Estrogen Stimulates Proliferation and Differentiation of Neural Stem/Progenitor Cells through Different Signal Transduction Pathways


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Primary Cultures
2.3. Western Immunoblot Analysis
2.4. BrdU Labeling
2.5. MTT Assay
2.6. Immunocytochemistry
3. Results and Discussion
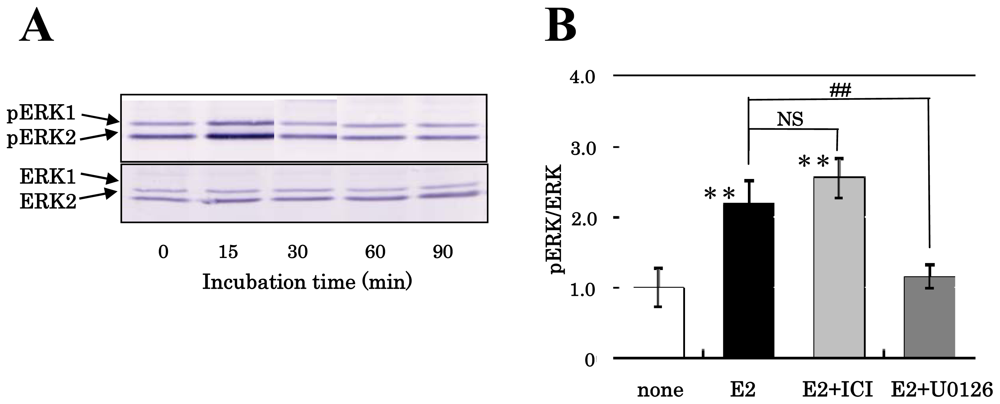
3.1. Effect of Estrogens on the Phosphorylation of MAPK/ERK of NS/PCs
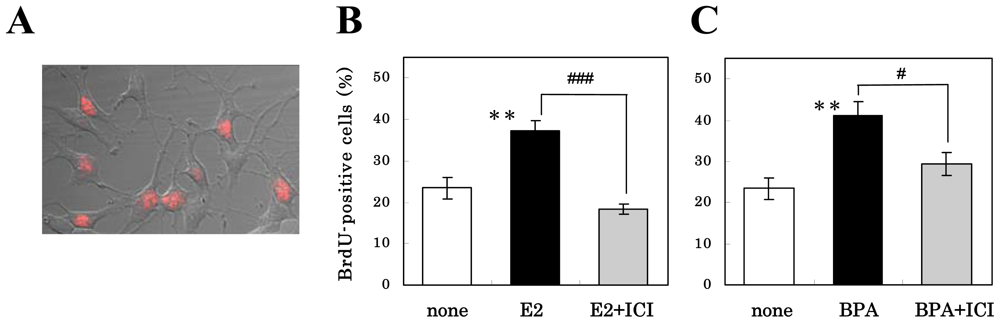
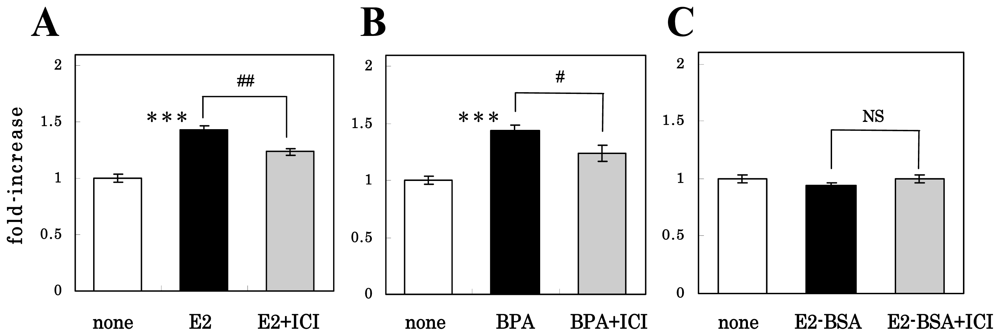
3.2. Mechanism of Action of Estrogens on Proliferation of NS/PCs
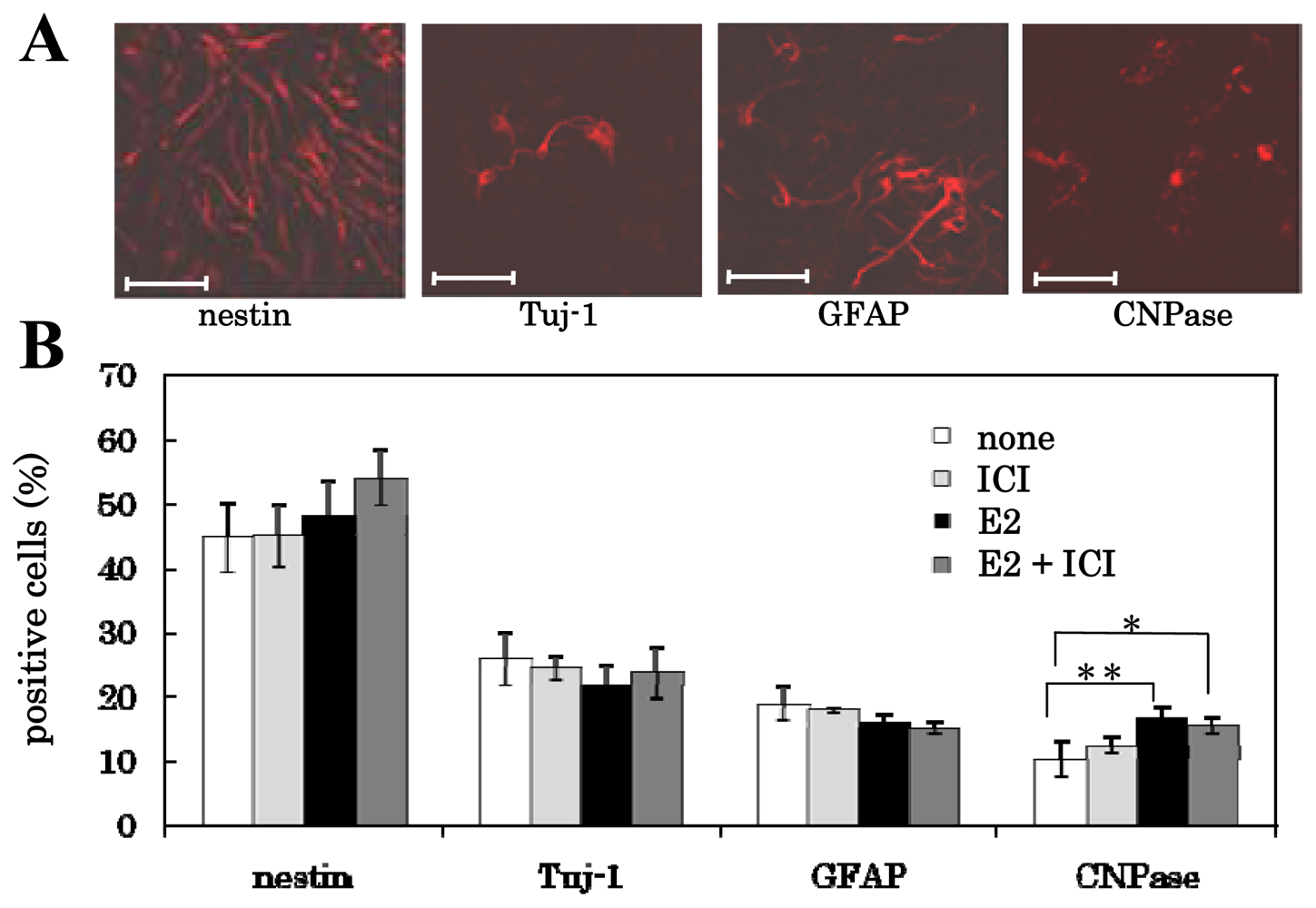
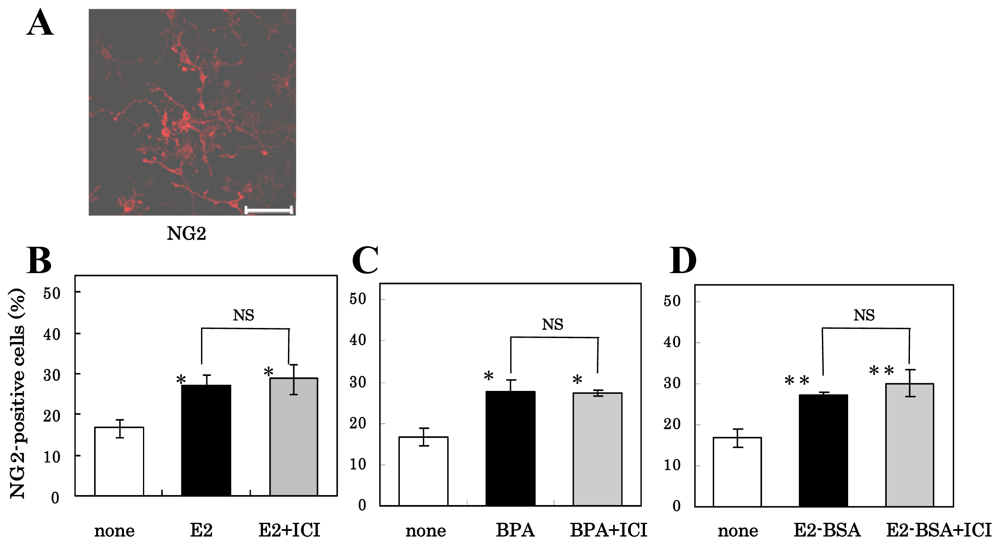
3.3. Mechanism of Action of Estrogens on Differentiation of NS/PCs
4. Conclusions
References
- Ebert, AD; Beres, AJ; Barber, AE; Svendsen, CN. Human neural progenitor cells over-expressing IGF-1 protect dopamine neurons and restore function in a rat model of Parkinson's disease. Exp. Neurol 2008, 209, 213–223. [Google Scholar]
- Imitola, J; Khoury, SJ. Neural stem cells and the future treatment of neurological diseases: Raising the standard. Methods Mol. Biol 2008, 438, 9–16. [Google Scholar]
- Lu, P; Jones, LL; Snyder, EY; Tuszynski, MH. Neural stem cells constitutively secrete neurotrophic factors and promote extensive host axonal growth after spinal cord injury. Exp. Neurol 2003, 181, 115–129. [Google Scholar]
- Lovell-Badge, R. The future for stem cell research. Nature 2001, 414, 88–91. [Google Scholar]
- Song, RX; Santen, RJ. Apoptotic action of estrogen. Apoptosis 2003, 8, 55–60. [Google Scholar]
- Watt, FM; Hogan, BLM. Out of eden: Stem cells and their niches. Science 2000, 287, 1427–1430. [Google Scholar]
- Brännvall, K; Korhonen, L; Lindholm, D. Estrogen-receptor-dependent regulation of neural stem cell proliferation and differentiation. Mol. Cell. Neurosci 2002, 21, 512–520. [Google Scholar]
