Reactive Oxygen Species Enhance TLR10 Expression in the Human Monocytic Cell Line THP-1

Abstract
:1. Introduction
2. Results and Discussion
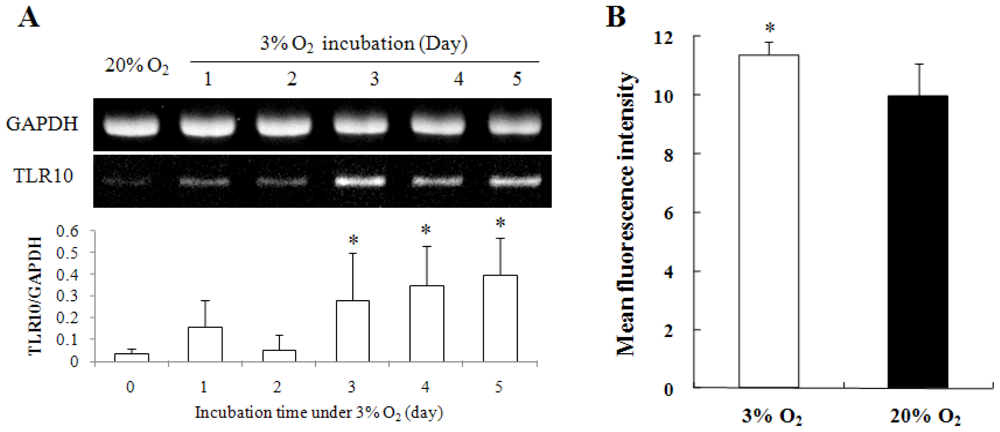
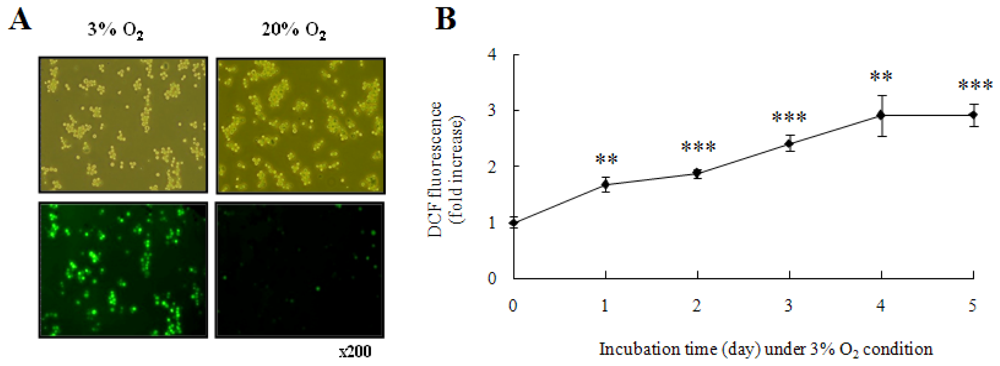
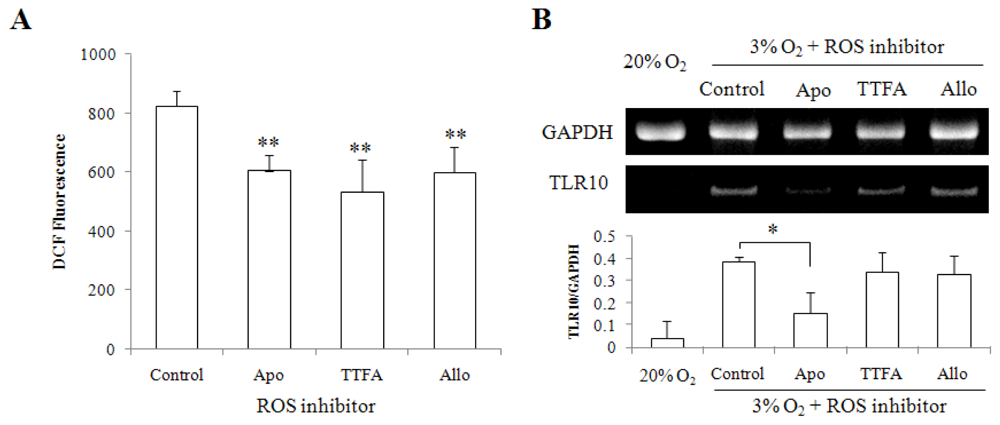
2.1. Hypoxia Enhanced Intracellular ROS Production and TLR10 Expression in THP-1 Cells
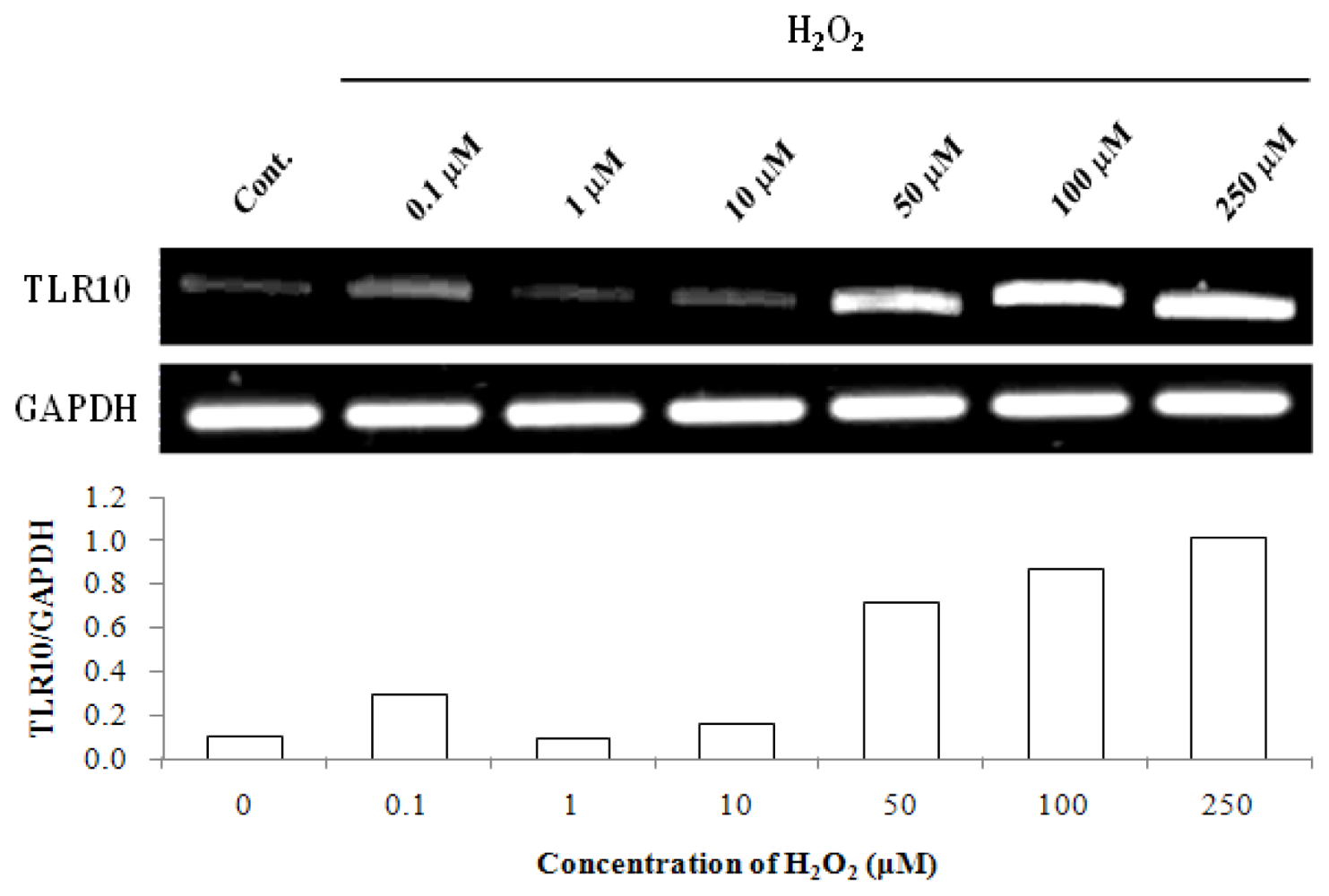
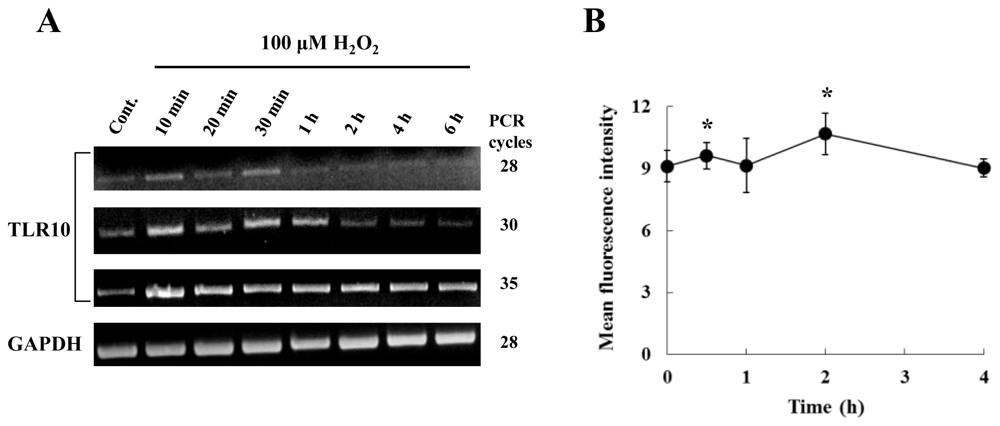
2.2. Hydrogen Peroxide Enhanced TLR10 Expression in THP-1 Cells in a Concentration-and Time-Dependent Manner
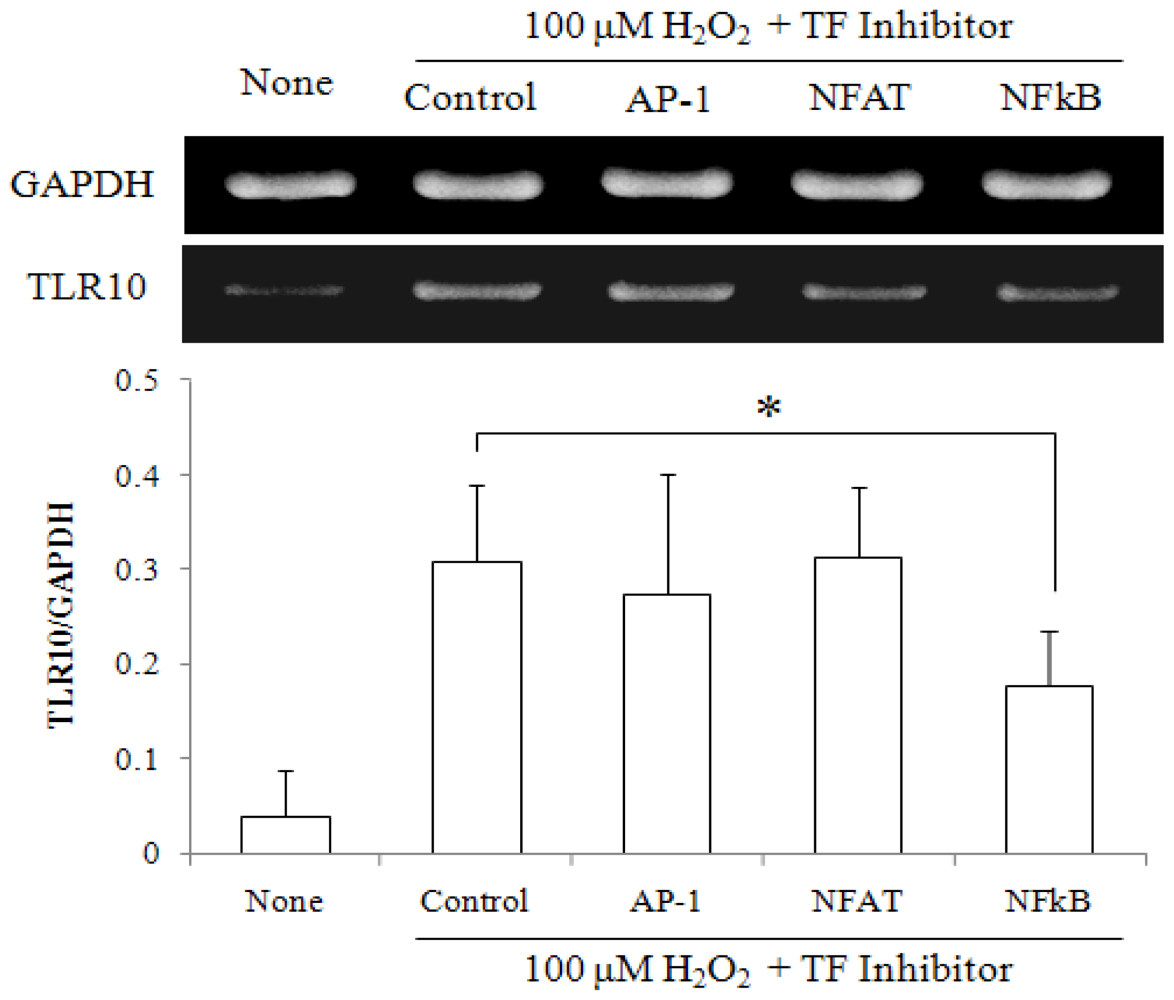
2.3. NF-κB Inhibitor Suppressed H2O2-Induced TLR10 Up-Regulation
2.4. Discussion
3. Experimental Section
3.1. Reagents
3.2. Cell Culture
3.3. RT-PCR
3.4. Measurement of Reactive Oxygen Species
3.5. Flow Cytometric Analysis
3.6. Prediction of TF Binding Sites
3.7. Luciferase Reporter Gene Assay
3.8. Statistical Analysis
4. Conclusions
Acknowledgements
References
- Takeda, K; Kaisho, T; Akira, S. Toll-like receptors. Annu. Rev. Immunol 2003, 21, 335–376. [Google Scholar]
- Akira, S; Takeda, K. Toll-like receptor signaling. Nat. Rev. Immunol 2004, 4, 499–511. [Google Scholar]
- Bell, JK; Mullen, GE; Leifer, CA; Mazzoni, A; Davis, DR; Segal, DM. Leucin-rich repeats and pathogen recognition in Toll-like receptors. Trends Immunol 2003, 24, 528–533. [Google Scholar]
- Chuang, TH; Ulevitch, RJ. Identification of hTLR: A novel human Toll-like receptor preferentially expressed in immune cells. Biochim. Biophys. Acta 2001, 1518, 157–161. [Google Scholar]
- Hasan, U; Chaffois, C; Gaillard, C; Saulnier, V; Merck, E; Tancredi, S; Guiet, C; Briere, F; Valch, J; Lebeque, S; Trinchieri, G; Bates, EEM. Human TLR10 is a functional receptor, expressed by B cells and plasmacytoid dendritic cells, which activates gene transcription through MyD88. J. Immunol 2005, 174, 2942–2950. [Google Scholar]
