Whole Cell Imprinting in Sol-Gel Thin Films for Bacterial Recognition in Liquids: Macromolecular Fingerprinting

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
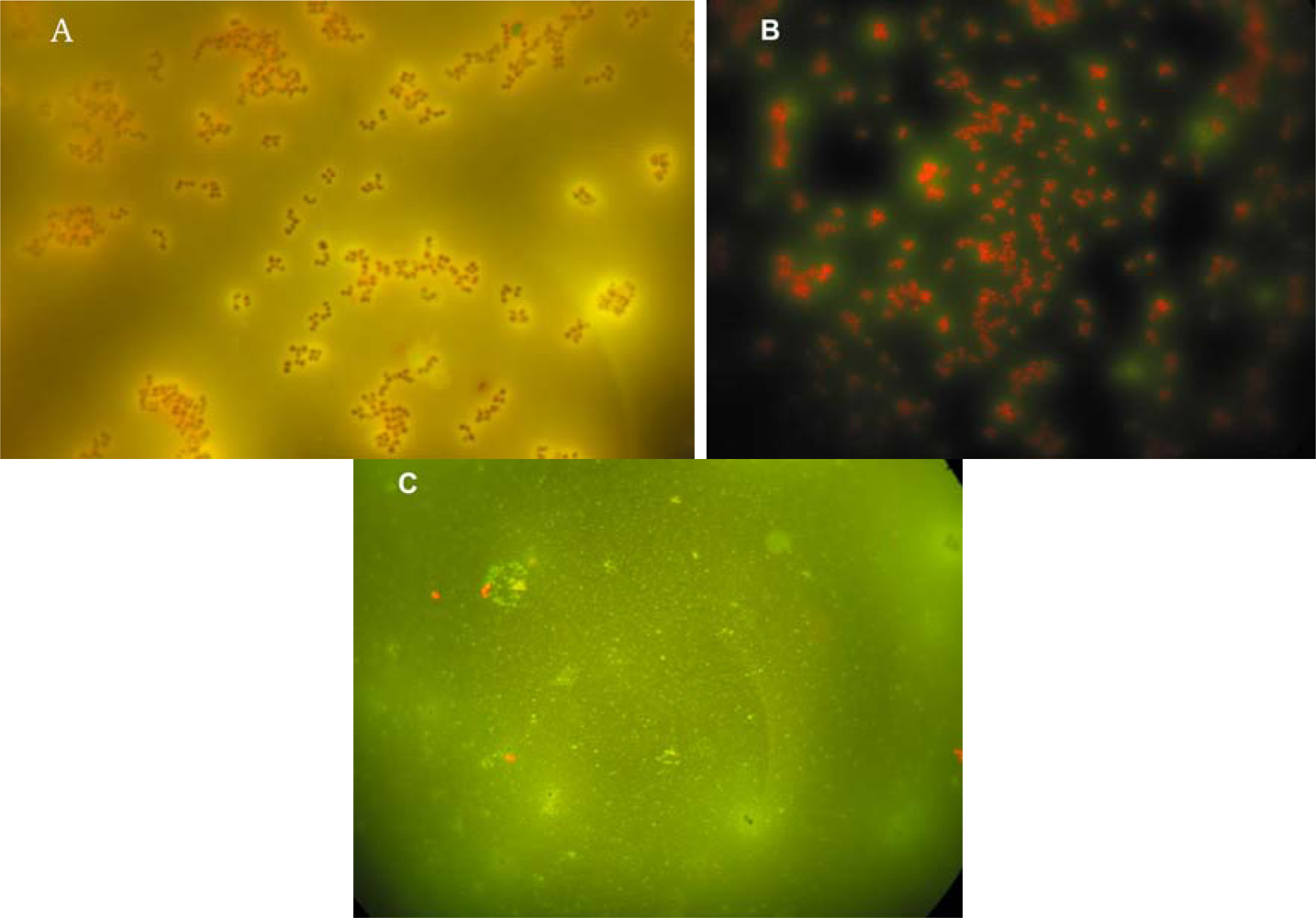
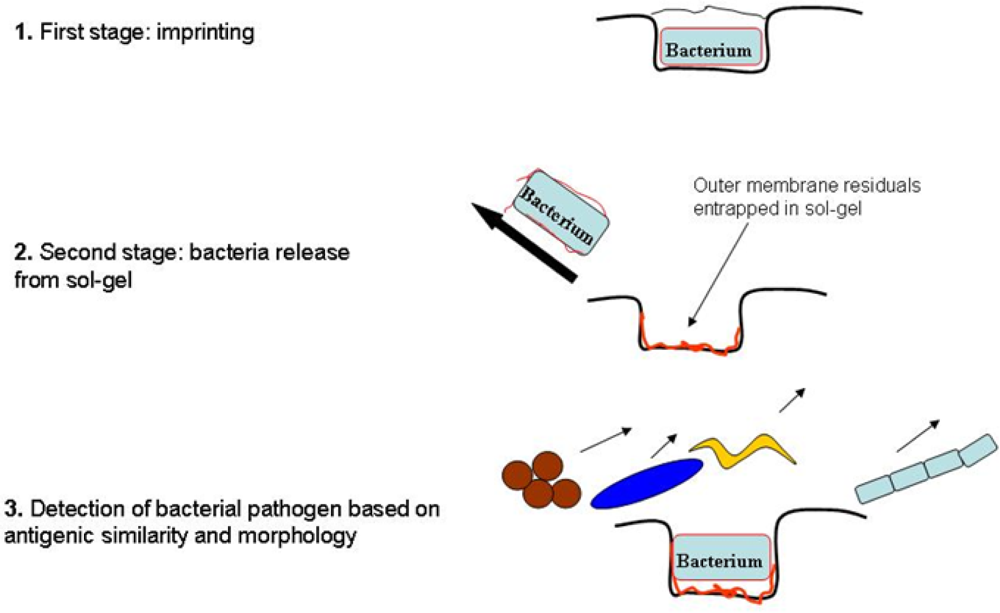
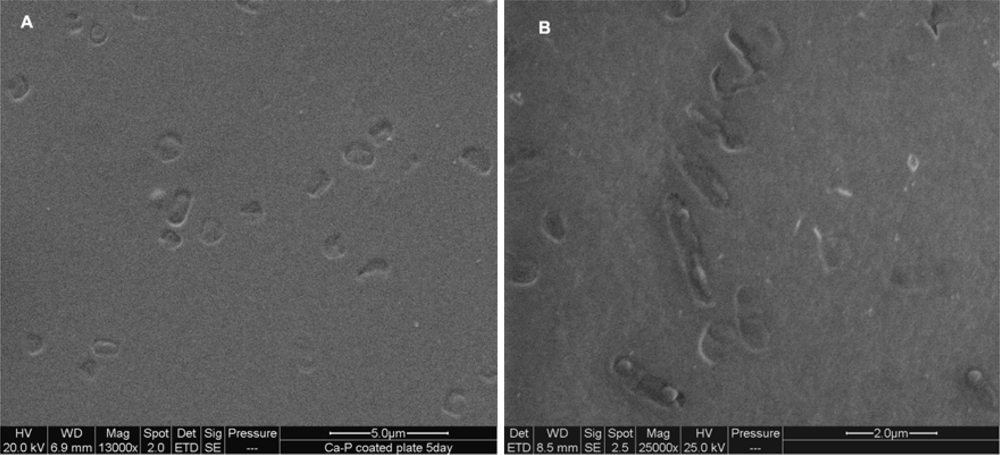
2.1. Imprinting of SG Films with Whole Cell Bacteria
2.2. Analysis of SG Entrapped Membrane Residuals
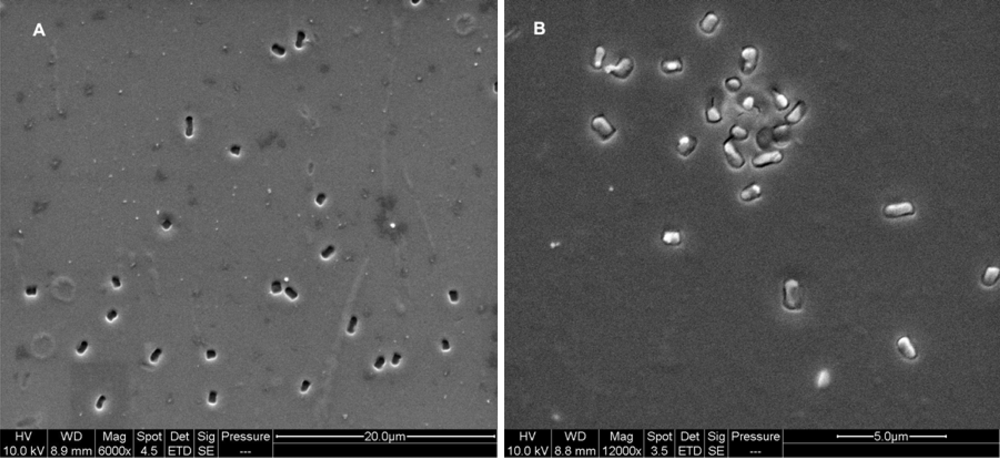
2.3. Imprinting of SG-films with Other Microorganisms
2.4. Sensitivity and Selectivity of Imprinted SG-films
3. Experimental Section
3.1. Experimental Microorganisms and Growth Conditions
3.2. Imprinted Sol-Gel Films Preparation
3.2.1. Stock of Sol-Gel Solution
3.2.2. Template cells preparation
3.3. Imprinting Procedure
3.4. Detection of Sensitivity and Selectivity Ability of Imprinted SG-films
3.5. Analysis of Outer Cell Components Left in Sol-Gel Cavities Following Templates Removal
3.5.1. Bacteria Lysates
3.5.2. Membrane Components Extraction from Sol-Gel
3.5.3. Membrane Proteins/Glycoproteins SDS-PAGE Determination
4. Conclusions
Acknowledgments
References and Notes
- Hench, LL; West, JK. Chemical Processing of Advanced Materials; A Wiley-Interscience Publication, John Wiley & Sons, Inc.: New-York, NY, USA, 1992. [Google Scholar]
- Soft Chemistry Routes to New Materials; Rouxel, J; Tourmoux, M; Brec, R (Eds.) Materials Science Forum, Translator; Tech. Publications Inc.: Zürich, Switzerland, 1994; Volume 152–153.
- Sanchez, C; Ribot, F. Design of hybrid organic-inorganic materials synthesized via sol-gel chemistry. New J. Chem 1994, 18, 1007–10047. [Google Scholar]
- Avnir, D; Klein, LC; Levy, D; Shubert, U; Wojcik, AB. Organo-silica sol-gel materials. In The Chemistry of Organic Silicon Compounds (e-book); Rapaport, Z, Apeloig, Y, Eds.; Wiley: New York, NY, USA, 1998; Volume 2, pp. 2317–2362. [Google Scholar]
- Walcarius, A; Collinson, MM. Analytical Chemistry with silica sol-gels: Traditional routes to new materials for chemical analysis. Annu. Rev. Anal. Chem 2009, 2, 121–143. [Google Scholar]
- Parejo, PG; Zayat, M; Levy, D. Highly efficient UV-absorbing thin-film coatings for protection of organic materials against photodegradation. J. Mater. Chem 2006, 16, 2165–2169. [Google Scholar]
