The Mitochondrial Protein Translocation Motor: Structural Conservation between the Human and Yeast Tim14/Pam18-Tim16/Pam16 co-Chaperones

Abstract
:1. Introduction
2. Results and Discussion
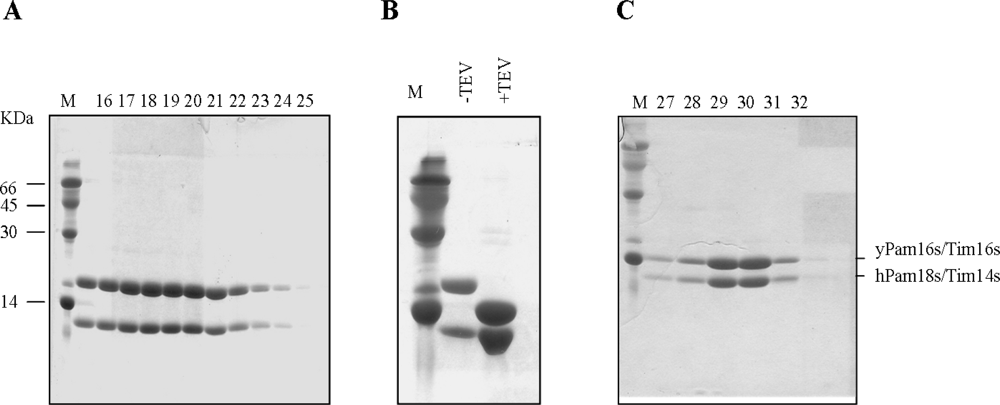
2.1. Purification of a Tim14/Pam18 - Tim16/Pam16 complex
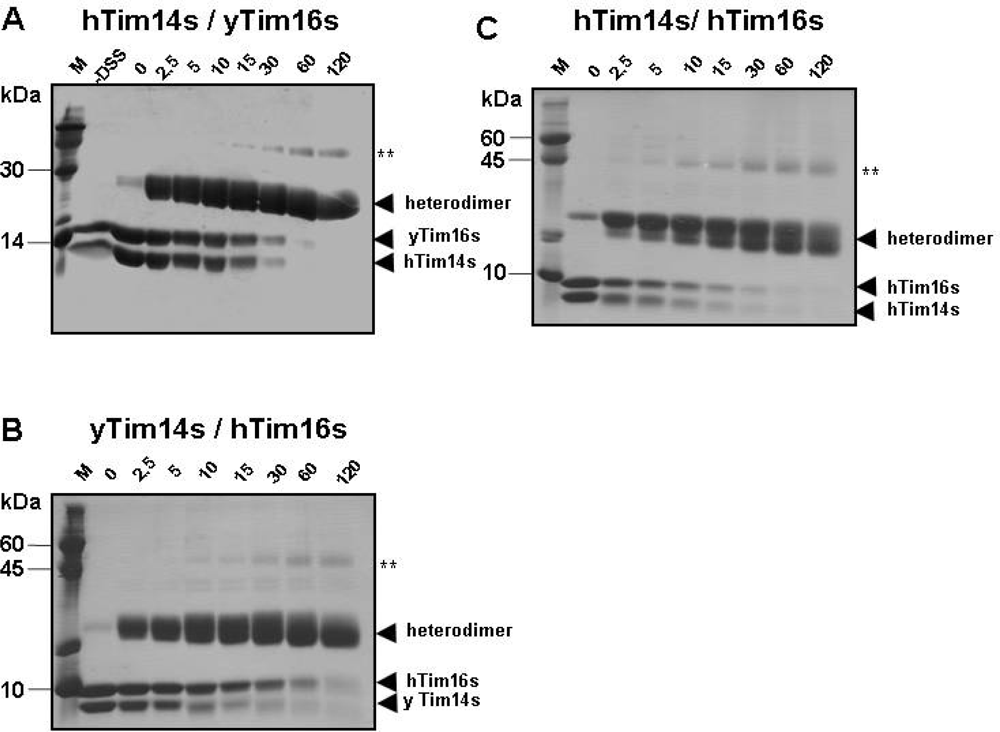
2.2. The oligomeric state of recombinant Tim14/Pam18s-Tim16/Tim16s complexes
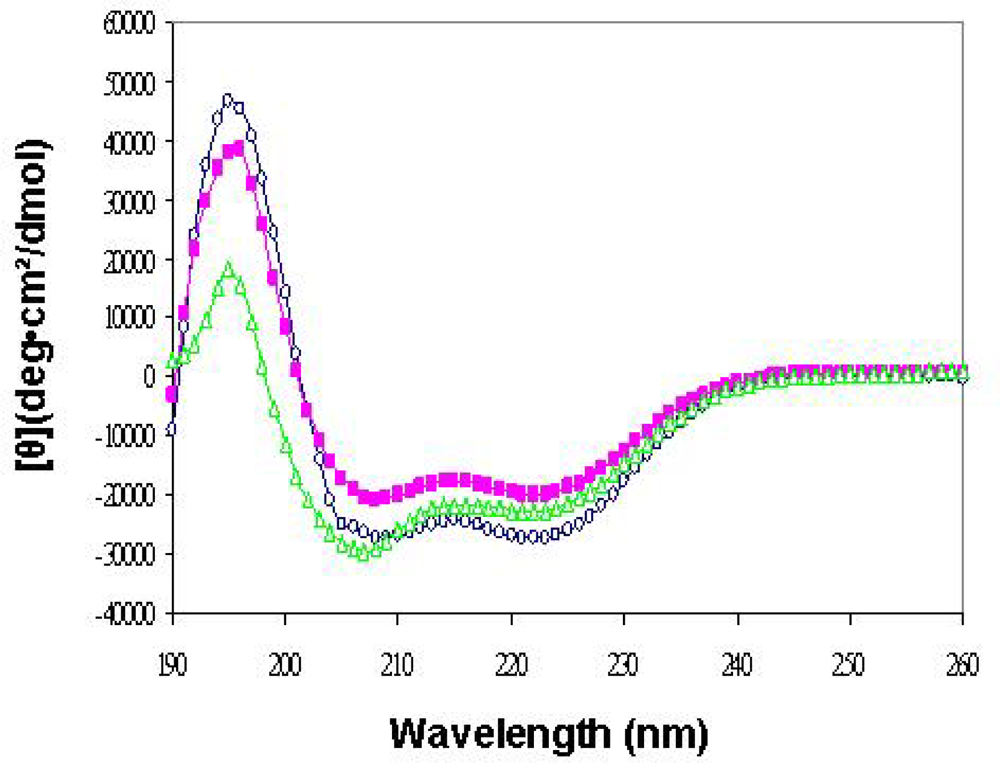
2.3. The folding and thermal stability of the purified complexes
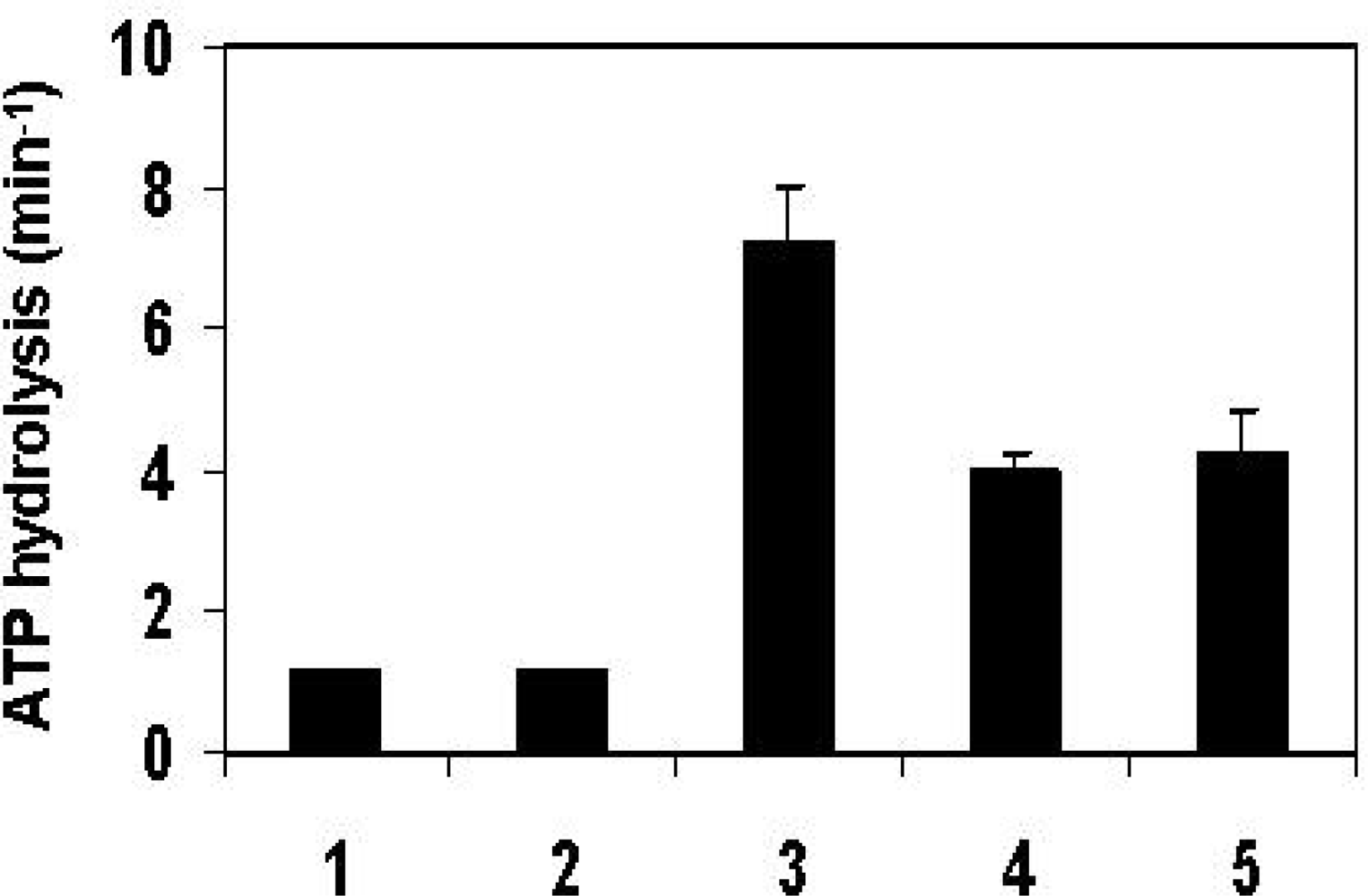
2.4. The effect of the y Tim14/Pam18-hTim16/Pam16 complex on the ATPase activity of yeast mtHsp70
3. Experimental Section
3.1. Cloning and Purification of the proteins used in this study
3.2. Circular dichroism (CD)
3.3. Cross-linking experiments
3.4. Miscellaneous
4. Conclusions
Acknowledgments
References and Notes
- Rapaport, D. How does the TOM complex mediate insertion of precursor proteins into the mitochondrial outer membrane? J. Cell Biol 2005, 171, 419–423. [Google Scholar]
- Mokranjac, D; Neupert, W. Protein import into mitochondria. Biochem. Soc. Trans 2005, 33, 1019–1023. [Google Scholar]
- Pfanner, N; Chacinska, A. The mitochondrial import machinery: preprotein-conducting channels with binding sites for presequences. Biochim. Biophys. Acta 2002, 1592, 15–24. [Google Scholar]
- Koehler, CM. New developments in mitochondrial assembly. Annu. Rev. Cell Dev. Biol 2004, 20, 309–335. [Google Scholar]
- Meinecke, M; Wagner, R; Kovermann, P; Guiard, B; Mick, DU; Hutu, DP; Voos, W; Truscott, KN; Chacinska, A; Pfanner, N; Rehling, P. Tim50 maintains the permeability barrier of the mitochondrial inner membrane. Science 2006, 312, 1523–1526. [Google Scholar]
- van der Laan, M; Rissler, M; Rehling, P. Mitochondrial preprotein translocases as dynamic molecular machines. FEMS Yeast Res 2006, 6, 849–861. [Google Scholar]