- Nyman, T; Stenmark, P; Flodin, S; Johansson, I; Hammarstrom, M; Nordlund, P. The crystal structure of the human toll-like receptor 10 cytoplasmic domain reveals a putative signaling dimmer. J. Biol. Chem 2008, 283, 11861–11865. [Google Scholar]
- Murdoch, C; Muthana, M; Lewis, CE. Hypoxia regulates macrophage Functions in Inflammation. J. Immunol 2005, 175, 6257–6263. [Google Scholar]
- Ock, J; Jeong, J; Choi, WS; Lee, WH; Kim, SH; Kim, IK; Suk, K. Regulation of Toll-like receptor 4 expression and its signaling by hypoxia in cultured microglia. J. Neurosci. Res 2007, 85, 1989–1995. [Google Scholar]
- Kim, SY; Choi, YJ; Joung, SM; Lee, BH; Jung, YS; Lee, JY. Hypoxic stress up-regulates the expression of Toll-like receptor 4 in macrophages via hypoxia-inducible factor. Immunology 2009, 129, 516–524. [Google Scholar]
- Kuhlicke, J; Frick, JS; Morote-Garcia, JC; Rosenberger, P; Eltzschig, H. Hypoxia inducible factor (HIF)-1 coordinates induction of Toll-like receptors TLR2 and TLR6 during hypoxia. PLoS One 2007, 2, e1364. [Google Scholar]
- Nishi, K; Oda, T; Takabuchi, S; Oda, S; Fukuda, K; Adachi, T; Semenza, GL; Shingu, K; Hirota, K. LPS induces hypoxia-inducible factor 1 activation in macrophage-differentiated cells in a reactive oxygen species-dependent manner. Antioxid. Redox Signal 2008, 10, 983–995. [Google Scholar]
- Chandel, NS; McClintock, DS; Feliciano, CE; Wood, TM; Melendez, JA; Rodriguez, AM; Schumacker, PT. Reactive oxygen species generated at mitochondrial complex III stabilize hypoxia-inducible factor-1α during hypoxia: A mechanism of O2 sensing. J. Biol. Chem 2000, 275, 25130–25138. [Google Scholar]