- Komarmeni, S; Sakka, S; Phule, PP; Laine, RM. Sol-Gel Synthesis and Processing; Ceramic Transactions, ACS: Westerville, OH, USA, 1998; Volume 95. [Google Scholar]
- Dunn, B; Zink, JI. Sol-gel chemistry and materials. Acc. Chem. Res 2007, 40, 729. [Google Scholar]
- Jeronimo, PCA; Araujo, AN; Montenegro, MCBSM. Optical sensors and biosensors based on sol-gel films: Review. Talanta 2007, 72, 13–27. [Google Scholar]
- Lukowiak, A; Strek, W. Sensing abilities of materials prepared by sol-gel technology. J. Sol-Gel Sci. Technol 2009, 50, 201–215. [Google Scholar]
- Avnir, D; Coradin, T; Lev, O; Livage, J. Resent bio-applications of sol-gel materials. J. Mater. Chem 2006, 16, 1013–1030. [Google Scholar]
- Besanger, TR; Brennan, JD. Entrapment of membrane proteins in sol-gel derived silica. J. Sol-Gel Sci. Technol 2006, 40, 209–225. [Google Scholar]
- Livage, J; Coradin, T; Roux, C. Encapsulation of biomolecules in silica gels. J. Phys. Condens. Mater 2001, 13, R673–691. [Google Scholar]
- Premkumar, JR; Rosen, R; Belkin, Sh; Lev, O. Sol-gel luminescence biosensors: Encapsulation of recombinant E.coli reporters in thick silicate films. Anal. Chim. Acta 2002, 462, 11–23. [Google Scholar]
- Coradin, T; Livage, J. Aqueous silicates in biological sol-gel applications new perspectives for old precursors. Acc. Chem. Res 2007, 40, 819–826. [Google Scholar]
- Takeuchi, T; Mukawa, T; Asanuma, H. Molecular Imprinting From fundamentals to Applications; Wiley-VCH: Weinheim, Germany, 2003. [Google Scholar]
- Lieberzeit, PA; Schirk, C; Glanznig, G; Gazda-Miarecka, S; Bindeus, R; Nannen, H; Kauling, J; Dickert, FL. From nano-pattering to functionality–surface and bulk imprinting for analytical purposes. Superlatt. Microstruct 2004, 36, 133–142. [Google Scholar]
- Fireman-Shoresh, Sh; Turyan, I; Mandler, D; Avnir, D; Marx, Sh. Chiral electrochemical recognition by very thin molecularly imprinted sol-gel films. Langmuir 2005, 21, 7842–7847. [Google Scholar]
- Premkumar, JR; Sagi, E; Rosen, R; Belkin, S; Modestov, ED; Lev, O. Fluorescent bacteria encapsulated in sol-gel derived silicate films. Chem. Mater 2002, 14, 2676–2686. [Google Scholar]
