The Effect of Exercise on the Skeletal Muscle Phospholipidome of Rats Fed a High-Fat Diet

Abstract
:1. Introduction
2. Results
2.1. Compliance with Training Programs and Physical Characteristics
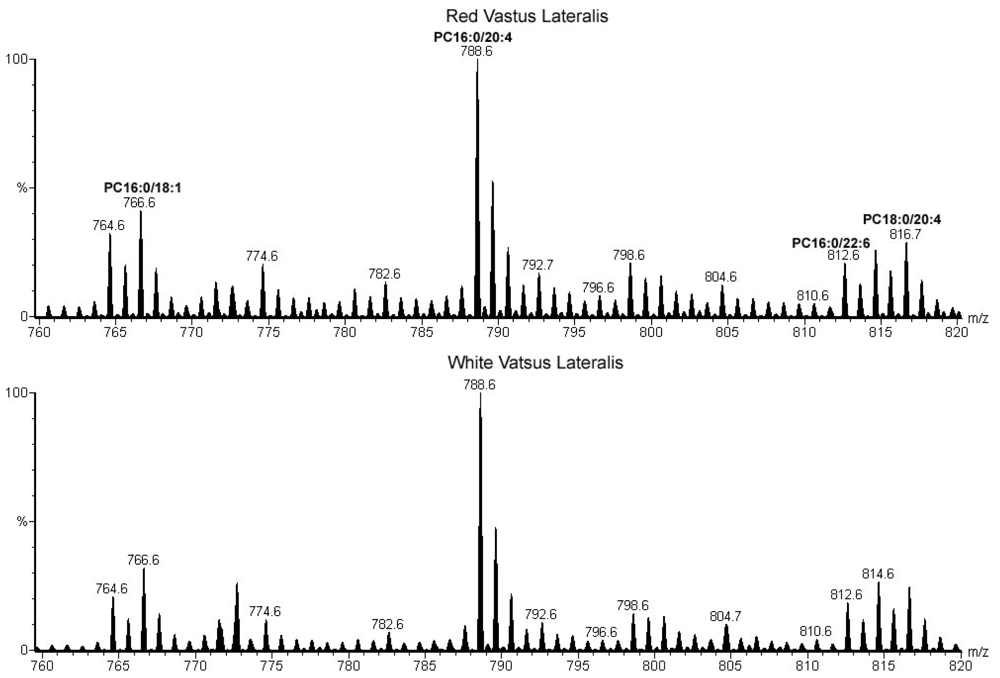
2.2. Comparison of Oxidative and Glycolytic Fibers
2.3. Comparison of Trained and Sedentary Rats
3. Discussion
4. Experimental Section
4.1. Animal Care, Dietary Treatments and Exercise-Training Program
4.2. Animal Sacrifice and Tissue Preparation
4.3. Phospholipid Extraction
4.4. Mass Spectrometry
4.5. Statistical Analysis
5. Conclusions
Acknowledgements
References
- Helge, JW; Kriketos, AD; Storlien, LH. Insulin sensitivity, muscle fibre types, and membrane lipids. Adv. Exp. Med 1998, 441, 129–138. [Google Scholar]
- Storlien, LH; Hulbert, AJ; Else, PL. Polyunsaturated fatty acids, membrane function and metabolic diseases such as diabetes and obesity. Curr. Opin. Clin. Nutr. Metab. Care 1998, 1, 559–563. [Google Scholar]
- Ekroos, K; Chernushevich, IV; Simons, K; Shevchenko, A. Quantitative profiling of phospholipids by multiple precursor ion scanning on a hybrid quadrupole time-of-flight mass spectrometer. Anal. Chem 2002, 74, 941–949. [Google Scholar]
- Han, X; Gross, RW. Shotgun lipidomics: electrospray ionization mass spectrometric analysis and quantitation of cellular lipidomes directly from crude extracts of biological samples. Mass Spectrom. Rev 2005, 24, 367–412. [Google Scholar]
- Pulfer, M; Murphy, RC. Electrospray mass spectrometry of phospholipids. Mass Spectrom. Rev 2003, 22, 332–364. [Google Scholar]
- Mitchell, TW; Turner, N; Hulbert, AJ; Else, PL; Hawley, JA; Lee, JS; Bruce, CR; Blanksby, SJ. Exercise alters the profile of phospholipid molecular species in rat skeletal muscle. J. Appl. Physiol 2004, 97, 1823–1829. [Google Scholar]
- Turner, N; Lee, JS; Bruce, CR; Mitchell, TW; Else, PL; Hulbert, AJ; Hawley, JA. Greater effect of diet than exercise training on the fatty acid profile of rat skeletal muscle. J. Appl. Physiol 2004, 96, 974–980. [Google Scholar]
- Lapachet, RA; Miller, WC; Arnall, DA. Body fat and exercise endurance in trained rats adapted to a high-fat and/or high-carbohydrate diet. J. Appl. Physiol 1996, 80, 1173–1179. [Google Scholar]