- Ishida, I; Kubo, H; Suzuki, S; Suzuki, T; Akashi, S; Inoue, K; Maeda, S; Kikuchi, H; Kondo, T. Hypoxia diminishes toll-like receptor 4 expression through reactive oxygen species generated by mitochondria in endothelial cells. J. Immunol 2002, 169, 2069–2075. [Google Scholar]
- Chandel, NS; Maltepe, E; Goldwasser, E; Mathieu, CE; Simon, MC; Schumacker, PT. Mitochondrial reactive oxygen species trigger hypoxia-induced transcription. Proc. Natl. Acad. Sci 1998, 95, 11715–11720. [Google Scholar]
- Baran, CP; Zeigler, MM; Tridandapani, S; Marsh, CB. The role of ROS and RNS in regulating life and death of blood monocytes. Curr. Pharm. Des 2004, 10, 855–866. [Google Scholar]
- Kress, M; Riedl, B; Reeh, PW. Effect of oxygen radicals on nociceptive afferents in the rat skin in vitro. Pain 1995, 62, 87–94. [Google Scholar]
- Li, C; Jackson, RM. Reactive species mechanisms of cellular hypoxia-reoxygenation injury. Am. J. Physiol. Cell Physiol 2002, 282, 227–241. [Google Scholar]
- Weiss, JS. Oxygen, ischemia and inflammation. Acta Physiol. Scand 1986, 548, 9–37. [Google Scholar]
- Oh, SO; Hong, JH; Kim, YR; Yoo, HS; Lee, SH; Lim, K; Hwang, BD; Exton, JH; Park, SK. Regulation of phospholipase D2 by H2O2 in PC12 cells. J. Neurochem 2000, 75, 2445–2454. [Google Scholar]
- Servitja, JM; Masgrau, R; Pardo, R; Sarri, E; Picatoste, F. Effects of oxidative stress on phospholipid signaling in rat cultured astrocytes and brain slices. J. Neurochem 2000, 75, 788–794. [Google Scholar]
- Rehli, M; Poltorak, A; Schwarzfischer, L; Krause, SW; Andreesen, R; Beutler, B. PU.1 and interferon consensus sequence-binding protein regulate the myeloid expression of the human Toll-like receptor 4 gene. J. Biol. Chem 2000, 275, 9773. [Google Scholar]
