google Scholar]
- Boersma, MG; Dinarieva, TY; Middelhoven, WJ; van Berkel, WJ; Doran, J; Vervoort, J; Rietjens, IM. 19F nuclear magnetic resonance as a tool to investigate microbial degradation of fluorophenols to fluorocatechols and fluoromuconates. Appl. Environ. Microbiol 1998, 64, 1256–1263. [Google Scholar]
- Bondar, VS; Boersma, MG; van Berkel, WJ; Finkelstein, ZI; Golovlev, EL; Baskunov, BP; Vervoort, J; Golovleva, LA; Rietjens, IM. Preferential oxidative dehalogenation upon conversion of 2-halophenols by Rhodococcus opacus 1G. FEMS Microbiol. Lett 1999, 181, 73–82. [Google Scholar]
- Boersma, MG; Solyanikova, IP; van Berkel, WJ; Vervoort, J; Golovleva, LA; Rietjens, IM. 19F NMR metabolomics for the elucidation of microbial degradation pathways of fluorophenols. J. Ind. Microbiol. Biotechnol 2001, 26, 22–34. [Google Scholar]
- Ishihara, K; Hamada, H; Hirata, T; Nakajima, N. Biotransformation using plant cultured cells. J. Mol. Catal. B: Enz 2003, 23, 145–170. [Google Scholar]
- Furuya, T; Ushiyama, M; Ashida, Y; Yoshikawa, T. Glycosylation of 2-phenylpropionic acid and its ethyl ester in suspension cultures of Nicotiana tabacum, Dioscoreophyllum cumminsii and Aconitum japonicum. Phytochemistry 1987, 26, 2983–2989. [Google Scholar]
- Furuya, T; Ushiyama, M; Ashida, Y; Yoshikawa, T; Orihara, Y. Biotransformation of phenylacetic acid and 2-phenyl-propionic acid in suspension culture of Coffea arabica. Phytochemistry 1988, 27, 803–807. [Google Scholar]
- Ushiyama, M; Furuya, T. Glycosylation of phenolic compounds by root culture of Panax ginseng. Phytochemistry 1989, 28, 3009–3013. [Google Scholar]
- Kamel, S; Brazier, M; Desmet, G; Fliniaux, M-A; Jacquin-Dubreuil, A. Glucosylation of butyric acid by cell suspension culture of Nicotiana plumbaginifolia. Phytochemistry 1992, 31, 1581–1583. [Google Scholar]
- Lewinson, E; Berman, E; Mazur, Y; Gressel, J. Glucosylation of exogenous flavanones by grapefruit (Citrus paradisi) cell cultures. Phytochemistry 1996, 25, 2531–2535. [Google Scholar]
- Kondo, Y; Shimoda, K; Kubota, N; Hamada, H; Hamada, H. Biotransformation of monofluorophenols by cultured cells of Eucalyptus perriniana. Plant Biotechnology 2006, 23, 329–331. [Google Scholar]
- Shimoda, K; Kondo, Y; Akagi, M; Abe, K; Hamada, H; Hamada, H. Synthesis of α-tocopheryl disaccharides as potential antiallergic agents. Chemistry Letters 2007, 36, 570–571. [Google Scholar]
- Shimoda, K; Kwon, S; Utsuki, A; Ohiwa, S; Katsuragi, H; Yonemoto, N; Hamada, H; Hamada, H. Glycosylation of capsaicin and 8-nordihydrocapsaicin by cultured cells of Catharanthus roseus. Phytochemistry 2007, 68, 1391–1396. [Google Scholar]
- Kaminaga, Y; Nagatsu, A; Akiyama, T; Sugimoto, N; Yamazaki, T; Maitani, T; Mizukami, H. Production of unnatural glycosides of curcumin with drastically enhanced water solubility by cell suspension cultures of Catharanthus roseus. FEBS Lett 2003, 555, 311–316. [Google Scholar]
- Shimoda, K; Hamada, H; Hamada, H. Phytoremediation of benzophenone and bisphenol A by glycosylation with immobilized plant cells. Environ. Health Insights 2009, 3, 19–25. [Google Scholar]






© 2009 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Shimoda, K.; Kubota, N.; Kondo, Y.; Sato, D.; Hamada, H. Glycosylation of Fluorophenols by Plant Cell Cultures. Int. J. Mol. Sci. 2009, 10, 1942-1949. https://doi.org/10.3390/ijms10051942
Shimoda K, Kubota N, Kondo Y, Sato D, Hamada H. Glycosylation of Fluorophenols by Plant Cell Cultures. International Journal of Molecular Sciences. 2009; 10(5):1942-1949. https://doi.org/10.3390/ijms10051942
Chicago/Turabian StyleShimoda, Kei, Naoji Kubota, Yoko Kondo, Daisuke Sato, and Hiroki Hamada. 2009. "Glycosylation of Fluorophenols by Plant Cell Cultures" International Journal of Molecular Sciences 10, no. 5: 1942-1949. https://doi.org/10.3390/ijms10051942
APA StyleShimoda, K., Kubota, N., Kondo, Y., Sato, D., & Hamada, H. (2009). Glycosylation of Fluorophenols by Plant Cell Cultures. International Journal of Molecular Sciences, 10(5), 1942-1949. https://doi.org/10.3390/ijms10051942