- Miller, WC; Bryce, GR; Conlee, RK. Adaptations to a high-fat diet that increase exercise endurance in male rats. J. Appl. Physiol 1984, 56, 78–83. [Google Scholar]
- Simi, B; Sempore, B; Mayet, MH; Favier, RJ. Additive effects of training and high-fat diet on energy metabolism during exercise. J. Appl. Physiol 1991, 71, 197–203. [Google Scholar]
- Burke, LM; Hawley, JA. Effects of short-term fat adaptation on metabolism and performance of prolonged exercise. Med. Sci. Sport Exer 2002, 34, 1492–1498. [Google Scholar]
- Lee, JS; Bruce, CR; Spriet, LL; Hawley, JA. Interaction of diet and training on endurance performance in rats. Exp. Physiol 2001, 86, 499–508. [Google Scholar]
- Han, X; Gross, RW. Electrospray ionization mass spectroscopic analysis of human erythrocyte plasma membrane phospholipids. Proc. Natl. Acad. Sci. USA 1994, 91, 10635–10639. [Google Scholar]
- Andersson, A; Nalsen, C; Tengblad, S; Vessby, B. Fatty acid composition of skeletal muscle reflects dietary fatty acid composition in humans. Am. J. Clin. Nutr 2002, 76, 1222–1229. [Google Scholar]
- Ayre, KJ; Hulbert, AJ. Dietary fatty acid profile influences the composition of skeletal muscle phospholipids in rats. J. Nutr 1996, 126, 653–662. [Google Scholar]
- Pan, DA; Storlien, LH. Dietary lipid profile is a determinant of tissue phospholipid fatty acid composition and rate of weight gain in rats. J. Nutr 1993, 123, 512–519. [Google Scholar]
- Schmid, PC; Deli, E; Schmid, HHO. Generation and remodeling of phospholipid molecular species in rat hepatocytes. Arch. Biochem. Biophys 1995, 319, 168–176. [Google Scholar]
- Ayre, KJ; Hulbert, AJ. Dietary fatty acid profile affects endurance in rats. Lipids 1997, 32, 1265–1270. [Google Scholar]
- Ruf, T; Valencak, T; Tataruch, F; Arnold, W. Running speed in mammals increases with muscle n-6 polyunsaturated fatty acid content. PloS ONE 2006, 1, e65. [Google Scholar]
- Cameron-Smith, D; Burke, LM; Angus, DJ; Tunstall, RJ; Cox, GR; Bonen, A; Hawley, JA; Hargreaves, M. A short-term, high-fat diet up-regulates lipid metabolism and gene expression in human skeletal muscle. Am. J. Clin. Nutr 2003, 77, 313–318. [Google Scholar]
- Conlee, RK; Hammer, RL; Winder, WW; Bracken, ML; Nelson, AG; Barnett, DW. Glycogen repletion and exercise endurance in rats adapted to a high fat diet. Metab. Clin. Exp 1990, 39, 289–294. [Google Scholar]