- Semenza, GL. Targeting HIF-1 for cancer therapy. Nat. Rev. Cancer 2003, 3, 721–732. [Google Scholar]
- Pugh, CW; Ratcliffe, PJ. Regulation of angiogenesis by hypoxic role of the HIF system. Nat. Med 2003, 9, 677–684. [Google Scholar]
- Strieter, RM. Mastering innate immunity. Nat. Med 2003, 9, 512–513. [Google Scholar]
- Lazarus, R; Raby, BA; Lange, C; Silverman, EK; Kwiatkowski, DJ; Vercelli, D; Klimecki, WJ; Martinez, FD; Weiss, ST. Toll-like receptor 10 (TLR10) genetic variation is associated with asthma in two independent samples. Am. J. Respir. Crit. Care Med 2004, 170, 594–600. [Google Scholar]
- Zhou, XX; Jia, WH; Shen, GP; Qin, HD; Yu, XJ; Chen, LZ; Feng, QS; Shugart, YY; Zheng, YX. Sequence variants in toll-like receptor 10 are associated with nasopharyngeal carcinoma risk. Cancer Epidemiol. Biomarkers Prev 2006, 15, 862–866. [Google Scholar]
- Roach, JC; Glusman, G; Rowen, L; Kaur, A; Purcell, MK; Smith, KD; Hood, LE; Aderem, A. The evolution of vertebrate Toll-like receptors. Proc. Natl. Acad. Sci. USA 2005, 102, 9577–9582. [Google Scholar]
- Zhou, H; Gu, J; Lamont, SJ; Gu, X. Evolutionary analysis for functional divergence of the toll-like receptor gene family and altered functional constraints. J. Mol. Evol 2007, 65, 119–123. [Google Scholar]
- Sun, J; Wiklund, F; Zheng, SL; Chang, B; Bälter, K; Li, L; Johansson, JE; Li, G; Adami, HO; Liu, W; Tolin, A; Turner, AR; Meyers, DA; Isaacs, WB; Xu, J; Grönberg, H. Sequence variants in Toll-like receptor gene cluster (TLR6-TLR-1-TLR10) and prostate cancer risk. J. Natl. Cancer Inst 2005, 97, 525–532. [Google Scholar]