- Brännvall, K; Bogdanovic, N; Korhonen, L; Lindholm, D. 19-Nortestosterone influences neural stem cell proliferation and neurogenesis in the rat brain. Eur. J. Neurosci 2005, 21, 871–878. [Google Scholar]
- McEven, BS. Estrogen actions throughout the brain. Recent Prog. Horm. Res 2002, 57, 357–384. [Google Scholar]
- Okada, M; Murase, K; Makino, A; Nakajima, M; Kaku, T; Furukawa, S; Furukawa, Y. Effects of estrogens on proliferation and differentiation of neural stem/progenitor cells. Biomed. Res 2008, 29, 163–170. [Google Scholar]
- Beato, M; Herrlich, P; Schutz, G. Steroid hormone receptors: Many actors in search of a plot. Cell 1995, 83, 851–857. [Google Scholar]
- Falkenstein, E; Tillmann, HC; Christ, M; Feuring, M; Wehling, M. Multiple actions of steroid hormones: A focus on rapid, nongenomic effects. Pharmacol. Rev 2000, 52, 513–556. [Google Scholar]
- Toran-Allerand, CD. Minireview: A plethora of estrogen receptors in the brain: Where will it end? Endocrinology 2004, 145, 1069–1074. [Google Scholar]
- Qiu, J; Bosch, MA; Tobias, SC; Krust, A; Graham, SM; Murphy, SJ; Korach, KS; Chambon, P; Scanlan, TS; R⊘nnekleiv, OK; Kelly, MJ. A G-protein-coupled estrogen receptor is involved in hypothalamic control of energy homeostasis. J. Neurosci 2006, 26, 5649–5655. [Google Scholar]
- Thomas, P; Pang, Y; Filardo, EJ; Dong, J. Identity of an estrogen membrane receptors coupled to a G protein in human breast cancer cells. Endocrinology 2005, 146, 624–632. [Google Scholar]
- Björnström, L; Sjöberg, M. Mechanism of estrogen receptor signaling convergence of genomic and nongenomic actions on target genes. Mol. Endocrinol 2005, 19, 833–842. [Google Scholar]
- Bouskine, A; Nebout, M; Mograbi, B; Brücker-Davis, F; Roger, C; Fenichel, P. Estrogens promote human testicular germ cell cancer through a membrane-mediated activation of extracellular regulated kinase and protein kinase A. Endocrinology 2008, 149, 565–573. [Google Scholar]