- Divine-Patch, L; Brooks, GA. Effects of training on VO2 max and VO2 during two running intensities in rats. Pflügers Arch 1980, 386, 215–219. [Google Scholar]
- Delp, MD; Duan, C. Composition and size of type I, IIA, IID/X, and IIB fibers and citrate synthase activity of rat muscle. J. Appl. Physiol 1996, 80, 261–270. [Google Scholar]
- Folch, J; Lees, M; Sloane-Stanley, GH. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem 1957, 226, 497–509. [Google Scholar]
- Han, X; Yang, J; Cheng, H; Ye, H; Gross, RW. Toward fingerprinting cellular lipidomes directly from biological samples by two-dimensional electrospray ionization mass spectrometry. Anal. Biochem 2004, 330, 317–331. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Molecular species | m/z [M + Li]+ | SED | TRAINED | P Value | |||
|---|---|---|---|---|---|---|---|
| WVL | RVL | WVL | RVL | T | M | ||
| PC16:0/18:1 | 766.6 | 12.5 ± 0.3 | 15.4 ± 0.5† | 12.3 ± 0.6 | 14.4 ± 0.3† | NS | <0.0001 |
| PC16:0/18:2 | 764.6 | 10.4 ± 0.4 | 12.6 ± 0.3 | 12.8 ± 0.9 | 16.1 ± 1.0*† | 0.0005 | 0.0007 |
| PC16:0/20:4 | 788.6 | 43.4 ± 0.7 | 36.0 ± 1.0† | 42.6 ± 0.9 | 36.0 ± 0.4† | NS | <0.0001 |
| PC16:0/22:6 | 812.6 | 8.3 ± 0.4 | 8.8 ± 0.5 | 7.2 ± 0.3 | 6.8 ± 0.3* | 0.0006 | NS |
| PC18:0/18:2, PC18:1/18:1 | 792.6 | 6.0 ± 0.5 | 8.0 ± 0.3 | 7.6 ± 1.8 | 8.5 ± 0.3 | NS | NS |
| PC18:0/20:4 | 816.6 | 9.4 ± 0.4 | 10.0 ± 0.3 | 8.4 ± 0.8 | 10.1 ± 0.2 | NS | 0.02 |
| PC18:1/20:4 | 814.6 | 10.1 ± 0.3 | 9.1 ± 0.1 | 9.2 ± 0.7 | 8.2 ± 0.3 | 0.04 | 0.02 |
| Molecular species | m/z [M − H] − | SED | TRAINED | P Value | |||
|---|---|---|---|---|---|---|---|
| WVL | RVL | WVL | RVL | T | M | ||
| PA16:0/18:2 | 671.5 | 1.1 ± 0.0 | 1.2 ± 0.0 | 1.2 ± 0.0 | 1.5 ± 0.1 | NS | 0.05 |
| PA16:0/20:4, PA18:2/18:2 | 695.5 | 2.6 ± 0.1 | 2.8±0.1 | 2.5 ± 0.1 | 2.6 ± 0.1 | NS | NS |
| PA18:0/20:4, CL(18:2)4 | 723.5 | 0.5 ± 0.0 | 0.9 ± 0.1 | 0.3 ± 0.0 | 0.9 ± 0.0 | NS | 0.005 |
| PA18:0,18:2, PA18:1/18:1 | 699.5 | 2.0 ± 0.0 | 2.5 ± 0.1 | 1.9 ± 0.1 | 2.5 ± 0.1† | NS | 0.0007 |
| PA18:0/22:6, PA18:1/22:5, PG16:0/18:1 | 747.6 | 6.5 ± 0.1 | 6.2 ± 0.1 | 6.9 ± 0.1 | 6.2 ± 0.1 | NS | NS |
| PA18:1/18:2 | 697.5 | 1.4 ± 0.1 | 2.1 ± 0.1 | 1.9 ± 0.1 | 2.8 ± 0.1† | 0.02 | 0.002 |