- Stojanovski, D; Rissler, M; Pfanner, N; Meisinger, C. Mitochondrial morphology and protein import-A tight connection? Biochim. Biophys. Acta 2006, 1763, 414–421. [Google Scholar]
- Matouschek, A; Pfanner, N; Voos, W. Protein unfolding by mitochondria. The Hsp70 import motor. EMBO Rep 2000, 1, 404–410. [Google Scholar]
- Horst, M; Azem, A; Schatz, G; Glick, BS. What is the driving force for protein import into mitochondria? Biochim. Biophys. Acta 1997, 1318, 71–78. [Google Scholar]
- Bomer, U; Meijer, M; Maarse, AC; Honlinger, A; Dekker, PJ; Pfanner, N; Rassow, J. Multiple interactions of components mediating preprotein translocation across the inner mitochondrial membrane. EMBO J 1997, 16, 2205–2216. [Google Scholar]
- Truscott, KN; Voos, W; Frazier, AE; Lind, M; Li, Y; Geissler, A; Dudek, J; Muller, H; Sickmann, A; Meyer, HE; Meisinger, C; Guiard, B; Rehling, P; Pfanner, N. A J-protein is an essential subunit of the presequence translocase-associated protein import motor of mitochondria. J. Cell Biol 2003, 163, 707–713. [Google Scholar]
- Moro, F; Sirrenberg, C; Schneider, HC; Neupert, W; Brunner, M. The TIM17.23 preprotein translocase of mitochondria: composition and function in protein transport into the matrix. EMBO J 1999, 18, 3667–3675. [Google Scholar]
- Slutsky-Leiderman, O; Marom, M; Iosefson, O; Levy, R; Maoz, S; Azem, A. The interplay between components of the mitochondrial protein translocation motor studied using purified components. J. Biol. Chem 2007, 282, 33935–33942. [Google Scholar]
- D’Silva, PD; Schilke, B; Walter, W; Andrew, A; Craig, EA. J protein cochaperone of the mitochondrial inner membrane required for protein import into the mitochondrial matrix. Proc. Nat. Acad. Sci. USA 2003, 100, 13839–13844. [Google Scholar]