- Nguyen-Ngoc, H; Tran-Minh, C. Fluorescent biosensor using whole cells in an inorganic translucent matrix. Annu. Chim. Acta 2007, 583, 161–165. [Google Scholar] [Green Version]
- Trögl, J; Ripp, S; Kancova, G; Sayler, GS; Churava, A; Pařík, P; Demnerová, K; J Hálová, J; Kubicová, L. Selectivity of whole cells optical biosensor with immobilized bioreporter Pseodomonas fluorescence HK44. Sens. Actuat. B 2005, B107, 98–110. [Google Scholar]
- Hurst, CJ; Knudsen, GR; McInerny, MJ; Stetzenbach, LD; Walter, MV. Manual of Environmental Microbiology; ASM Press: Washington, DC, USA, 1997. [Google Scholar]
- Dickert, FL; Lieberzeit, PA; Heyden, O. Sensor strategies for microorganism detection- from physical principles to imprinting procedures. Anal. Bioanal. Chem 2003, 377, 540–549. [Google Scholar]
- Dickert, FL; Heyden, O. Bioimprinting of polymers and sol-gel phases. Selective detection of yeasts with imprinted polymers. Anal. Chem 2002, 74, 1302–1306. [Google Scholar]
- Cunliffe, D; Alexander, C. Molecular Imprinted Polymers: Science and Technology; Yan, M, Ramström, O, Eds.; Dekker: New-York, NY, USA, 2002; pp. 249–283. [Google Scholar]
- Perez, N; Alexander, C; Vulfson, EN. Surface imprinting of microorganisms. In Molecularly Imprinted Polymers Man-Made Mimics of Antibodies and Their Applications in Analytical Chemistry; Sellergren, B, Ed.; Elsevier Science: Amsterdam, The Netherlands, 2001; Chapter 11, pp. 295–303. [Google Scholar]
- Harvey, SD; Mong, GM; Ozanich, RM; Mclean, JS; Goodvin, Sh.M; Valentine, NB; Fredrikson, JK. Preparation and evaluation of spore-specific affinity-augmented bio-imprinted beads. Anal. Bioanal. Chem 2006, 386, 211–2127. [Google Scholar]
- Armon, R; Dosoretz, C; Starosvetsky, J; Orshansky, F; Saadi, I. Sol-gel applications in environmental biotechnology. J. Biotechnol 1996, 51, 279–285. [Google Scholar]
- Armon, R; Starosvetsky, J; Saadi, I. Sol-gel as reaction matrix for bacterial enzymatic activity. J. Sol-Gel Sci. Technol 2000, 19, 289–292. [Google Scholar]
- Dosoretz, C; Armon, R; Starosvetzky, J; Rothschild, N. Entrapment of parathion hydrolase from Pseudomonas spp. in sol-gel glass. J. Sol-Gel Sci. Technol 1996, 7, 7–11. [Google Scholar]