- Titolo, D; Mayer, CM; Dhillon, SS; Cai, F; Belsham, DD. Estrogen facilitates both phosphatidylinositol 3-kinase/Akt and ERK1/2 mitogen-activated protein kinase membrane signaling required for long-term neuropeptide Y transcriptional regulation in clonal, immortalized neurons. J. Neuroci 2008, 28, 6473–6482. [Google Scholar]
- Mendelsohn, EM; Karas, RH. Rapid progress for non-nuclear estrogen receptor signaling. J. Clin. Inv 2010, 120, 2277–2279. [Google Scholar]
- Wakeling, AE; Dukes, M; Bowler, J. A potent specific pure antiestrogen with clinical potential. Cancer Res 1991, 51, 3867–3873. [Google Scholar]
- Howell, A; DeFriend, DJ; Robertson, JF; Blamey, RW; Anderson, L; Anderson, E; Sutcliffe, FA; Walton, P. Pharmacokinetics, pharmacological and anti-tumour effects of the specific anti-oestrogen ICI 182,780 in women with advanced breast cancer. Br. J. Cancer 1996, 74, 300–308. [Google Scholar]
- Furukawa, Y; Urano, T; Minamimura, M; Nakajima, M; Okuyama, S; Furukawa, S. 4-Methylcatechol-induced heme oxygenase-1 exerts a protective effect against oxidative stress in cultured neural stem/progenitor cells via PI3 kinase/Akt pathway. Biomed. Res 2010, 31, 45–52. [Google Scholar]
- Dominguez, R; Jalali, C; de Lacalle, S. Morphological effects of estrogen on cholinergic neurons in vitro involves activation of extracellular signal-regulated kinases. J. Neurosci 2004, 24, 982–990. [Google Scholar]
- Zhao, L; O’Neill, K; Brinton, RD. Estrogenic agonist activity of ICI 182,780 (faslodex) in hippocampal neurons: Implications for basic science understanding of estrogen signaling and development of estrogen modulators with a dual therapeutic profile. J. Pharm. Exp. Ther 2006, 319, 1124–1132. [Google Scholar]
- Samuels, IS; Karlo, JC; Faruzzi, AN; Pickering, K; Herrup, K; Sweatt, JD; Saitta, SC; Landreth, GE. Deletion of ERK2 mitogen-activated protein kinase identifies its key roles in cortical neurogenesis and cognitive function. J. Neurosci 2008, 28, 6983–6995. [Google Scholar]