- Mokranjac, D; Sichting, M; Neupert, W; Hell, K. Tim14, a novel key component of the import motor of the TIM23 protein translocase of mitochondria. EMBO J 2003, 22, 4945–4956. [Google Scholar]
- D’Silva, PR; Schilke, B; Walter, W; Craig, EA. Role of Pam16’s degenerate J domain in protein import across the mitochondrial inner membrane. Proc. Natl. Acad. Sci. USA 2005, 102, 12419–12424. [Google Scholar]
- Frazier, AE; Dudek, J; Guiard, B; Voos, W; Li, Y; Lind, M; Meisinger, C; Geissler, A; Sickmann, A; Meyer, HE; Bilanchone, V; Cumsky, MG; Truscott, KN; Pfanner, N; Rehling, P. Pam16 has an essential role in the mitochondrial protein import motor. Nat. Struct. Mol. Biol 2004, 11, 226–233. [Google Scholar]
- Kozany, C; Mokranjac, D; Sichting, M; Neupert, W; Hell, K. The J domain-related cochaperone Tim16 is a constituent of the mitochondrial TIM23 preprotein translocase. Nat. Struct. Mol. Biol 2004, 11, 234–241. [Google Scholar]
- Li, Y; Dudek, J; Guiard, B; Pfanner, N; Rehling, P; Voos, W. The presequence translocaseassociated protein import motor of mitochondria. Pam16 functions in an antagonistic manner to Pam18. J. Biol. Chem 2004, 279, 38047–38054. [Google Scholar]
- Mokranjac, D; Bourenkov, G; Hell, K; Neupert, W; Groll, M. Structure and function of Tim14 and Tim16, the J and J-like components of the mitochondrial protein import motor. EMBO J 2006, 25, 4675–4685. [Google Scholar]
- Davey, KM; Parboosingh, JS; McLeod, DR; Chan, A; Casey, R; Ferreira, P; Snyder, FF; Bridge, PJ; Bernier, FP. Mutation of DNAJC19, a human homologue of yeast inner mitochondrial membrane co-chaperones, causes DCMA syndrome, a novel autosomal recessive Barth syndrome-like condition. J. Med. Genet 2006, 43, 385–393. [Google Scholar]