- Armon, R; Starosvetzky, J; Green, M. Denitrification by a mixture of bacterial strains derived from an up-flow sludge blanket reactor, following entrapment in sol-gel glass. Lett. Appl. Microbiol 1995, 20, 25–28. [Google Scholar]
- Laor, Y; Zolkov, C; Armon, R. Immobilizing humic acid in a sol-gel matrix: a new tool to study humic-contaminants sorption interactions. Environ. Sci. Technol 2002, 36, 1054–1060. [Google Scholar]
- Zolkov, C; Avnir, D; Armon, R. Tissue-derived cell growth on hybrid sol-gel films. J. Mater. Chem 2004, 14, 2200–2205. [Google Scholar]
- Zolkov, C; Avnir, D; Armon, R. Sol-gel based poliovirus-1 detector. J. Virol. Methods 2009, 155, 132–135. [Google Scholar]
- Murray, RGE; Hall, M; Thompson, BG. Cell division in Deinococcus radiodurans. Can. J. Microbiol 1983, 29, 1412–1423. [Google Scholar]
- Armon, R; Arella, M; Payment, P. A highly efficient second-step concentration technique for bacteriophages and enteric viruses using ammonium sulfate and Tween 80. Can. J. Microbiol 1988, 34, 651–655. [Google Scholar]
- Wanner, G; Formanek, H; Galli, D; Wirth, R. Localization of aggregation substances of Enterococcus faecalis after induction by sex pheromones. An ultrastructural comparison using immuno labelling, transmission and high resolution scanning electron microscopic techniques. Arch. Microbiol 1989, 151, 491–497. [Google Scholar]
- Schaer-Zammaretti, P; Ubbink, J. Imaging of lactic acid bacteria with AFM- elasticity and adhesion maps and their relationship to biological and structural data. Ultramicroscopy 2003, 97, 199–208. [Google Scholar]
- Bradford, M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem 1976, 72, 248–254. [Google Scholar]
- Andrews, AT. Electrophoresis: Theory, Techniques, and Biochemical and Clinical Applications, 2nd ed; Clarendon Press: Oxford, UK, 1986. [Google Scholar]
- Oakley, BR; Kirsch, DR; Morris, NR. A simplified ultrasensitive silver stain for detecting proteins in polyacrylamide gels. Anal Biochem 1980, 105, 361–363. [Google Scholar]