- Purdue, MP; Lan, Q; Wang, SS; Kricker, A; Menashe, I; Zhang, TZ; Hartge, P; Grulich, AE; Zhang, Y; Morton, LM; Vajdic, CM; Holford, TR; Severson, RK; Leaderer, BP; Cerhan, JR; Yeager, M; Cozen, W; Jacobs, K; Davis, S; Rothman, N; Chanock, SJ; Chatterjee, N; Armstrong, BK. A pooled investigation of Toll-like receptor gene variants and risk of non-Hodgkin lymphoma. Carcinogenesis 2009, 30, 275–281. [Google Scholar]
- Chandel, N; Schumacker, P; Arch, RH. Reactive oxygen species are downstream products of TRAF-mediated signal transduction. J. Biol. Chem 2001, 276, 42728–42736. [Google Scholar]
- Liu, H; Colaritti, R; Rovira, I; Finkel, T. Redox-dependent transcriptional regulation. Circ. Res 2005, 97, 967–974. [Google Scholar]
- Halliwell, B; Gutteridge, JMC. Free Radicals in Biology and Medicine; Oxford University Press: New York, NY, USA, 1999. [Google Scholar]
- Cummins, EP; Taylor, CT. Hypoxia-responsive transcription factors. Eur. J. Physiol. (Pflügers Archiv.) 2005, 450, 363–71. [Google Scholar]
- Ma, Q. Transcriptional responses to oxidative stress: pathological and toxicological implications. Pharmacol. Ther 2010, 125, 376–393. [Google Scholar]
- Nanduri, J; Yang, G; Kumar, GK; Semenza, GL; Prabhakar, NR. Transcriptional responses to intermittent hypoxia. Respir. Physiol. Neurobiol 2008, 277–281. [Google Scholar]
- Peyssonnaux, C; Datta, V; Cramer, T; Doedens, A; Theodorakis, EA; Gallo, RL; Hurtado-Ziola, N; Nizet, V; Johnson, RS. HIF-1a expression regulates the bactericidal capacity of phagocytes. J. Clin. Invest 2005, 115, 1806–1815. [Google Scholar]
- Anand, RJ; Gribar, SC; Li, J; Kohler, JW; Branca, MF; Dubowski, T; Sodhi, CP; Hackam, DJ. Hypoxia causes an increase in phagocytosis by macrophages in a HIF-1α-dependent manner. J. Leukoc. Biol 2007, 82, 1257–1265. [Google Scholar]