- Jubinsky, PT; Messer, A; Bender, J; Morris, RE; Ciraolo, GM; Witte, DP; Hawley, RG; Short, MK. Identification and characterization of Magmas, a novel mitochondria-associated protein involved in granulocyte-macrophage colony-stimulating factor signal transduction. Exp. Hematol 2001, 29, 1392–1402. [Google Scholar]
- Jubinsky, PT; Short, MK; Mutema, G; Morris, RE; Ciraolo, GM; Li, M. Magmas expression in neoplastic human prostate. J. Mol. Histol 2005, 36, 69–75. [Google Scholar]
- Jubinsky, PT; Short, MK; Mutema, G; Witte, DP. Developmental expression of Magmas in murine tissues and its co-expression with the GM-CSF receptor. J. Histochem. Cytochem 2003, 51, 585–596. [Google Scholar]
- Peng, J; Huang, CH; Short, MK; Jubinsky, PT. Magmas gene structure and evolution. Silico Biol 2005, 5, 251–263. [Google Scholar]
- Iosefson, O; Levy, R; Marom, M; Slutsky-Leiderman, O; Azem, A. The Pam18/Tim14-Pam16/Tim16 complex of the mitochondrial translocation motor: The formation of a stable complex from marginally stable proteins. Protein Sci 2007, 16, 316–322. [Google Scholar]
- Neupert, W; Herrmann, JM. Translocation of proteins into mitochondria. Annu. Rev. Biochem 2007, 76, 723–749. [Google Scholar]
- Kang, PJ; Ostermann, J; Shilling, J; Neupert, W; Craig, EA; Pfanner, N. Requirement for hsp70 in the mitochondrial matrix for translocation and folding of precursor proteins. Nature 1990, 348, 137–143. [Google Scholar]
- Westermann, B; Prip-Buus, C; Neupert, W; Schwarz, E. The role of the GrpE homologue, Mge1p, in mediating protein import and protein folding in mitochondria. EMBO J 1995, 14, 3452–3460. [Google Scholar]
- Laloraya, S; Gambill, BD; Craig, EA. A role for a eukaryotic GrpE-related protein, Mge1p, in protein translocation. Proc. Natl. Acad. Sci. USA 1994, 91, 6481–6485. [Google Scholar]