- Baumeister, W; Barth, M; Hegerl, R; Guckenberger, R; Hahn, M; Saxton, WO. Three-dimensional structure of the regular surface layer (HPI layer) of Deinococcus radiodurans. J. Mol. Biol 1986, 187, 241–250. [Google Scholar]
- Thompson, BG; Murray, RG. Isolation and characterization of the plasma membrane and the outer membrane of Deinococcus radiodurans strain Sark. Can. J. Microbiol 1981, 27, 729–734. [Google Scholar]
- Thompson, BG; Murray, RGE; Boyce, JF. The association of the surface array and the outer membrane of Deinococcus radiodurans. Can. J. Microbiol 1982, 28, 1081–1088. [Google Scholar]
- Hayden, O; Dickert, FL. Selective microorganisms detection with cell surface imprinted polymers. Adv. Mater 2001, 13, 1480–1483. [Google Scholar]
- Hayden, O; Bindeus, R; Haderspock, C; Mann, K-J; Wirl, B; Dickert, FL. Mass-sensitive detection of cells, viruses and enzymes with artificial receptors. Sens. Actuat. B 2003, B91, 316–319. [Google Scholar]
- Diaz-Garcia, ME; Laiño, RB. Molecular imprinting in sol-gel materials: recent developments and applications. Microchim. Acta 2005, 149, 19–36. [Google Scholar]










© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Cohen, T.; Starosvetsky, J.; Cheruti, U.; Armon, R. Whole Cell Imprinting in Sol-Gel Thin Films for Bacterial Recognition in Liquids: Macromolecular Fingerprinting. Int. J. Mol. Sci. 2010, 11, 1236-1252. https://doi.org/10.3390/ijms11041236
Cohen T, Starosvetsky J, Cheruti U, Armon R. Whole Cell Imprinting in Sol-Gel Thin Films for Bacterial Recognition in Liquids: Macromolecular Fingerprinting. International Journal of Molecular Sciences. 2010; 11(4):1236-1252. https://doi.org/10.3390/ijms11041236
Chicago/Turabian StyleCohen, Tally, Jeanna Starosvetsky, Uta Cheruti, and Robert Armon. 2010. "Whole Cell Imprinting in Sol-Gel Thin Films for Bacterial Recognition in Liquids: Macromolecular Fingerprinting" International Journal of Molecular Sciences 11, no. 4: 1236-1252. https://doi.org/10.3390/ijms11041236
APA StyleCohen, T., Starosvetsky, J., Cheruti, U., & Armon, R. (2010). Whole Cell Imprinting in Sol-Gel Thin Films for Bacterial Recognition in Liquids: Macromolecular Fingerprinting. International Journal of Molecular Sciences, 11(4), 1236-1252. https://doi.org/10.3390/ijms11041236