| PE16:0p/18:2 | 698.5 | 1.3 ± 0.0 | 1.9 ± 0.1 | 1.8 ± 0.1 | 2.6 ± 0.1† | 0.006 | 0.003 |
| PE16:0p/20:4 | 722.5 | 9.2 ± 0.2 | 9.1 ± 0.2 | 9.9 ± 0.4 | 9.6 ± 0.4 | NS | NS |
| PE16:0/22:6, PS16:0/18:0 | 762.6 | 2.6 ± 0.1 | 2.4 ± 0.1 | 2.4 ± 0.0 | 2.3 ± 0.1 | NS | NS |
| PE18:0/18:2, PE18:1/18:1 | 742.6 | 3.9 ± 0.1 | 5.4 ± 0.1 | 4.9 ± 0.1 | 7.6 ± 0.3*† | 0.0005 | <0.0001 |
| PE18:0/20:4 | 766.6 | 23.1 ± 0.4 | 24.8 ± 0.5 | 24.2 ± 0.2 | 27.0 ± 0.2 | NS | 0.02 |
| PE18:0/22:6 | 790.6 | 10.1 ± 0.2 | 13.4 ± 0.4† | 8.9 ± 0.2 | 12.0 ± 0.2† | 0.02 | <0.0001 |
| PS18:0/22:6 | 834.6 | 3.6 ± 0.2 | 2.3 ± 0.2 | 2.8 ± 0.2 | 1.9 ± 0.2 | NS | 0.05 |
| PI18:0/20:4 | 885.6 | 29.2 ± 0.4 | 21.7 ± 0.5† | 27.9 ± 0.6 | 18.3 ± 0.9† | NS | <0.0001 |
| PI18:0/22:5 | 911.6 | 1.4 ± 0.0 | 1.5 ± 0.1 | 1.3 ± 0.0 | 1.1 ± 0.0 | 0.04 | NS |
| PI18:0/22:6 | 909.6 | 1.5 ± 0.0 | 1.7 ± 0.1 | 1.3 ± 0.0 | 1.3 ± 0.0 | 0.02 | NS |
| Fatty acid | %Total |
|---|---|
| 16:0 | 17.9 |
| 16:1 (n−7) | 20.9 |
| 18:0 | 19.7 |
| 18:1(n−9) | 35.1 |
| 18:2 (n−6) | 6.4 |
© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Mitchell, T.W.; Turner, N.; Else, P.L.; Hulbert, A.J.; Hawley, J.A.; Lee, J.S.; Bruce, C.R.; Blanksby, S.J. The Effect of Exercise on the Skeletal Muscle Phospholipidome of Rats Fed a High-Fat Diet. Int. J. Mol. Sci. 2010, 11, 3954-3964. https://doi.org/10.3390/ijms11103954
Mitchell TW, Turner N, Else PL, Hulbert AJ, Hawley JA, Lee JS, Bruce CR, Blanksby SJ. The Effect of Exercise on the Skeletal Muscle Phospholipidome of Rats Fed a High-Fat Diet. International Journal of Molecular Sciences. 2010; 11(10):3954-3964. https://doi.org/10.3390/ijms11103954
Chicago/Turabian StyleMitchell, Todd W., Nigel Turner, Paul L. Else, Anthony J. Hulbert, John A. Hawley, Jong Sam Lee, Clinton R. Bruce, and Stephen J. Blanksby. 2010. "The Effect of Exercise on the Skeletal Muscle Phospholipidome of Rats Fed a High-Fat Diet" International Journal of Molecular Sciences 11, no. 10: 3954-3964. https://doi.org/10.3390/ijms11103954
APA StyleMitchell, T. W., Turner, N., Else, P. L., Hulbert, A. J., Hawley, J. A., Lee, J. S., Bruce, C. R., & Blanksby, S. J. (2010). The Effect of Exercise on the Skeletal Muscle Phospholipidome of Rats Fed a High-Fat Diet. International Journal of Molecular Sciences, 11(10), 3954-3964. https://doi.org/10.3390/ijms11103954