Erwinia mallotivora sp., a New Pathogen of Papaya (Carica papaya) in Peninsular Malaysia

Abstract
:1. Introduction
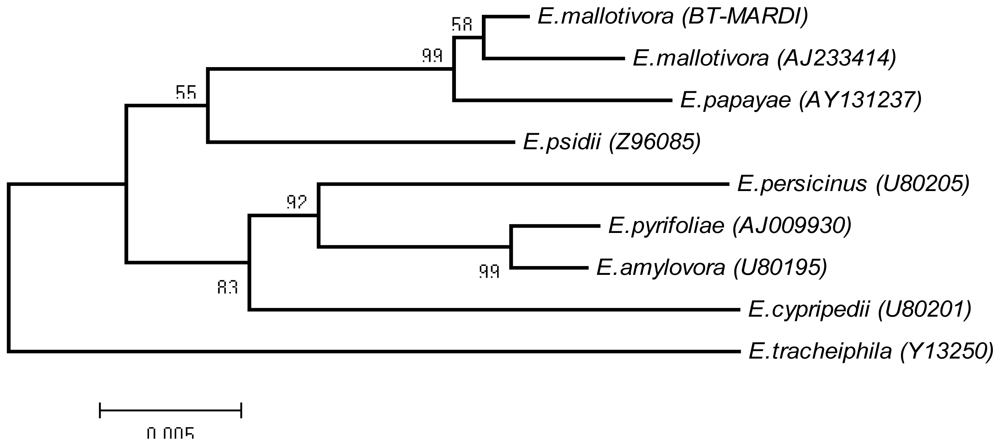
2. Results and Discussion
3. Experimental Section
3.1. Bacterial Isolation
3.2. Koch’s Postulate
3.3. PCR Amplification and Cloning of 16S rRNA Gene
4. Conclusions
Acknowledgements
References
- Gwyne, DC. Fire blight in perry pears and cider apples in the South West of England. Acta Hortic 1984, 151, 41–47. [Google Scholar]
- Reifschneider, FJB; Lopes, CA. Bacterial top and stalk rot of maize in Brazil. Plant Dis 1982, 66, 519–520. [Google Scholar]
- Rabu, MR; Mat Lin, R. Prospect of papaya in the world market: Malaysia perspective. Proceeding of First International Symposium on Papaya, Genting Higlands, Malaysia, 22–24 November 2005.
- Maktar, NH; Kamis, S; Mohd Yusof, FZ; Hussain, NH. Erwinia papayae causing papaya dieback in Malaysia. Plant Pathol 2008, 57, 774. [Google Scholar]
- Gardan, L; Christen, R; Achouak, W; Prior, P. Erwinia papayae sp. nov., a pathogen of papaya (Carica papaya). IJSEM 2004, 54, 107–113. [Google Scholar]
- King, EO; Ward, MK; Raney, DE. Two simple media for the demonstration of pyocyanin and fluorescin. J. Lab. Clin. Med 1954, 44, 301–307. [Google Scholar]
- Goto, M. Erwinia mallotivora sp. nov., the causal organism of bacterial leaf spot of Mallotus japonicus. Int. J. Syst. Bacteriol 1976, 26, 467–473. [Google Scholar]
- Perombelon, MCM. Potato diseases caused by soft rot erwinias: an overview of pathogenesis. Plant Pathol 2002, 51, 1–12. [Google Scholar]
- Toth, IK; Bell, KS; Holeva, MC; Birch, PRJ. Soft rot erwiniae: from genes to genomes. Mol. Plant Pathol 2003, 4, 17–30. [Google Scholar]
- Leu, LS; Lee, CC; Huang, TC. Papaya black rot caused by Erwinia cypripedii. Plant Prot. Bull 1980, 22, 377–384. [Google Scholar]
- Trujillo, EE; Schroth, MN. Two bacterial diseases of papaya trees caused by Erwinia species in the Northern Mariana Island. Plant Dis 1982, 66, 116–120. [Google Scholar]
- Webb, RR. Epidemiology and control of bacterial canker of papaya caused by an Erwinia sp. on St. Croix, U.S. Virgin Island. Plant Dis 1985, 69, 305–309. [Google Scholar]
- Hildedrand, M; Dickler, E; Geider, K. Occurance of Erwinia amylovora on insects in a fire blight orchard. J. Phytopathol 2000, 148, 251–256. [Google Scholar]
- Agrios, GN. Plant Pathology, 5th ed; Elsevier; Academic Press: London, UK, 2005; pp. 398–401. [Google Scholar]
- Schaad, NW. Laboratory Guide for Identification of Plant Pathogenic Bacteria, 2nd ed; APS Press: St. Paul, MN, USA, 1988; p. 41. [Google Scholar]
- Gardan, L; Gouy, C; Christen, R; Samson, R. Elevation of three subspecies of Pectobacterium carotovorum to species level: Pectobacterium atrosepticum sp. nov., Pectobacterium betavasculorum sp. nov. and Pectobacterium wasabiae sp. nov. IJSEM 2002, 53, 381–391. [Google Scholar]
- Lee, A; Taylor, GH. A comparison of electron staining agents for the transmission electron microscopy examination of coal. Int. J. Coal Geol 1999, 39, 329–337. [Google Scholar]
- Weisburg, WG; Barns, SM; Pelletier, DA; Lane, DJ. 16S ribosomal DNA amplification for phylogenetic study. J. Bacteriol 1991, 173, 697–703. [Google Scholar]
- Kumar, S; Dudley, J; Nei, M; Tamura, K. MEGA: A biologist-centric software for evolutionary analysis of DNA and protein sequences. Brief. Bioinf 2008, 9, 299–306. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Characteristic | E. mallotivora (BT-Mardi) | E. mallotivora [7] | E. papaya [5] |
|---|---|---|---|
| Blue pigment on King’s B agar | − | − | + |
| Citrate utilization | + | + | −a |
| Reducing substances from sucrose | + | + | − |
| d-Mannitol | + | + | − |
| l-Arabinose | − | − | + |
© 2011 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Mat Amin, N.; Bunawan, H.; Redzuan, R.A.; Jaganath, I.B.S. Erwinia mallotivora sp., a New Pathogen of Papaya (Carica papaya) in Peninsular Malaysia. Int. J. Mol. Sci. 2011, 12, 39-45. https://doi.org/10.3390/ijms12010039
Mat Amin N, Bunawan H, Redzuan RA, Jaganath IBS. Erwinia mallotivora sp., a New Pathogen of Papaya (Carica papaya) in Peninsular Malaysia. International Journal of Molecular Sciences. 2011; 12(1):39-45. https://doi.org/10.3390/ijms12010039
Chicago/Turabian StyleMat Amin, Noriha, Hamidun Bunawan, Rohaiza Ahmad Redzuan, and Indu Bala S. Jaganath. 2011. "Erwinia mallotivora sp., a New Pathogen of Papaya (Carica papaya) in Peninsular Malaysia" International Journal of Molecular Sciences 12, no. 1: 39-45. https://doi.org/10.3390/ijms12010039
APA StyleMat Amin, N., Bunawan, H., Redzuan, R. A., & Jaganath, I. B. S. (2011). Erwinia mallotivora sp., a New Pathogen of Papaya (Carica papaya) in Peninsular Malaysia. International Journal of Molecular Sciences, 12(1), 39-45. https://doi.org/10.3390/ijms12010039