- Bolliger, L; Deloche, O; Glick, BS; Georgopoulos, C; Jeno, P; Kronidou, N; Horst, M; Morishima, N; Schatz, G. A mitochondrial homolog of bacterial GrpE interacts with mitochondrial hsp70 and is essential for viability. EMBO J 1994, 13, 1998–2006. [Google Scholar]
- Mokranjac, D; Sichting, M; Popov-Celeketic, D; Berg, A; Hell, K; Neupert, W. The import motor of the yeast mitochondrial TIM23 preprotein translocase contains two different J proteins, Tim14 and Mdj2. J. Biol. Chem 2005, 280, 31608–31614. [Google Scholar]
- Bonshtien, AL; Weiss, C; Vitlin, A; Niv, A; Lorimer, GH; Azem, A. Significance of the Nterminal domain for the function of chloroplast cpn20 chaperonin. J. Biol. Chem 2007, 282, 4463–4469. [Google Scholar]
- Weiss, C; Niv, A; Azem, A. Two-step purification of mitochondrial Hsp70, Ssc1p, using Mge1(His)(6) immobilized on Ni-agarose. Protein Expr. Purif 2002, 24, 268–273. [Google Scholar]


 ) CD spectra of hTim16/Pam16s (
) CD spectra of hTim16/Pam16s (
 ) CD spectra of yTim14/Pam18s-hTim16/Pam16s complex (
) CD spectra of yTim14/Pam18s-hTim16/Pam16s complex (
 ) CD spectra of hTim14/Pam18s/-hTim16/Pam16s complex. Similar spectra were obtained for all constructs presented in Table 1.
) CD spectra of hTim16/Pam16s (
) CD spectra of yTim14/Pam18s-hTim16/Pam16s complex (
) CD spectra of hTim14/Pam18s/-hTim16/Pam16s complex. Similar spectra were obtained for all constructs presented in Table 1.
) CD spectra of hTim14/Pam18s/-hTim16/Pam16s complex. Similar spectra were obtained for all constructs presented in Table 1.
) CD spectra of hTim16/Pam16s (
) CD spectra of yTim14/Pam18s-hTim16/Pam16s complex (
) CD spectra of hTim14/Pam18s/-hTim16/Pam16s complex. Similar spectra were obtained for all constructs presented in Table 1. ) 0.1 mg/mL hTim16/Pam16s. B) (
) 0.1 mg/mL yTim14/Pam16s-hTim16/Pam16s. C) (
) 0.1 mg/mL hTim14/Pam18-hTim16/Pam16s.