- Farré, D; Roset, R; Huerta, M; Adsuara, JE; Roselló, L; Albà, MM; Messeguer, X. Identification of patterns in biological sequences at the ALGGEN server: PROMO and MALGEN. Nucleic Acids Res 2002, 31, 3651–3653. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Transcription Factors | Relative Luciferase Activity (Fold) | |
|---|---|---|
| ROS Treated | Positive Control | |
| NF- κB | 1.46 ± 0.45* | 5.12 ± 1.52*** |
| AP-1 | 1.32 ± 0.26** | 18.37 ± 3.79*** |
| NF-AT | 1.21 ± 0.13*** | 3.04 ± 0.90*** |
© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Kim, D.; Kim, Y.J.; Koh, H.S.; Jang, T.Y.; Park, H.E.; Kim, J.Y. Reactive Oxygen Species Enhance TLR10 Expression in the Human Monocytic Cell Line THP-1. Int. J. Mol. Sci. 2010, 11, 3769-3782. https://doi.org/10.3390/ijms11103769
Kim D, Kim YJ, Koh HS, Jang TY, Park HE, Kim JY. Reactive Oxygen Species Enhance TLR10 Expression in the Human Monocytic Cell Line THP-1. International Journal of Molecular Sciences. 2010; 11(10):3769-3782. https://doi.org/10.3390/ijms11103769
Chicago/Turabian StyleKim, Donghee, Yeon Ju Kim, Hyun Sook Koh, Tae Yang Jang, Hyo Eun Park, and Jae Young Kim. 2010. "Reactive Oxygen Species Enhance TLR10 Expression in the Human Monocytic Cell Line THP-1" International Journal of Molecular Sciences 11, no. 10: 3769-3782. https://doi.org/10.3390/ijms11103769
APA StyleKim, D., Kim, Y. J., Koh, H. S., Jang, T. Y., Park, H. E., & Kim, J. Y. (2010). Reactive Oxygen Species Enhance TLR10 Expression in the Human Monocytic Cell Line THP-1. International Journal of Molecular Sciences, 11(10), 3769-3782. https://doi.org/10.3390/ijms11103769