- Kudo, C; Wada, K; Masuda, T; Yonemura, T; Shibuya, A; Fujimoto, Y; Nakajima, A; Niwa, H; Kamisaki, Y. Nonylphenol induces the death of neural stem cells due to activation of the caspase cascade and regulation of the cell cycle. J. Neurochem 2003, 88, 1416–1423. [Google Scholar]
- Song, RX; Santen, RJ. Apoptotic action of estrogen. Apoptosis 2003, 8, 55–60. [Google Scholar]
- Lebesque, D; Traub, M; Butte-Smith, MD; Chen, C; Zukin, RS; Kelly, MJ; Etgen, AM. Acute administration of non-classical estrogen receptor agonists attenuates ischemia-induced hippocampal neuron loss in middle-aged female rats. PLoS One 2010, 30, 12950–12957. [Google Scholar]





© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Okada, M.; Makino, A.; Nakajima, M.; Okuyama, S.; Furukawa, S.; Furukawa, Y. Estrogen Stimulates Proliferation and Differentiation of Neural Stem/Progenitor Cells through Different Signal Transduction Pathways. Int. J. Mol. Sci. 2010, 11, 4114-4123. https://doi.org/10.3390/ijms11104114
Okada M, Makino A, Nakajima M, Okuyama S, Furukawa S, Furukawa Y. Estrogen Stimulates Proliferation and Differentiation of Neural Stem/Progenitor Cells through Different Signal Transduction Pathways. International Journal of Molecular Sciences. 2010; 11(10):4114-4123. https://doi.org/10.3390/ijms11104114
Chicago/Turabian StyleOkada, Makiko, Akihisa Makino, Mitsunari Nakajima, Satoshi Okuyama, Shoei Furukawa, and Yoshiko Furukawa. 2010. "Estrogen Stimulates Proliferation and Differentiation of Neural Stem/Progenitor Cells through Different Signal Transduction Pathways" International Journal of Molecular Sciences 11, no. 10: 4114-4123. https://doi.org/10.3390/ijms11104114
APA StyleOkada, M., Makino, A., Nakajima, M., Okuyama, S., Furukawa, S., & Furukawa, Y. (2010). Estrogen Stimulates Proliferation and Differentiation of Neural Stem/Progenitor Cells through Different Signal Transduction Pathways. International Journal of Molecular Sciences, 11(10), 4114-4123. https://doi.org/10.3390/ijms11104114