) 0.1 mg/mL hTim16/Pam16s. B) (
) 0.1 mg/mL yTim14/Pam16s-hTim16/Pam16s. C) (
) 0.1 mg/mL hTim14/Pam18-hTim16/Pam16s.
) 0.1 mg/mL hTim16/Pam16s. B) (
) 0.1 mg/mL yTim14/Pam16s-hTim16/Pam16s. C) (
) 0.1 mg/mL hTim14/Pam18-hTim16/Pam16s.
) 0.1 mg/mL hTim16/Pam16s. B) (
) 0.1 mg/mL yTim14/Pam16s-hTim16/Pam16s. C) (
) 0.1 mg/mL hTim14/Pam18-hTim16/Pam16s.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name of construct | Tm (°C) |
|---|---|
| yTim14/Pam18s | 16.5 |
| yTim16/Pam16s | 29 |
| yTim14/Pam18-yTim16/Pam16 | 41 |
| hTim14/Pam18s | 45 |
| hTim16/Pam16 | 22.5 |
| hTim14/Pam18s-yTim16/Pam16s | 52 |
| yTim14/Pam18s-hTim16/Pam16s | 35 |
| hTim14/Pam18s-hTim16/Pam18s | 49 |
| Description | Amino acids included in construct | Abbreviated name | |
|---|---|---|---|
| 1 | soluble domain of human Tim14/Pam18 | 24–116 | hTim14s/Pam18s |
| 2 | soluble domain of human Tim16/Pam16 | 24–125 | hTim16s/Pam16s |
| 3 | soluble domain of yeast Tim14/Pam18* | 84–168 | yTim14s |
| 4 | soluble domain of yeast Tim16/Pam16* | 25–130 | yTim16s/Pam16s |
| 5 | yeast Tim14s/Pam18s in complex with human Tim16s/Pam16s | 84–168 | yTim14s/Pam18s-hTim16s/Pam16s |
| 24–125 | |||
| 6 | Human Tim14s/Pam18s in complex with yeast Tim16s/Pam16s | 24–116 | hTim14s/Pam18s-yTim16s/Pam16s |
| 25–130 | |||
| 7 | human Tim14/Pam18 in complex with human Tim16s/Pam16s* | 24–116 | hTim14s/Pam18s-hTim16s/Pam16s |
| 24–125 | |||
| 8 | yeast Tim14s/Pam18s in complex with yeast Tim16s/Pam16 | 84–168 | yTim14s/Pam18s-yTim16s/Pam16s |
| 25–130 | |||
| 9 | full length yeast Tim14 | 1–168 | yTim14/Pam18 |
| 10 | yeast Tim14/Pam18 in complex with yeast Tim16/Pam16 (both full length) | 1–168 | yTim14s/Pam18s-yTim16s/Pam16s |
| 1–149 | |||
© 2009 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Elsner, S.; Simian, D.; Iosefson, O.; Marom, M.; Azem, A. The Mitochondrial Protein Translocation Motor: Structural Conservation between the Human and Yeast Tim14/Pam18-Tim16/Pam16 co-Chaperones. Int. J. Mol. Sci. 2009, 10, 2041-2053. https://doi.org/10.3390/ijms10052041
Elsner S, Simian D, Iosefson O, Marom M, Azem A. The Mitochondrial Protein Translocation Motor: Structural Conservation between the Human and Yeast Tim14/Pam18-Tim16/Pam16 co-Chaperones. International Journal of Molecular Sciences. 2009; 10(5):2041-2053. https://doi.org/10.3390/ijms10052041
Chicago/Turabian StyleElsner, Shira, Dana Simian, Ohad Iosefson, Milit Marom, and Abdussalam Azem. 2009. "The Mitochondrial Protein Translocation Motor: Structural Conservation between the Human and Yeast Tim14/Pam18-Tim16/Pam16 co-Chaperones" International Journal of Molecular Sciences 10, no. 5: 2041-2053. https://doi.org/10.3390/ijms10052041
APA StyleElsner, S., Simian, D., Iosefson, O., Marom, M., & Azem, A. (2009). The Mitochondrial Protein Translocation Motor: Structural Conservation between the Human and Yeast Tim14/Pam18-Tim16/Pam16 co-Chaperones. International Journal of Molecular Sciences, 10(5), 2041-2053. https://doi.org/10.3390/ijms10052041